ЭЛЕКТРОФОРЕЗ ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ | Поликлиника ЦКБ РЖД-Медицина
Электрофорез лекарственных веществ – особый электрофармакологический метод, основанный на сочетанном использовании постоянного тока и вводимых с его помощью лекарственных веществ. Из электрических токов для лекарственного электрофореза применяются гальванический (в 80-85 %), диадинамические, синусоидальные модулированные (в выпрямленном режиме), прямоугольный импульсный и флюктуирующий (форма № 3) токи.
Механизм физиологического воздействия
При электрофорезе лекарственные вещества в организм проникают через выводные протоки потовых и сальных желез, межклеточные промежутки, волосяные фолликулы и в меньшей степени – чресклеточно. Во время процедуры лекарственные вещества проникают неглубоко: сразу после элекрофореза основная часть лекарства обнаруживается в эпидермисе и дерме, создавая депо. Однако от процедуры к процедуре глубина электрогенного перемещения вводимого препарата возрастает. К тому же следует иметь в виду, что за счет диффузии часть лекарственных веществ быстро достигает кровеносных и лимфатических сосудов, разносясь ко всем органам и тканям. Весьма важно, что из кровотока лекарственные вещества вторично поступают преимущественно в органы и ткани, расположенные в зоне проведения процедуры. Это обосновывает целесообразность использования лекарственного электрофореза для лечения как поверхностно, так и глубоко расположенных патологических процессов, а также заболеваний внутренних органов.
Однако от процедуры к процедуре глубина электрогенного перемещения вводимого препарата возрастает. К тому же следует иметь в виду, что за счет диффузии часть лекарственных веществ быстро достигает кровеносных и лимфатических сосудов, разносясь ко всем органам и тканям. Весьма важно, что из кровотока лекарственные вещества вторично поступают преимущественно в органы и ткани, расположенные в зоне проведения процедуры. Это обосновывает целесообразность использования лекарственного электрофореза для лечения как поверхностно, так и глубоко расположенных патологических процессов, а также заболеваний внутренних органов.
Действие лекарственного электрофореза как электрофармакологического метода складывается из сочетанного действия физического фактора (гальванический или другие токи) и введенного лекарственного вещества. Ответная реакция организма при этом не является простой суммацией эффектов, вызванных этими двумя факторами, составляющими единый терапевтический комплекс. Она значительно сложнее и разнообразнее. Важно помнить, что действие вводимых электрофорезом лекарств развивается несколькими путями (рефлекторное, местное и гуморальное) и, варьируя технику и методику проведения процедуры, ими можно управлять.
Она значительно сложнее и разнообразнее. Важно помнить, что действие вводимых электрофорезом лекарств развивается несколькими путями (рефлекторное, местное и гуморальное) и, варьируя технику и методику проведения процедуры, ими можно управлять.
Особенности и достоинства лекарственного электрофореза:
1. Лекарственные вещества, вводимые электрофорезом, задерживаются в поверхностных слоях кожи и образуют здесь так называемое кожное депо ионов. В нем лекарства могут сохраняться от 12-24 ч до 15-20 суток (адреналин, цинк, медь и др.). Задержка введенных веществ в кожном депо способствует их более длительному действию и медленному выведению из организма.
2. Метод лекарственного электрофореза позволяет создавать высокую локальную (в патологическом очаге) концентрацию препарата, не насыщая им весь организм. Согласно имеющимся данным, после электрофореза содержание лекарств в тканях области воздействия в несколько раз выше, чем после общепринятых способов введения той же дозы препарата.
3. В отличие от инъекционных способов введения электрофорез позволяет доставить лекарства к патологическому очагу, в котором имеются нарушения микроциркуляции и регионарного кровообращения в виде капиллярного стаза, тромбоза сосудов, инфильтрации и некроза. Такие патологические очаги плохо поддаются лечению традиционными фармакотерапевтическими методами, т.к. поступление лекарственных веществ в них затруднено. При электрофорезе же лекарственные вещества могут поступать в патологический очаг не только гематогенным, но и электрогенным путем.
4. При электрофорезе побочные и аллергические реакции наблюдаются во много раз реже, чем при пероральном или парентеральном применении этих же лекарств. Уменьшение или полное отсутствие побочных реакций при электрофорезе обусловлено рядом причин: невысокой концентрацией лекарства в крови; введением их в наиболее чистом виде; положительным влиянием физического фактора на общую реактивность и иммунобиологический статус организма и др
5. При электрофорезе в организм вводятся только те лекарственные ионы или ингредиенты лекарств, на терапевтическое действие которых рассчитывают. Противоионы и различные примеси, которые могут тормозить действие основного лекарственного иона, в организм при этом не попадают, а остаются на прокладке.
При электрофорезе в организм вводятся только те лекарственные ионы или ингредиенты лекарств, на терапевтическое действие которых рассчитывают. Противоионы и различные примеси, которые могут тормозить действие основного лекарственного иона, в организм при этом не попадают, а остаются на прокладке.
6. В соответствии с сущностью метода при электрофорезе в организм лекарства поступают в виде ионов. И это очень важно, т.к. в ионной форме лекарства значительно активнее, чем в молекулярной, в которой они вводятся при обычных способах их применения.
7. Многих пациентов, прежде всего детей, пожилых пациентов привлекает абсолютная безболезненность метода при его правильном проведении.
8. При лекарственном электрофорезе исключается введение в организм растворителя. Это немаловажное достоинство метода, ибо вводимый при других способах лекарственной терапии растворитель деформирует кожу, нарушает микроциркуляцию и метаболизм в ней, может служить причиной развития постинъекционных инфильтратов.
9. При всей важности приведенных выше особенностей метода все же основным достоинством лекарственного электрофореза, думается, является то, что лекарственное вещество здесь действует на фоне различных, имеющих терапевтическое значение изменений, вызываемых используемым электрическим током. Именно благодаря этому отчетливое специфическое и выраженное лечебное действие вводимых электрофорезом лекарств проявляется при более низких концентрациях, которые при обычных путях их введения были бы малоэффективны.
Продолжительность процедуры зависит от локализации воздействия и вида используемого тока. При общих и сегментарно-рефлекторных методиках она обычно не превышает 15-20 мин, а при местных процедурах – 30-40 мин. Использование флюктуирующих или синусоидальных модулированных токов (в выпрямленном режиме) требует некоторого уменьшения продолжительности лекарственного электрофореза, а при проведении его по методике электросна длительность воздействия, наоборот, обычно удлиняется. Курс лечения лекарственным электрофорезом в зависимости от тяжести состояния больного может быть различным по продолжительности: от 10-12 до 16-20 процедур, проводимых ежедневно или через день.
Курс лечения лекарственным электрофорезом в зависимости от тяжести состояния больного может быть различным по продолжительности: от 10-12 до 16-20 процедур, проводимых ежедневно или через день.
Показания:
Для лекарственного электрофореза определяются фармакотерапевтическими свойствами вводимого препарата, а также показаниями к использованию физического фактора (гальванического или других постоянных токов). В связи с широким перечнем лекарств, пригодных для электрофореза, и разнообразием используемых электрических токов показания для назначения метода весьма разнообразны.
В принципе трудно найти заболевание, при котором не мог бы быть назначен лекарственный электрофорез. Наиболее целесообразно лекарственный электрофорез применять при тех заболеваниях, при которых показаны как лекарственные вещества, так и используемый при этом электрический ток.
Противопоказания:
- индивидуальная непереносимость лекарственного вещества,
- противопоказания к использованию лекарства и самого электрического тока
Запись на процедуры в физиотерапевтическое отделение нашей поликлиники проводится в любое удобное для Вас время по телефонам:
(499) 262-11-29, 262-93-61
Перейти в разделы:
Физиотерапевтическое отделение>>
Водогрязелечение
Ванны
Душ
SPA-терапия
Иглорефлексотерапия
Косметология
показания и противопоказания, методики, алгоритм проведения, механизм действия, преимущества
Даже в небольшой поликлинике всегда можно найти кабинет для проведения физпроцедур. Но мало кто из среднестатистических пациентов владеет информацией обо всем спектре таких манипуляций, об их лечебных свойствах и пользе для здоровья. Многие даже скептически относятся к рекомендациям доктора заглянуть на физпроцедуры. И совершенно зря. Такое воздействие помогает в кратчайшие сроки справиться со многими нарушениями здоровья, сделать лечение более эффективным, предупредить осложнения и обострения хронических заболеваний. К самым популярным физпроцедурам можно, безусловно, отнести электрофорез, показания и противопоказания к проведению которого давайте рассмотрим, уточним, какая польза и вред могут быть человеку от таких процедур.
Но мало кто из среднестатистических пациентов владеет информацией обо всем спектре таких манипуляций, об их лечебных свойствах и пользе для здоровья. Многие даже скептически относятся к рекомендациям доктора заглянуть на физпроцедуры. И совершенно зря. Такое воздействие помогает в кратчайшие сроки справиться со многими нарушениями здоровья, сделать лечение более эффективным, предупредить осложнения и обострения хронических заболеваний. К самым популярным физпроцедурам можно, безусловно, отнести электрофорез, показания и противопоказания к проведению которого давайте рассмотрим, уточним, какая польза и вред могут быть человеку от таких процедур.
Электрофорез, как метод физиотерапевтического воздействия, подразумевает влияние на человеческий организм постоянных электрических импульсов, которые оказывают общий и местный терапевтический эффект. Такая процедура помогает ввести лекарственные средства через кожу, а также сквозь слизистые оболочки.
Электрофорез — польза и вред процедуры
Польза электрофореза
Данный метод физиотерапии считается одним из наиболее популярных. Он оказывает ряд положительных эффектов на организм пациента. Такие процедуры позволяют понизить интенсивность воспалительных процессов, они оказывают мягкое противоотечное действие и неплохо устраняют болевой синдром. Также электрофорез способствует расслаблению повышенного мышечного тонуса и отлично успокаивает. Его проведение положительно сказывается на процессах микроциркуляции, активизирует регенеративные процессы в тканях и запускает синтез ряда биологически активных веществ, представленных витаминами, микроэлементами, гормонами и пр. Кроме всего прочего данная процедура положительно сказывается на состоянии иммунитета.
Он оказывает ряд положительных эффектов на организм пациента. Такие процедуры позволяют понизить интенсивность воспалительных процессов, они оказывают мягкое противоотечное действие и неплохо устраняют болевой синдром. Также электрофорез способствует расслаблению повышенного мышечного тонуса и отлично успокаивает. Его проведение положительно сказывается на процессах микроциркуляции, активизирует регенеративные процессы в тканях и запускает синтез ряда биологически активных веществ, представленных витаминами, микроэлементами, гормонами и пр. Кроме всего прочего данная процедура положительно сказывается на состоянии иммунитета.
При проведении электрофореза лекарства проникают внутрь организма сквозь межклеточные пространства, а также через сальные и потовые железы, трансформируясь в положительные или отрицательные ионы. Большая часть медикамента задерживается в коже и в области подкожно-жировой клетчатки, проникая в кровоток лишь через сутки и более после такой процедуры. Благодаря этому электрофорез характеризуется пролонгированным действием.
Активные вещества при такой процедуре накапливаются в проблемном участке, ведь прокладку с медикаментом накладывают непосредственно на патологический очаг. Они существенно превышают дозировку, которую можно безопасно достичь при инъекционном или же пероральном введении. Лекарство минует пищеварительный тракт, благодаря чему не может вызвать большинства побочных эффектов.
Показания электрофореза
Электрофорез проводится как часть комплексного лечения при коррекции многих заболеваний разных органов и систем. Его используют в терапевтической, неврологической, хирургической, гинекологической, травматологической, стоматологической и педиатрической практике. При этом процедуры могут проводиться неоднократно.
Так электрофорез помогает пациентам с недугами дыхательной системы, представленными , и .
Ним лечат разные ЛОР-заболевания, такие процедуры показаны пациентам с , и .
Электрофорез может использоваться для терапии многих недугов пищеварительного тракта: гастрита, язвенного поражения желудка либо двенадцатиперстной кишки, холецистита, панкреатита и колита.
Такие процедуры показаны и при болезнях сердца и сосудов. Так электрофорез часто назначают пациентам с гипертонией (первой и второй степени), гипотонией, атеросклерозом, стенокардией, варикозным расширением вен, мерцательной аритмией и эндартериитом.
Проведение электрофореза показано при ряде недугов мочеполовой системы, представленных пиелонефритом, циститом, простатитом, эндометриозом, андекситом и эндометритом. Такие процедуры поспособствуют лечению цервицита и вагинита.
Еще электрофорез может принести пользу при многих заболеваниях нервной системы. Ним лечат невриты и невралгии, р
Комбинированные методы физиотерапии (физико-фармакологические воздействия)
Среди основных комбинированных методов физиотерапевтического воздействия на ткани и органы челюстно-лицевой области выделяют: электрофорез, ультрафонофорез, магнитофорез и лазерофорез.
Лекарственный электрофорез
Электрофорез (введение лекарственных веществ с помощью гальванического тока) через кожу и слизистые не отвечает законам химического электролиза, поскольку кожа проявляет свою электрохимическую активность. Электрохимический перенос веществ через кожу тормозится за счет:
Электрохимический перенос веществ через кожу тормозится за счет:
1) химической мантии – слоя кожного жира, пота и отшелушений;
2) поляризационными явлениями;
3) участия в электротринспорте более подвижных водородных и гидроксильных групп.
Принято считать, что при электрофорезе вводится от 3 до 10 % лекарственного вещества, нанесенного на прокладку.
Основными путями проникновения лекарств в организм при электрофорезе являются выводные протоки сальных и потовых желез. При электрофорезе лекарственных веществ через слизистую основным механизмом является чрезклеточное проникновение.
Экспериментальными исследованиями (В.С. Улащик, 1981 г.) установлено, что при электрофорезе вводимый препарат проникает на глубину 2-3 см.
Фармакодинамика вводимых при помощи электрофореза веществ имеет следующие особенности:
1) образование подслизистого депо;
2) избирательное накопление лекарственного препарата в тканях, относящихся к тому же метамеру, что и раздражаемая током поверхность.
Фонофорез
Фонофорез (введение лекарственного вещества с помощью ультразвука) осуществляется по тем же основным механизмам, что и электрофорез. Известную роль при этом играют чрезклеточные и межклеточные механизмы, так как одним из важнейших свойств ультразвука является деполимеризирующее и разрыхляющее действие, оказываемое им на костную ткань челюстей и слизистую оболочку альвеолярного отростка.
Глубина проникновения лекарственных препаратов при ультрафонофорезе незначительна и составляет 2-6 мм. (А.Ш. Лазаретник, 1961 г.), а количественное соотношение составляет 5-6 % от препарата, помещенного на прокладку. При чем количество вещества, проникающего через слизистую больше, чем при ультрафонофорезе через кожу.
Лазерофорез
Лазерофорез, как комплексный метод стимулирующей терапии, имеет в своей основе сочетанное воздействие двух основных факторов: воздействие самого лекарственного вещества и воздействие низкоинтенсивного лазерного излучения.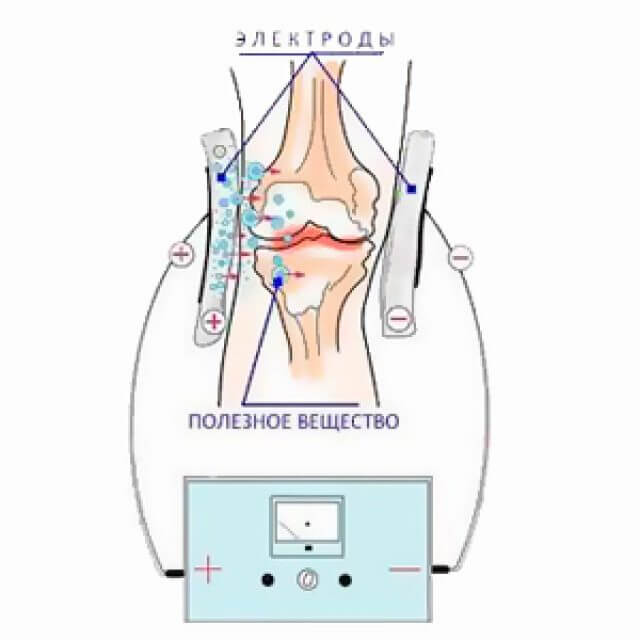 Этот метод получил в настоящее время широкое распространение в различных разделах медицины, и в том числе, в стоматологии. .
Этот метод получил в настоящее время широкое распространение в различных разделах медицины, и в том числе, в стоматологии. .
Механизм проникновения лекарственных веществ под воздействием НИЛИ чрезвычайно сложен, еще полностью не изучен. Установлено, что непосредственное воздействие НИЛИ на ткани происходит только до 0,8 см, а все эффекты, происходящие на большей глубине вызываются за счет опосредованных механизмов подлежащих тканей, находящихся под действием низкоинтенсивного лазерного излучения в «возбужденном состоянии». Механизм ЛФ рассматривается автором на клеточном и тканевом уровне. Было установлено, что НИЛИ стимулирует проникновение лекарственных препаратов за счет основных механизмов: повышения проницаемости слизистой оболочки полости рта; улучшения реологичеких свойств крови; повышения тканевого метаболизма; улучшения микроциркуляции; стимуляции кислородного обмена; активизации мембранных рецепторов «клеток-мишеней».
Методы физиотерапевтической оптимизации ортодонтического лечения зубочелюстных аномалий
Проблема сокращения сроков ортодонтического лечения решается несколькими путями: за счет стимуляции перемещения зубов во время активного периода, путем уменьшения плотности челюстных костей, и за счет укрепления новообразованной кости во время ретенционного периода.
Методы оптимизации активного периода ортодонтического лечения
С.И. Дорошенко и Н.В. Ращенко предлагают использовать вакуум-терапию для лечения ретенированных зубов, обусловленных наличием сверхкомплектных. Согласно разработанного ими способа после удаления сверхкомплектных зубов на альвеолярный отросток в области ретенированных зубов воздействуют вакуумом до появления гематомы в течение 2-3 мин. В результате осуществления данного способа обеспечивается восстановление зубного ряда с меньшей травматичностью.
Действие низкочастотной вибрации на ткани зуба и пародонта в процессе ортодонтического лечения изучали Г.И. Лютик, С.И. Дорошенко. Вибрация передается через зуб, на который прикладывается, по периодонту, вызывая эффект «насоса», стимулирующий гемодинамику, процессы тканевого обмена, а, следовательно, и активизирующий процессы резорбции и костеобразования. Использование вибрационного воздействия при ортодонтическом лечении аномалий положения зубов и зубочелюстных деформаций позволяет сократить сроки перемещения зубов в 1,5-2 раза.
Исследованиями, проведенными Ф. Х. Саидом, установлено, что такие методы стимулирующей терапии, как вибрационное воздействие, вакуум-терапия и магниторезонансная рефлексотерапия миллиметрового диапазона могут быть успешно применены как для стимуляции процесса прорезывания ретенированных зубов (сверхкомплектных и комплектных), так и для сокращения сроков аппаратурного лечения в 1,5-2 раза.
Свойства постоянного магнитного поля снижать воспалительные процессы, усиливать приток крови в зоне его действия, активизировать обменные процессы используют для стимуляции перемещения зубов Н.И. Гунько. Оптимальные параметры низкочастотного магнитного поля: магнитная индукция – 22 мТл, частота — 50 Гц, экспозиция — 18 мин., импульсный режим. Под влиянием такого поля происходит интенсификация репаративного остеогенеза, усиливается минерализация кости.
Е.Ю. Симановская и Л.М. Гвоздева при ортодонтическом лечении зубочелюстных аномалий у подростков применяли вакуум-терапию в сочетании с электрофорезом 2% -ного водного раствора хлористого лития (в течение 15-25 мин через день, число сеансов 5-10). Выбор для электрофоретического воздействия раствора хлористого лития основан на его высокой гигроскопичности, а также способности растворять минерализованные зубные отложения. Взаимодействие раствора хлористого лития, вводимого электрофоретическим путем, с минеральными компонентами костной ткани приводит к набуханию последней, что снижает резистентность, плотность кости, а, следовательно, делает ее более податливой для растяжения в период активного ортодонтического лечения. При этом в период деминерализации под действием очагового дозированного вакуума происходит усиление пенетрирующего эффекта электрофореза, что активизирует репаративные процессы в костной ткани и способствует направленной перестройке костных структур в области перемещаемых зубов.
Выбор для электрофоретического воздействия раствора хлористого лития основан на его высокой гигроскопичности, а также способности растворять минерализованные зубные отложения. Взаимодействие раствора хлористого лития, вводимого электрофоретическим путем, с минеральными компонентами костной ткани приводит к набуханию последней, что снижает резистентность, плотность кости, а, следовательно, делает ее более податливой для растяжения в период активного ортодонтического лечения. При этом в период деминерализации под действием очагового дозированного вакуума происходит усиление пенетрирующего эффекта электрофореза, что активизирует репаративные процессы в костной ткани и способствует направленной перестройке костных структур в области перемещаемых зубов.
С целью сокращения сроков ортодонтического лечения зубочелюстных деформаций А.В. Козел с соавт. проводили лекарственный электрофорез 0,5-3% — ным раствором трилона Б (курс 10 сеансов по 15 мин) в области подлежащих перемещению зубов. Указанное комплексное воздействие способствует местной декальцинации кости, а декальцинированная костная ткань легче резорбируется, чем не декальцинированная, в связи с чем, разработанную методику можно рекомендовать для ослабления костной ткани с целью последующего корпусного перемещения зубов.
Указанное комплексное воздействие способствует местной декальцинации кости, а декальцинированная костная ткань легче резорбируется, чем не декальцинированная, в связи с чем, разработанную методику можно рекомендовать для ослабления костной ткани с целью последующего корпусного перемещения зубов.
Для стимуляции ортодонтического лечения широко применяется лидаза. Известно, что действие Л направлено, прежде всего, на гиалуроновую кислоту. Л расщепляет ее до глюкозамина и глюкуроновой кислоты (Машковский М.Д). Соответственно изменяется структура протеогликанов, они распадаются на мономеры, что отражается на организации межклеточного вещества костной ткани. Доказано, что протеогликаны, составляющие 10% всех неколлагеновых белков матрикса кости, играют важную роль в развитии и функционировании костной ткани.
Боровский Е.В. выявил, что под воздействием лидазы повышается проницаемость эмали и дентина.
И.Е. Андросовой с соавт. изложены основы механизма действия препаратов гиалуронидазного действия как факторов, стимулирующих перемещение или прорезывание аномалийно расположенных зубов. Авторы полагают, что для формирования грануляционно-фиброзной ткани, на основе которой формируется кость альвеолярного отростка и альвеолы прорезавшегося постоянного зуба, необходимым условием является образование гранулемы. В этом процессе основной действующей «фигурой» становится молодой фибробласт, интенсивно синтезирующий мукополисахариды, а затем белок — коллаген. Авторы рекомендуют стимулировать процессы прорезывания постоянных зубов ферментами гиалуронидазного действия.
Авторы полагают, что для формирования грануляционно-фиброзной ткани, на основе которой формируется кость альвеолярного отростка и альвеолы прорезавшегося постоянного зуба, необходимым условием является образование гранулемы. В этом процессе основной действующей «фигурой» становится молодой фибробласт, интенсивно синтезирующий мукополисахариды, а затем белок — коллаген. Авторы рекомендуют стимулировать процессы прорезывания постоянных зубов ферментами гиалуронидазного действия.
Эффективный метод стимулирования самостоятельного прорезывания ретенированных зубов, путем применения вибрационного воздействия и вакуум-терапии разработан С.И. Дорошенко. В комплексе воздействия с физическими факторами авторами применялся ультрафонофорез хонсурида и лидазы. Такое введение лекарственных веществ позволяет локально депонировать их в тканях, пролонгировать их действие, уменьшить дозу и снизить концентрацию в крови. Применение медикаментозного воздействия в комплексе с ФФ позволило существенно сократить сроки ортодонтического лечения больных с ретенированными зубами.
Механизм электрофореза лидазы А.И. Перцовским с соавт. Автором in vivo в эксперименте на бедренной мышце кролика установлено, что под воздействием гальванического тока гиалуронидаза проникает в глубь мышечной ткани и сохраняется в течение 12 – 24 часов.
Кроме того, установлено, что гиалуронидаза теряет свою ферментативную активность при повышении температуры среды, в которую помещен фермент, гальванического тока и ультразвука.
С целью оптимизации аппаратурно-хирургического метода лечения зубочелюстных аномалий, на кафедре разработан ускоренный комплексный метод лечения больных с использованием лазерофореза лидазы. Лазерофорез проводили по следующей методике: порошок лидазы (64 ЕД) растворяли в 2 мл лидокаина. Полученным раствором инфильтрировали переходную складку в области проекции ретенированного зуба или проекции корня аномалийно расположенного зуба, создавая подслизистое депо лекарственного вещества (рис. 191).
Затем, контактным методом проводили облучение ГНЛ. При этом использовали аппарат УФЛ — 1 типа ЛГ-75, со световодом типа «кварц-полимер» и насадки (рис. 192). Насадки прикладывали к слизистой оболочке по переходной складке (рис. 193).
При этом использовали аппарат УФЛ — 1 типа ЛГ-75, со световодом типа «кварц-полимер» и насадки (рис. 192). Насадки прикладывали к слизистой оболочке по переходной складке (рис. 193).
Лечение больных проводилось по следующей схеме:
1. Проведение 7 процедур дооперационного этапа лазерофореза через день. Параметры излучения: длина волны – 0,63 мкм, плотность мощности – 200 мВт/см2, экспозиция 10 мин.
2. Изготовление ортодонтического аппарата.
3. Припасовка и сдача ортодонтического аппарата.
4. Проведение оперативного вмешательства: компактостеотомия в области аномалийно расположенного зуба (зубов).
5. Проведение послеоперационного этапа лазерофореза на область компактостеотомии с теми же параметрами излучения. Количество процедур 2 курса по 7 процедур через день. Между курсами ЛФЛ перерыв не менее 1 недели.
6. Проведение активного периода ортодонтического лечения.
7. Проведение ретенционного периода ортодонтического лечения.
Разработанный метод существенным образом улучшает динамику лечебного процесса и позволяет сократить сроки лечения в среднем в 1,5 разы за счет сохранения ферментативной активности лидазы, локального повышения порозности костной ткани челюстей и замедления репаративной регенерации на стадии фазы формирования тканеспецифических структур. Это проявляется сокращением инерционного периода от момента приложения силы ортодонтического аппарата к аномалийно расположенного зубу до начала его перемещения, сокращением срока активного периода лечения при неизменном сроке ретенционного периода.
Методы оптимизации ретенционного периода ортодонтического лечения
Я.Г. Бик обосновал индуцирующее влияние метилурацила на регенерацию костной ткани пародонта. Метилурацил влияет на остеогенную дифференцировку соединительно-тканных клеток, стимулирует коллагенез и остеогенез. С целью регенерации костной ткани после ортодонтического лечения автор применял лекарственные препараты, стимулирующие синтез белка. Важным стимулирующим действием на синтез белков в организме обладают анаболические стероиды – фенаболил, ретаболил, а так же настероидные антибиотики – калия ортотат. Кроме того, эти вещества оказывают положительное влияние на азотистый обмен и способствуют фиксации кальция в костях, ускоряют кальцификацию костной ткани при локальном остеопорозе, вызванном ортодонтическим лечением. Для оптимизации этих процессов широко применяют комплексы микроэлементов, содержащих кальций, фосфор и магний.
С целью регенерации костной ткани после ортодонтического лечения автор применял лекарственные препараты, стимулирующие синтез белка. Важным стимулирующим действием на синтез белков в организме обладают анаболические стероиды – фенаболил, ретаболил, а так же настероидные антибиотики – калия ортотат. Кроме того, эти вещества оказывают положительное влияние на азотистый обмен и способствуют фиксации кальция в костях, ускоряют кальцификацию костной ткани при локальном остеопорозе, вызванном ортодонтическим лечением. Для оптимизации этих процессов широко применяют комплексы микроэлементов, содержащих кальций, фосфор и магний.
Индуцирующим влиянием на регенерацию тканей пародонта в области перемещаемых зубов обладают производные пиримидиновых оснований, в частности, метилурацил. Он оптимизирует остеогенную дифференцировку соединительнотканных клеток, стимулирует остеогенез.
Zhou D. экспериментально обосновал возможность применения индометацина для ингибирования остеопороза после ортодонтической компрессии.
Ортодонтия
Под редакцией проф. В.И. Куцевляка
Механизм действия: гормоны с внутриклеточными рецепторами
Механизм действия: гормоны с внутриклеточными рецепторами
Рецепторы стероидных и тироидных гормонов расположены внутри клеток-мишеней, в цитоплазме или ядре, и функционируют как лиганд-зависимых факторов транскрипции . То есть комплекс гормон-рецептор связывается с промоторными областями чувствительных генов и стимулирует, а иногда и ингибирует транскрипцию этих генов.
Таким образом, механизм действия стероидных гормонов заключается в модуляции экспрессии генов в клетках-мишенях.Путем избирательного воздействия на транскрипцию из батареи генов изменяется концентрация этих соответствующих белков, что, несомненно, может изменить фенотип клетки.
Структура внутриклеточных рецепторов
Рецепторы стероидов и гормонов щитовидной железы являются членами большой группы («надсемейства») факторов транскрипции. В некоторых случаях в клетках экспрессируются несколько форм данного рецептора, что усложняет ответ. Все эти рецепторы состоят из единой полипептидной цепи, которая, при упрощенном анализе, состоит из трех отдельных доменов:
- Аминоконец : В большинстве случаев эта область участвует в активации или стимуляции транскрипции, взаимодействуя с другими компонентами транскрипционного аппарата.Последовательность сильно варьирует среди разных рецепторов.
- ДНК-связывающий домен : аминокислоты в этой области отвечают за связывание рецептора со специфическим
последовательности ДНК. - Карбокси-конец или лиганд-связывающий домен : это область, которая связывает гормон.
В дополнение к этим трем коровым доменам, две другие важные области рецепторного белка представляют собой последовательность ядерной локализации, которая направляет белок в ядро, и домен димеризации, который отвечает за фиксацию двух рецепторов вместе в форме
способен связывать ДНК.
Связывание гормональных рецепторов и взаимодействие с ДНК
Являясь липидами, стероидные гормоны проникают в клетку путем простой диффузии через плазматическую мембрану. Гормоны щитовидной железы попадают в клетку путем облегченной диффузии. Рецепторы существуют либо в цитоплазме, либо в ядре, где они встречаются с гормоном. Когда гормон связывается с рецептором, происходит характерная серия событий:
- Активация рецептора — это термин, используемый для описания конформационных изменений рецептора, вызванных связывающим гормоном.Основным следствием активации является то, что рецептор становится компетентным связывать ДНК.
- Активированные рецепторы связываются с «элементами гормонального ответа» , которые представляют собой короткие специфические последовательности ДНК, расположенные в промоторах гормонально-чувствительных генов. В большинстве случаев комплексы гормон-рецептор связывают ДНК парами, как показано на рисунке ниже.
- Транскрипция тех генов, с которыми связан рецептор, нарушена. Чаще всего связывание рецептора стимулирует транскрипцию.Таким образом, комплекс гормон-рецептор функционирует как фактор транскрипции.
Как и следовало ожидать, существует ряд вариантов описанных выше тем в зависимости от конкретного рассматриваемого рецептора. Например, при отсутствии гормона некоторые внутриклеточные рецепторы действительно связывают свои гормональные ответные элементы слабо и подавляют транскрипцию, но, когда они образуют комплекс с гормоном, активируются и сильно стимулируют транскрипцию. Некоторые рецепторы связывают ДНК не с другим в своем роде, а с другим внутриклеточным рецептором.
В качестве конкретного примера рассмотрим глюкокортикоиды, тип стероидного гормона, который, вероятно, влияет на физиологию всех клеток организма. На изображении справа изображена пара
рецепторы глюкокортикоидов (синий и зеленый вверху), связанные с их ответным элементом гормона ДНК (внизу). На этом изображении не видны два стероидных гормона.
Консенсусная последовательность элемента гормонального ответа в этом случае (называемого элементом глюкокортикоидного ответа ) — это GGTACANNNTGTTCT, где N — любой нуклеотид.
Отправляйте комментарии [email protected]
Гормоны с рецепторами клеточной поверхности
Механизм действия: гормоны с рецепторами клеточной поверхности
Белковые и пептидные гормоны, катехоламины, такие как адреналин, и эйкозаноиды, такие как простагландины, обнаруживают свои рецепторы, украшающие плазматическую мембрану клеток-мишеней.
Связывание гормона с рецептором запускает серию событий, которые приводят к генерации так называемых вторичных мессенджеров внутри клетки (гормон является первым мессенджером).Затем вторые мессенджеры запускают серию молекулярных взаимодействий, которые изменяют физиологическое состояние клетки. Другой термин, используемый для описания всего этого процесса, — это преобразование сигнала .
Структура рецепторов клеточной поверхности
Рецепторы клеточной поверхности представляют собой интегральные мембранные белки и, как таковые, имеют области, которые участвуют в трех основных доменах:
- Внеклеточные домены: Некоторые остатки, находящиеся вне клетки, взаимодействуют с гормоном и связывают его — другим термином для этих областей является лиганд-связывающий домен .
- Трансмембранные домены: Гидрофобные участки аминокислот «удобны» в липидном бислое и служат для закрепления рецептора в мембране.
- Цитоплазматические или внутриклеточные домены: Хвосты или петли рецептора, находящиеся в цитоплазме, реагируют на связывание гормона, каким-либо образом взаимодействуя с другими молекулами, что приводит к образованию вторичных мессенджеров. Таким образом, цитоплазматические остатки рецептора представляют собой эффекторную область молекулы.
Было идентифицировано несколько отличительных вариаций в структуре рецептора. Как показано ниже, некоторые рецепторы представляют собой простые однопроходные белки; многие рецепторы факторов роста принимают эту форму. Другие, например рецептор инсулина, содержат более одной субъединицы. Другой класс, который включает бета-адренорецепторы, проходит через мембрану семь раз.
Рецепторные молекулы не изолированы сами по себе и не фиксируются в одном месте плазматической мембраны.В некоторых случаях другие интегральные мембранные белки взаимодействуют с рецептором, чтобы модулировать его активность. Некоторые типы рецепторов группируются в мембране после связывания гормона. Наконец, как подробно описано ниже, взаимодействие гормонально-связанного рецептора с другими мембранными или цитоплазматическими белками является ключом к генерации вторичных мессенджеров и передаче гормонального сигнала.
Системы вторых сообщений
Подумайте, что бы произошло, если бы поздно ночью вы заметили горящее здание. Надеюсь, вы наберете 911 или аналогичный номер службы экстренной помощи. Вы проинформируете диспетчера о пожаре, а диспетчер, в свою очередь, свяжется с несколькими пожарными и «активирует» их. Затем пожарные быстро приступили к работе, поливая огонь водой, устанавливая заграждения на дорогах и тому подобное. Они также, вероятно, будут активировать других «игроков», таких как полиция и следователи по пожарной безопасности, которые придут позже, чтобы попытаться определить причину пожара. Важно отметить, что как только пожар потушен (или здание полностью разрушено), пожарные возвращаются на станцию и ложатся спать.
Реакция сообщества на пожар, по крайней мере в некотором смысле, аналогична системе вторичных посыльных, участвующей в действии гормона. В описанном сценарии вы — «первый посланник», диспетчер — «приемник», пожарные — «вторые посланники».
В настоящее время в ячейках распознаются четыре системы вторичного обмена сообщениями, как показано в таблице ниже. Обратите внимание, что не только несколько гормонов используют одну и ту же систему вторичных мессенджеров, но и один гормон может использовать более одной системы. Понимание того, как клетки объединяют сигналы нескольких гормонов в согласованный биологический ответ, остается проблемой.
| Второй посланник | Примеры гормонов, которые используют эту систему |
|---|---|
| Циклический усилитель | Адреналин и норадреналин, глюкагон, лютеинизирующий гормон, фолликулостимулирующий гормон, тиреотропный гормон, кальцитонин, паратиреоидный гормон, антидиуретический гормон |
| Активность протеинкиназы | Инсулин, гормон роста, пролактин, окситоцин, эритропоэтин, несколько факторов роста |
| Кальций и / или фосфоинозитиды | Эпинефрин и норадреналин, ангиотензин II, антидиуретический гормон, гонадотропин-рилизинг-гормон, тироид-рилизинг-гормон. |
| Циклический GMP | Натуральный гормон предсердия, оксид азота |
Во всех случаях кажущийся слабым сигнал, генерируемый связыванием гормона с его рецептором, усиливается внутри клетки в каскад действий, которые изменяют физиологическое состояние клетки. Ниже представлены два примера систем вторичных посыльных, обычно используемых гормонами. В качестве примеров используются глюкагон и инсулин, оба из которых в конечном итоге работают через молекулярный переключатель, включающий фосфорилирование белка.Имейте в виду, что в обоих случаях очень сложная система значительно упрощается.
Cyclic AMP Second Messenger Systems
Циклический аденозинмонофосфат (цАМФ) представляет собой нуклеотид, вырабатываемый из АТФ под действием фермента аденилатциклазы. Внутриклеточная концентрация цАМФ увеличивается или уменьшается за счет различных гормонов, и такие колебания влияют на множество клеточных процессов. Одним из заметных и важных эффектов повышенных концентраций цАМФ является активация цАМФ-зависимой протеинкиназы, называемой протеинкиназой А.
Протеинкиназа А номинально находится в каталитически неактивном состоянии, но становится активной, когда связывает цАМФ. После активации протеинкиназа А фосфорилирует ряд других белков, многие из которых сами являются ферментами, которые либо активируются, либо подавляются путем фосфорилирования. Такие изменения ферментативной активности внутри клетки явно изменяют ее состояние.
Теперь давайте соберем эту информацию, чтобы понять механизм действия гормона, такого как глюкагон:
- Глюкагон связывает свой рецептор в плазматической мембране клеток-мишеней (например,грамм. гепатоциты).
- Связанный рецептор взаимодействует и через набор G-белков включает аденилатциклазу, которая также является интегральным мембранным белком.
- Активированная аденилатциклаза начинает преобразовывать АТФ в циклический АМФ, что приводит к повышению внутриклеточной концентрации цАМФ.
- Высокие уровни цАМФ в цитозоле делают вероятным, что протеинкиназа А будет связываться цАМФ и, следовательно, каталитически активна.
- Активная протеинкиназа A «бегает по клетке», добавляя фосфаты к другим ферментам, тем самым изменяя их конформацию и модулируя их каталитическую активность — — — abracadabra, клетка изменилась!
- Уровни цАМФ снижаются из-за разрушения цАМФ-фосфодиэстеразой и инактивации аденилатциклазы.
В приведенном выше примере действие гормона заключалось в изменении активности ранее существовавших компонентов в клетке. Повышение уровня цАМФ также оказывает важное влияние на транскрипцию определенных генов.
Системы второго мессенджера тирозинкиназы
Рецепторы некоторых белковых гормонов сами по себе являются протеинкиназами, которые активируются связыванием гормона. Киназная активность, связанная с такими рецепторами, приводит к фосфорилированию остатков тирозина на других белках.Инсулин является примером гормона, рецептором которого является тирозинкиназа.
Гормон связывается с доменами, экспонируемыми на поверхности клетки, что приводит к конформационному изменению, которое активирует киназные домены, расположенные в цитоплазматических областях рецептора. Во многих случаях рецептор фосфорилируется как часть процесса активации киназы. Активированный рецептор фосфорилирует множество внутриклеточных мишеней, многие из которых являются ферментами, которые активируются или инактивируются при фосфорилировании.
Рисунок справа предназначен для изображения рецептора тирозинкиназы, подобного тому, который используется в инсулине. После связывания гормона рецептор претерпевает конформационные изменения, фосфорилирует себя, а затем фосфорилирует множество внутриклеточных мишеней. |
Как было показано с системами вторичного мессенджера цАМФ, активация рецепторных тирозинкиназ приводит к быстрой модуляции ряда целевых белков внутри клетки.Интересно, что некоторые из мишеней рецепторных киназ представляют собой протеинфосфатазы, которые после активации рецепторной тирозинкиназой становятся способными удалять фосфаты из других белков и изменять их активность. Опять же, кажущееся незначительное изменение из-за связывания гормонов усиливается во множестве эффектов внутри клетки.
В некоторых случаях связывание гормона с поверхностным рецептором вызывает каскад тирозинкиназы, даже если рецептор сам по себе не является тирозинкиназой. Рецептор гормона роста является одним из примеров такой системы — взаимодействие гормона роста с его рецептором приводит к активации цитоплазматических тирозинкиназ с результатами, концептуально аналогичными результатам, наблюдаемым в случае рецепторных киназ.
Судьба гормонально-рецепторного комплекса
Нормальное функционирование ячейки зависит от каскадов вторичных мессенджеров, являющихся переходными событиями. Действительно, ряд раковых заболеваний связан с рецепторами, которые постоянно стимулируют системы вторичных мессенджеров. Одной из важных составляющих негативного регулирования действия гормонов является интернализация рецепторов клеточной поверхности. Во многих случаях интернализация стимулируется связыванием гормонов.
Интернализация происходит за счет эндоцитоза через структуры, называемые покрытыми ямками.Образующиеся эндосомы (иногда называемые «рецептосомами») могут сливаться с лизосомами, что приводит к разрушению рецептора и гормона. В других случаях оказывается, что гормон диссоциирует и рецептор повторно используется путем слияния эндосомы обратно в плазматическую мембрану.
Отправляйте комментарии [email protected]
Профилактические и профилактические механизмы действия биоактивных компонентов граната
Плоды граната обладают сильными противовоспалительными, антиоксидантными, противодействующими ожирению и противоопухолевыми свойствами, что с древних времен привело к росту его популярности в качестве функционального пищевого и нутрицевтического источника.Его можно разделить на три части: семена, кожуру и сок, каждая из которых имеет лечебные свойства. В нескольких исследованиях исследуются его биоактивные компоненты как средства, позволяющие связать их с определенным положительным эффектом и разрабатывать будущие продукты и терапевтические применения. Многие полезные эффекты связаны с присутствием эллаговой кислоты, эллагитаннинов (включая пуникалагины), пунической кислоты и других жирных кислот, флавоноидов, антоцианидинов, антоцианов, эстрогенных флавонолов и флавонов, которые являются его наиболее терапевтически полезными компонентами.Однако синергетическое действие компонентов граната, по-видимому, превосходит по сравнению с отдельными составляющими. Многообещающие результаты были получены при лечении некоторых заболеваний, включая ожирение, инсулинорезистентность, воспаление кишечника и рак. Хотя умеренное потребление граната не приводит к побочным эффектам, необходимы будущие исследования для оценки безопасности и потенциального взаимодействия с лекарствами, которые могут изменить биодоступность биологически активных компонентов граната, а также лекарств.Цель этого обзора — обобщить эффекты для здоровья и механизмы действия экстрактов граната при хронических воспалительных заболеваниях.
1. Введение
Гранат ( Punica granatum ), древний плодоносящий листопадный кустарник, является преобладающим представителем двух видов, входящих в семейство Punicaceae [1]. Это уроженец Гималаев на севере Индии, но его выращивали и натурализовали на всем Ближнем Востоке, во всем европейском Средиземноморском регионе, в более засушливых частях Юго-Восточной Азии, в северной и тропической Африке и в некоторой степени в Соединенных Штатах, в частности Калифорния и Аризона [2].Плод граната — это большая ягода, характеризующаяся наличием тонких мембран внутри, которые позволяют взвесить множество плодов, каждая из которых окружена мешочками, содержащими сок. Плод можно разделить на три части: семена и сок, которые составляют примерно 3 и 30% веса плода, соответственно, и кожуру, которая включает упомянутую внутреннюю сеть мембран [1] с различным химическим составом и потенциалом. медицинские пособия.
Экстракты граната использовались с древних времен для лечения ряда заболеваний, включая паразитарные и микробные инфекции, диарею, язвы, афты, кровотечения и респираторные осложнения [3, 4].Современные применения включают заместительную гормональную терапию и гигиену полости рта, а также лечение подавления иммунитета и сердечно-сосудистых осложнений [5]. Кроме того, в настоящее время исследуются другие терапевтические свойства, такие как противоопухолевые, противовоспалительные, противовирусные, антибактериальные, противодиарейные и против ожирения. Хотя гранат употребляли и использовали в качестве лечебной пищи на Ближнем Востоке в течение тысяч лет, недавно он приобрел популярность в Соединенных Штатах [6]. Высокая антиоксидантная активность граната по сравнению с другими фруктами и антиоксидантными напитками, наряду с его противовоспалительными свойствами и свойствами против ожирения, повысила интерес к исследованию его потенциальных нутрицевтических и функциональных пищевых применений.Количество научных публикаций о гранате и его полезных для здоровья свойствах значительно увеличилось за последнее десятилетие. Поиск в PubMed от 23 ноября 2012 г. обнаружил 677 публикаций, связанных с гранатом, 619 из которых были опубликованы за последние десять лет и 425 — за последние пять. Кроме того, несколько продуктов, содержащих гранат, в том числе 100% соки, напитки, содержащие гранат, жидкие и порошкообразные полифенольные экстракты граната, а также средства по уходу за кожей, содержащие экстракты граната или масла семян, были недавно представлены на рынке США и рекламируются. за их благотворное влияние на здоровье [6].Таким образом, изучение биоактивных компонентов граната и их связывание с конкретными механизмами действия и воздействием на здоровье открывает перспективы для будущего терапевтического развития натуральных продуктов, полученных из граната. Цель этого обзора — обобщить эффекты для здоровья и механизмы действия экстрактов граната при хронических воспалительных заболеваниях.
2. Химический состав граната
За последнее десятилетие был достигнут значительный прогресс в изучении механизмов действия компонентов граната на здоровье и болезни.Похоже, что экстракты всех различных частей фруктов обладают лечебными свойствами [1]. Интересно, что недавние исследования сообщают, что даже кора, корни и листья растения граната также обладают лечебными свойствами [7]. Целью большинства недавних исследований граната является определение его терапевтических компонентов. Эллаговая кислота, эллагитаннины (включая пуникалагины), пуническая кислота, флавоноиды, антоцианидины, антоцианы, эстрогенные флавонолы и флавоны, по-видимому, являются наиболее терапевтически полезными компонентами граната [8], особенно эллагитаннины, которые выделяют эллаговую кислоту при гидролизе.Таблицы 1, 2, 3 и 4 суммируют химический состав каждой из различных частей плодов граната.
| |||||||||||||||||||||||||||||||||||||||||||
Границы | От молекулярных сигнатур до предиктивных биомаркеров: моделирование патофизиологии болезней и механизма действия лекарств
Введение
Несмотря на постоянно растущее число клинических испытаний, скорость доставки новых лекарств в клинику замедляется (Pammolli et al., 2011). Здесь профилирование Omics и высокопроизводительные технологии скрининга лекарств на стыке крупномасштабных клинических данных запустили новые концептуальные стратегии, направленные на улучшение стратификации пациентов для обеспечения точной медицины (Trusheim et al., 2011; Hollebecque et al., 2014). Для реализации таких подходов необходимо решить ряд вопросов, в том числе: (i) отражение клинической категоризации фенотипа на описании на молекулярном уровне, (ii) определение молекулярных факторов, механически управляющих прогрессированием заболевания, (iii) медикаментозное вмешательство, конкретно направленное на такие механизмы прогрессирования и (iv) прогностические биомаркеры, позволяющие проводить анализ соответствия назначению относительно соответствия релевантной патофизиологии и механизма действия лекарства на уровне отдельного пациента (Heinzel et al., 2012).
Клинически хорошо установленный пример — HER2-положительный рак молочной железы, характеризующийся сверхэкспрессией члена семейства рецепторов эпидермального фактора роста (ERBB2), играющего механистическую роль в прогрессировании заболевания. В случае, если фактор оказывается положительным для пациента, конкретная картина может быть применена для лечения, направленного на подавление сигналов роста (Hicks and Kulkarni, 2008). Тем не менее, клиническая картина рака молочной железы показывает гетерогенную патофизиологию, не считая подтипов, положительных по HER2.Следовательно, для комплексной оценки фенотипов прогрессирующего рака молочной железы необходимы мультимаркерные панели, например, реализованные с помощью мультиплексного анализа, содержащего 70 индивидуальных молекулярных характеристик (Buyse et al., 2006). Такие многомаркерные панели в целом стали многообещающей стратегией для характеристики сложных клинических проявлений, например, с использованием панели сывороточных маркеров для прогнозирования ишемической болезни сердца у пациентов с симптомами или профиля протеомики мочи для ранней диагностики диабетической болезни почек (LaFramboise et al., 2012; Zürbig et al., 2012).
Неспособность идентифицировать один причинный фактор в качестве заместителя для определения прогрессирования сложного клинического фенотипа становится очевидной при сравнении эффективности панелей маркеров с отдельными маркерами, с последними, например, в обзоре Hellemons et al. для начала и прогрессирования диабетической болезни почек (Hellemons et al., 2012). В клинической практике может использоваться другой тип биомаркера, обеспечивающий считывание фенотипа, в первую очередь отражающее функциональный статус органа в отличие от патофизиологических характеристик.При заболевании почек такие функциональные маркеры используются в ведении пациентов, а также в дизайне клинических исследований, включая расчетную скорость клубочковой фильтрации (рСКФ) и протеинурию (отражающие клубочковую фильтрацию и проникновение макромолекул через стенку капилляров клубочков, соответственно).
Связь этих параметров с обострением диабетической болезни почек, а также с увеличением частоты конечных точек, таких как сердечно-сосудистые события, неоспорима (Adler et al., 2003). Однако эти маркеры не дают информации о конкретных молекулярных характеристиках заболевания.Функциональные маркеры делают невозможным расслоение для индивидуализированной терапии в концепции точной медицины.
Молекулярный путь, представляющий наибольший интерес в нынешних клинических условиях диабетической болезни почек, — это ренин-ангиотензиновая система (РАС), по своей активности, прежде всего, контролируя кровяное давление и баланс жидкости. Блокада РАС помогла снизить частоту почечных событий у пациентов с сахарным диабетом и без него (Ruggenenti et al., 1998; Brenner et al., 2001). В исследовании Lewis et al. Блокада рецепторов ангиотензина ирбесартаном снижает риск первичной комбинированной конечной точки (удвоение исходной концентрации креатинина в сыворотке, развитие терминальной стадии почечной недостаточности или смерть от любой причины) в течение периода наблюдения продолжительностью 2,6 года на 20% по сравнению с плацебо (Льюис и др., 2001). Тем не менее, 50% пациентов в группе ирбесартана достигли первичной конечной точки через 54 месяца. В целях повышения эффективности терапии антагонистами РАС блокатор рецепторов ангиотензина был объединен с плацебо или ингибитором ангиотензинпревращающего фермента (АПФ) лизиноприлом (Fried et al., 2013). Комбинированная терапия не снизила частоту комбинированной конечной точки для почек. Напротив, наблюдался повышенный риск гиперкалиемии и острого повреждения почек, что подтверждает другие сообщения, ставящие под сомнение безопасность этого подхода (Mann et al., 2008; Parving et al., 2008).
Наряду с РАС, органоспецифические молекулярные процессы, включающие воспаление и окислительный стресс, были вовлечены в прогрессирующий тубулоинтерстициальный фиброз, лучший гистологический, а следовательно, молекулярный механистический предиктор неблагоприятного прогноза почечной болезни (Rodríguez-Iturbe and García García, 2010).Бардоксолон, активатор фактора 2 ядерного фактора, связанного с эритроидом-2, с антиоксидантной способностью увеличивал рСКФ у пациентов с запущенным диабетическим заболеванием почек (Pergola et al., 2011). Однако крупное проспективное контролируемое рандомизированное исследование с твердыми конечными точками пришлось прекратить из-за серьезных побочных эффектов (De Zeeuw et al., 2013).
Как показано в этих примерах для хронической болезни почек (но в своей концептуальной основе, верной для множества широко распространенных хронических заболеваний), многие из недавних интервенционных исследований не достигли своих целей.Здесь биомаркеры обещают сыграть ключевую роль в отборе пациентов для исследований и / или в прогнозировании долгосрочного воздействия препарата на точные конечные точки. Предварительная стратификация в рандомизированных контролируемых испытаниях путем разделения пациентов по ответу на лекарства, измеряемых биомаркерами, выступающими в качестве суррогата конечной точки, с последующей рандомизацией групп по отдельности является подходом, который, по крайней мере со статистической точки зрения, предпочтительнее, чем анализ post-hoc ( Де Леон, 2012). Такая стратегия обогащения в настоящее время e.g., протестировано в исследовании SONAR (ссылка на Clinicaltrials.gov NCT01858532), посвященном диабетической нефропатии (DN).
Однако в отношении соответствия конкретных лекарств биомаркеры должны иметь прогностическую ценность, т. Е. Биомаркеры должны на уровне конкретного пациента определять респондентов, получающих пользу от эффекта лекарства. При этом необходимо учитывать различные уровни, включая генетические и экологические компоненты, определяющие проявление болезни и ее прогрессирование. В мишени лекарственного средства может быть обнаружен генетический полиморфизм, влияющий на связывание лекарственного средства, но полиморфизм может дополнительно включать транспорт и метаболизм лекарственного средства (Johnson, 2001).Тем временем значительная часть генетических полиморфизмов стала релевантной этикетке лекарственного средства с точки зрения его эффективности, а также токсичности и побочных эффектов (Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США, 2014). Фармакогеномика ясно продемонстрировала, что генетический фон индивидуума вносит неоднородность в ответ на лекарственный препарат.
Тем не менее, этот параметр предполагает однородную популяцию пациентов в отношении молекулярно-механистических факторов, определяющих прогрессирование заболевания, только демонстрируя различия в генетических особенностях одного и того же молекулярно-механистического контекста.В таких условиях функциональные биомаркеры оказываются достаточными для выявления прогрессирующего заболевания, а вариативность лекарственного средства полностью объясняется генетическим фоном в отношении механизма действия конкретного лекарственного средства.
Дополнительная точка зрения может заключаться в том, что молекулярный механистический фон и относящиеся к прогрессированию молекулярные факторы per se разнообразны и зависят от пациента, естественно определяя лекарственный ответ (Mayer et al., 2012). В таком сценарии биомаркер должен служить представителем ключевых механистических факторов, характеризующих и управляющих заболеванием на уровне конкретного пациента, в сочетании с обучением конкретному вмешательству механизма заболевания в механизм действия лекарства.Для определения этих ограничений необходима подробная молекулярная карта клинического фенотипа и его влияния на механизм действия лекарства, и здесь интеграция профилей Omics помогает идентифицировать такие механизмы (Fechete et al., 2011; Mühlberger et al., 2012) .
Априорная стратификация пациентов на основе правильно подобранной панели биомаркеров, отражающих патофизиологию данного пациента (группы), позволяющая определить соответствие с механизмом действия конкретного препарата, представляется перспективным подходом.Как недавно обсуждалось Himmelfarb et al. свежие подходы имеют решающее значение в поиске методов лечения заболеваний почек, приносящих пользу пациентам, подчеркивая важность улучшения трансляционного аспекта в клинических исследованиях (Himmelfarb and Tuttle, 2013). В данном случае омикс-технологии значительно расширили ландшафт данных, характеризующих хроническое заболевание почек, однако, в первую очередь, в основном расширив набор кандидатов, очевидно релевантных процессов и путей, наряду с большим количеством кандидатов в биомаркеры, которые индивидуально препятствуют клинически значимым оценка прогрессирования заболевания (Fechete et al., 2011; Hellemons et al., 2012).
Интегративные подходы в области системной биологии были предложены для достижения консенсуса в описании патофизиологии хронического заболевания почек, включая молекулярные модели DN, а также рено-кардиальной оси (He et al., 2012; Komorowsky et al., 2012; Mayer et al., 2012; Heinzel et al., 2013). Тем не менее, процесс трансляции должен быть соблюден, объединяя патофизиологию болезней, маркеры стратификации, позволяющие стратегии обогащения, в сочетании с подходящими лекарствами на молекулярно-механистическом уровне для обеспечения точной медицины (Mirnezami et al., 2012). В этой работе мы приводим пример такой процедуры на DN, являющейся основной клинической картиной, ведущей к терминальной стадии почечной недостаточности.
Материалы и методы
Общие источники данных
гены, кодирующие белки, идентифицированные как связанные с DN, были собраны из общедоступных источников данных транскриптомики, дополненных молекулярными характеристиками, сообщающими о такой ассоциации в научной литературе. Молекулярные сигнатуры, информирующие о механизме действия ингибитора АПФ, были извлечены из общедоступных источников транскриптомики.Белки, обсуждаемые как биомаркеры или кандидаты в лекарственные мишени в контексте DN, были извлечены из научной литературы, а набор мишеней был дополнительно расширен за счет известных лекарственных мишеней для лекарств, которые в настоящее время используются в клинических испытаниях, включая конечные точки для почек. Информация о межбелковом взаимодействии и карты молекулярных путей были получены из общедоступных баз данных.
Молекулярные данные клинического фенотипа
Поиск литературы в NCBI Pubmed с использованием строки запроса диабетические нефропатии [majr] И (анализ микроматрицы [mh] ИЛИ профилирование экспрессии генов [mh]) И человек [mh] НЕ обзор привел к 37 исследованиям транскриптомики.Явно ограничиваясь исследовательскими исследованиями экспрессии мРНК на основе массивов в ткани почек человека, было получено четыре исследования, подходящих для включения в дальнейший анализ. Для Бертье и др. и Cohen et al. сигнатуры экспрессии могут быть получены непосредственно из публикаций (Cohen et al., 2008; Berthier et al., 2009). Для Woroniecka et al. и Baelde et al. необработанные профили экспрессии были получены из Gene Expression Omnibus (GSE30122, GSE1009) (Baelde et al., 2004; Woroniecka et al., 2011). Нормализация робастного усреднения по нескольким массивам (RMA) для набора данных Woroniecka et al.и нормализация MAS5 для набора данных Baelde et al., с последующим анализом значимости микрочипов (SAM) была использована для выявления признаков, демонстрирующих дифференциальную регуляцию, при сравнении диабетической болезни почек и здоровых контрольных образцов. В случае микродиссекции материала пробы проводили раздельный анализ клубочков и тубулоинтерстициального компартмента.
Для дальнейшего дополнения набора связанных с DN функций подход к анализу литературы, основанный на аннотации Pubmed Medical Subject Headings (MeSH) и публикация ссылок на гены, представленных в gene2pubmed (ftp: // ftp.ncbi.nlm.nih.gov/gene/DATA/gene2pubmed.gz) был выполнен. Поиск в Pubmed с использованием диабетических нефропатий [majr] И человеческих [mh] в качестве строки запроса был выполнен для идентификации публикаций, релевантных в контексте DN, в результате было получено 10 766 публикаций. Гены, кодирующие белки, подробно обсуждаемые в этих публикациях, были извлечены из gene2pubmed путем фильтрации на основе Pubmed ID и Taxonomy ID (9606 для человека).
Наконец, наборы дифференциально регулируемых признаков, идентифицированные в отдельных исследованиях транскриптомики, а также набор генов из литературных источников были объединены в пространстве имен генов Ensembl (Таблица 1).
Таблица 1. Пространство молекулярных данных диабетической нефропатии .
Биомаркер и аннотация из научной литературы
NCBI Pubmed поиск публикаций, содержащих диабетических нефропатий , далее квалифицируемых по одному из следующих квалификаторов патология, физиопатология, энзимология, метаболизм, осложнения, кровь, диагностика, моча и эпидемиология как основная концепция MeSH, требующая дальнейшего концепций MeSH Биологические маркеры или Онкологические маркеры, биологический выполняли для идентификации публикаций, в которых обсуждаются кандидаты в биомаркеры.Для поиска кандидатов в лекарственные мишени использовался термин Diabetic Nephropathies с классификаторами лекарственная терапия и терапия , соответственно. Поиск показал 615 публикаций по биомаркерам и 2692 публикации по лекарственным мишеням. Их соответствующие идентификаторы Pubmed впоследствии были использованы для извлечения генов человека из файла gene2pubmed, в результате чего было получено 54 кандидата в биомаркеры и 19 кандидатов-мишеней для лекарственных средств.
Аннотация цели через исследуемые препараты
Данные клинических испытаний для завершенных и текущих клинических испытаний были получены из ClinicalTrials.gov (http://clinicaltrials.gov/). Расширенный поиск, представленный на веб-странице ClinicalTrials.gov, использовался для определения исследований, которые удовлетворяли следующим двум критериям: Тип исследования равен Интервенционные исследования и Условие содержит диабетической нефропатии , в результате чего выявлено 206 клинических исследований. Заголовок и описание испытания были вручную пересмотрены, чтобы сосредоточить внимание на заболевании почек, в результате чего было рассмотрено 124 исследования. Соответствующие лекарственные вмешательства были сопоставлены с их записями в DrugBank (Law et al., 2014), извлекая перечисленные мишени для лекарственных препаратов человека, которые затем наносятся на карту в пространстве имен генов Ensembl. Всего с использованием этого подхода было идентифицировано 86 лекарственных мишеней, одна из которых также была частью 19 целевых кандидатов, извлеченных из научной литературы, в основном охватывающей фундаментальные и трансляционные исследования.
Механизм действия препарата Молекулярные данные
Набор ингибиторов АПФ был извлечен из системы классификации анатомо-терапевтических химических веществ (АТХ), поддерживаемой Всемирной организацией здравоохранения (ВОЗ).16 соединений, классифицированных как ингибиторы АПФ , простой (код ATC: C09AA), были идентифицированы и использованы для последующего извлечения данных из DrugMatrix (https://ntp.niehs.nih.gov/drugmatrix/index.html). Для шести из 16 препаратов в DrugMatrix были доступны наборы генов, на которые влияет присутствие препарата в ткани почек крысы после введения препарата. Полученные наборы генов крыс впоследствии были картированы из идентификаторов Unigene (Sayers et al., 2009) в идентификаторы крысы Ensembl и оттуда в гены ортологов человека согласно Ensembl (таблица 2).
Таблица 2. Механизм действия препарата Пространство данных .
Данные о молекулярном пути и взаимодействии белков
Информация о членстве в путях
KEGG и Panther для генов, кодирующих белок, была получена через службу REST KEGG и из файла базы данных в виде простого текста, доступного на веб-сайте Panther, соответственно (Thomas et al., 2003; Kanehisa et al., 2014). Данные человеческого белок-белкового взаимодействия из BioGRID, INTACT и Reactome были извлечены из соответствующих текстовых файлов, предоставленных отдельными источниками данных (Stark et al., 2006; Kerrien et al., 2012; Croft et al., 2014). Идентификаторы генов и белков, предоставленные в исходных источниках, были сопоставлены с их соответствующими идентификаторами генов Ensembl. Данные белок-белкового взаимодействия были дополнительно объединены в сеть белок-белкового взаимодействия с использованием идентификаторов генов Ensembl в качестве общего знаменателя для отдельных сетей.
Идентификация молекулярных путей и процессов
Молекулярные пути и процессы были проанализированы, с одной стороны, на основе обзора литературы о путях KEGG и Panther, которые уже обсуждались как релевантные в контексте DN.Во втором подходе de novo идентификация молекулярных процессов DN была выполнена с использованием набора функций патофизиологии DN. Алгоритм сегментации для идентификации процессов в сети взаимодействия белок-белок DN был разработан для сборки модели молекулярного процесса для DN. С использованием аналогичной процедуры была построена модель молекулярного механизма действия для ингибиторов АПФ с использованием сигнатур экспрессии, полученных из DrugMatrix.
DN пути из литературы
NCBI Pubmed поиск публикаций с использованием строки запроса «диабетическая нефропатия» [ti] ИЛИ «диабетическая нефропатия» [ti]) И (путь [ti] ИЛИ пути [ti]) был выполнен, в результате было найдено 53 публикации, содержащие ключевые слова в названии.Впоследствии было выполнено распознавание именованных сущностей для аннотирования появления названий путей в соответствии с записями KEGG и Panther в заголовке и аннотации этих публикаций. Наконец, отрывки, содержащие название пути, были проверены вручную, чтобы гарантировать ассоциацию идентифицированного пути в контексте DN, что привело к 27 отдельным путям, которые обсуждались в литературе как пораженные DN. Отношения между путями были выведены на основе общих генов и количества белок-белковых взаимодействий, охватывающих границы пути.
Молекулярные модели процессов
Расчет моделей молекулярных процессов выполнялся в соответствии с процедурой, описанной в Mayer et al. (2012); Heinzel et al. (2014). По сути, выполняются три основных этапа: (i) отображение сигнатуры признака, являющейся либо ассоциацией патофизиологии DN (таблица 1), либо набором механизмов действия ACE (таблица 2) в консолидированной сети взаимодействия белков с последующим индуцированным выделением подграфа. . Из подграфа удаляются узлы с нулевой степенью. (ii) идентификация молекулярных процессов с использованием алгоритма сегментации (MCODE с настройками по умолчанию, Bader and Hogue, 2003), и (iii) определение межпроцессных отношений, определяемых количеством белок-белковых взаимодействий, наблюдаемых между любыми фактическими двумя молекулярными процессами в сравнении против количества взаимодействий между двумя случайными наборами узлов с совпадающим размером набора узлов.
Анализ обогащения
Для определения значимости обогащения наборов молекулярных признаков в молекулярных процессах и путях использовался точный критерий Фишера с уровнем значимости 0,05. Поправка Бенджамини Хохберга была использована для корректировки множественного тестирования.
Результаты
DN Молекулярные пути
В результате скрининга научной литературы было обнаружено 27 молекулярных путей, наблюдаемых в контексте DN, в соответствии с аннотацией пути KEGG и Panther (рис. 1).На карте пути преобладают связанные компоненты передачи сигналов, при этом основными элементами являются взаимодействие MAPK-VEGF и Jak-STAT-цитокин-цитокиновый рецептор, дополнительно взаимодействующее с передачей сигналов TGF-бета, охватывая, среди прочего, механизмы реакции гипоксии и фиброза, соответственно (Rudnicki et al. др., 2009; Loeffler, Wolf, 2014). Дополнительные механистические аспекты включают реакцию на стресс и вовлечение внеклеточного матрикса (McLennan et al., 2013; Tan and de Haan, 2014). Кроме того, включен ряд специфических путей в контексте метаболизма, а также RAS, при этом последний, однако, не показывает прямых связей с другими путями на уровне перекрытия молекулярных признаков или на уровне прямого взаимодействия с белками.
Рис. 1. Пути развития диабетической нефропатии. Узлы на графике представляют пути KEGG и Panther (диаметр узла масштабируется с количеством назначенных генов, кодирующих белок), края между узлами масштабируются с числом перекрывающихся генов, а также взаимодействия генов через пути в соответствии с сетью взаимодействия белков. Пути отмечены для удержания кандидатов в биомаркеры (зеленый) и кандидатов-мишеней (красный).
В результате скрининга кандидатов в биомаркеры в научной литературе было выявлено 54 гена, кодирующих белок, из литературы были извлечены кандидаты в лекарственные мишени, а в результате клинических испытаний было выявлено 104 таких гена.Из 54 кандидатов-биомаркеров 23 назначены на карту путей DN, из 104 кандидатов-мишеней задействовано 52 (таблица 3).
Таблица 3. Аннотация молекулярных путей, диабетическая нефропатия .
Значительный охват в отношении биомаркеров, а также кандидатов-мишеней снова наблюдается для центральных компонентов передачи сигналов, включая передачу сигналов хемокинов, взаимодействие цитокин-цитокиновый рецептор, дополненное передачей сигналов MAPK и PI3K-Akt. Также рассматриваются механизмы, включая такие ключевые функции, как VEGFA и TGFB1.Для противодействия структурным изменениям внеклеточного матрикса не наблюдается специфического нацеливания, и, по-видимому, небольшие усилия прилагаются к адаптации стрессовой реакции. Для семи из 20 обсуждаемых путей биомаркеры или целевые аннотации не идентифицированы, и, кроме того, большое количество таких признаков присваивается также за пределами ландшафта пути, представленного на Рисунке 1. Яркие примеры для определения пустотных биомаркеров включают фактор роста соединительной ткани (CTGF) как Фактор фиброза, не назначенный в KEGG, то же самое верно и для уромодулина (UMOD), который, как было показано, связан с прогрессирующим заболеванием, включая генетические полиморфизмы (Deshmukh et al., 2013; Джеймс и др., 2013). CTGF также обсуждается в терапевтическом контексте с использованием подхода на основе моноклональных антител (Adler et al., 2010).
Тестирование набора функций патофизиологии DN, полученного из консолидации профилей транскриптомики в отношении обогащения данного ландшафта путей DN, выявило семь таких путей как значимые, однако без центральных механизмов, таких как реакция гипоксии или передача сигналов TGFB. Напротив, другие пути за пределами карты, представленной на рисунке 1, оказались значительно обогащенными, включая очаговую адгезию, молекулы клеточной адгезии и соединения адгезии, связанные с аспектами передачи сигналов, вовлеченными в заболевание.
DN Молекулярная модель
В дополнение к анализу молекулярных путей, как определено в KEGG и Panther, мы выполнили процедуру сегментации сети, направленную на идентификацию сегментов молекулярного процесса DN, определяемых топологическими характеристиками специфичного для DN подграфа. Из всего 881 гена, кодирующего белок, включенного в набор генов молекулярной патофизиологии DN (таблица 1), 880 также были частью консолидированной сети взаимодействия, а 634 были идентифицированы как члены индуцированного подграфа (рисунок 2A).Из общего набора из 880 признаков 246 генов, кодирующих белок, не взаимодействовали ни с одной другой функцией консенсусного набора DN, поэтому не учитывались при вычислении молекулярной модели. Очевидно, что элементы, извлеченные из литературы, относительно незначительно совпадают по сравнению с подписями из транскриптомики. Из общего количества 516 уникальных особенностей, объединенных в результате четырех экспериментов по профилированию транскриптомики, и 414 особенностей, взятых из научной литературы, 49 являются общими.
Рисунок 2.Молекулярная модель диабетической нефропатии. (A) Индуцированный подграф, где каждый узел представляет ген, кодирующий белок, который сообщается как связанный с DN, края обозначают взаимодействия в соответствии с базовой сетью взаимодействий. Элементы, полученные в результате исследований Omics, выделены красным цветом, элементы, выделенные из литературных источников, — зеленым, элементы, идентифицированные в обоих источниках данных, показаны синим. (B) Молекулярная модель, представляющая DN, где каждый узел представляет собой сегмент процесса с масштабированием диаметра узла с количеством задействованных генов, кодирующих белок, и границами между узлами, масштабируемыми с количеством взаимодействий генов между узлами в соответствии с взаимодействием белков сеть.Сегменты указаны для содержания кандидатов в биомаркеры (зеленый) и кандидатов-мишеней в лекарственные средства (красный).
После сегментации MCODE 200 молекулярных особенностей остались в сегментах процесса, образуя молекулярную модель, содержащую 23 сегмента процесса (рис. 2B). Среднее количество генов, кодирующих белок на сегмент процесса, составляет 6, причем самый большой сегмент кодирует 29 признаков, наименьший 3. Эквивалентно графику путей на Рисунке 1, график процесса служит аппроксимацией индивидуальных характеристик молекулярного процесса вместе с их зависимостями.Шесть сегментов процесса модели процесса содержат аннотацию как биомаркера, так и целевого кандидата, а другие кодируют только один из двух или ни одного. Из 54 кандидатов-биомаркеров 22 включены в молекулярную модель, соответствующее число для 104 кандидатов-мишеней составляет 16.
DN Молекулярная модель и механизм действия лекарственного средства Вмешательство модели
Объединение сигнатур транскриптомики, отражающих влияние ингибиторов АПФ на почечный интерактом в модели на крысах с использованием шести репрезентативных препаратов, привело к 2058 молекулярным характеристикам (таблица 2), при этом 661 признак был идентифицирован как минимум в двух из шести сигнатур лекарств.Сопоставление этого согласованного подмножества функций ACE в консолидированной сети взаимодействия позволило представить 656 функций. Индуцированный подграф включал 332 признака, после сегментации, в результате которой получилось 12 сегментов процесса, содержащих в общей сложности 92 молекулярных признака (Рисунок 3, слева). Средний размер набора функций процесса составлял 8, максимум 19 и минимум 3.
Рис. 3. Молекулярная модель механизма действия ингибитора АПФ и взаимодействие с молекулярной моделью DN. Молекулярная модель механизма действия ACE (слева) и молекулярная модель DN (справа), с перекрывающимися сегментами процесса моделей лекарственного средства и фенотипа, обозначенными пунктирными линиями.Сегменты молекулярного процесса (U) молекулярной модели механизма действия ACE, показывающие вмешательство в молекулярную модель DN, выделены синим цветом, соответствующие сегменты взаимодействующего процесса на стороне DN показаны красным.
Вмешательство молекулярной модели механизма действия ACE с молекулярной моделью DN на уровне перекрытия признаков (рис. 3) позволило идентифицировать определенные сегменты процесса молекулярной модели DN, также содержащие кандидаты в биомаркеры (таблица 4).
Таблица 4.Вмешательство сегмента диабетической нефропатии .
Все четыре сегмента процесса DN, показывающие вмешательство в модель механизма действия препарата ACE, содержат кандидатов в биомаркеры. Два сегмента обеспечивают значительное обогащение также на уровне молекулярных путей, показывая интеграцию передачи сигналов хемокинов и цитокинов, RAS и каскадов комплемента и коагуляции для одного сегмента процесса, второй сегмент процесса отражает компоненты передачи сигналов PI3K-Akt в контексте передачи сигналов TGFB и взаимодействие рецепторов ЕСМ.
Кандидаты в биомаркеры, выступающие в качестве заместителя для интерференции молекулярных моделей ACE и DN, включают лиганд 5 хемокинов (мотив CC), участвующий в иммунорегуляторах и воспалительных процессах, гемоглобин альфа 1 и 2 вместе с гаптоглобином, цитокин-трансформирующим фактором роста, бета-1, а также с фактором транскрипции NFKB1, включая, наконец, актин альфа-1, участвующий в подвижности, структуре и целостности клеток.
Обсуждение
В ходе трансляционных и доклинических исследований для широкого спектра клинических представлений было предложено впечатляющее количество мишеней для лекарственных средств, при этом значительное количество препаратов было продолжено в клинических испытаниях.Только в первой половине 2014 года на платформе Clinicaltrials.gov было зарегистрировано около 10 000 новых клинических исследований. Если рассматривать диабетическую нефропатию как клинический фенотип, то на сайте Clinicaltrials.gov идентифицировано 124 интервенционных испытания в любом статусе, конкретно включающих термин заболевания, охватывающих 45 отдельных лекарственных препаратов, направленных на 86 известных целей. Из научной литературы выявлено еще 18 мишеней для лекарственных препаратов.
Наряду с рядом испытаний с использованием лекарств и комбинаций лекарств, направленных на изучение известных факторов, влияющих на прогрессирование DN, таких как RAS, мишени лекарств неравномерно распределены по молекулярным путям, следовательно, механизмы, приписываемые болезни.
Из литературных источников 27 различных путей в соответствии с аннотациями путей KEGG и Panther обсуждаются как связанные с DN, из которых 19 содержат мишени для лекарств. К ним относятся хорошо известные механизмы, имеющие отношение к DN, включая реакцию гипоксии или фиброз, в сочетании с большим набором сигнальных компонентов. Кроме того, 52 лекарственные мишени встроены в молекулярный контекст за пределами описанного в литературе ландшафта путей ДНК.
Аналогичная ситуация для кандидатов в биомаркеры.54 уникальных белка, извлеченных из научной литературы, обсуждаются в контексте любых биомаркеров, охватывающих 14 из 27 путей, при этом 31 кандидат в биомаркеры не назначен ни одному из членов выделенной карты путей ДНК.
Интересно, что прогностическая эффективность в отношении прогрессирования заболевания любого из индивидуальных биомаркеров-кандидатов оказалась ограниченной. Например, в обзоре Hellemons et al. 13 релевантных маркеров были обнаружены в контексте нефропатии при диабете, пять из которых были обнаружены как значимо связанные с началом, а также с прогрессированием DN, снова охватывая различные механизмы, включая воспаление (например,g., C-реактивный белок), взаимодействие и гомеостаз клеточной поверхности (например, E-селектин, ICAM1) и метаболизм (уровни триглицеридов) (Hellemons et al., 2012).
По-видимому, отдельные биомаркеры, отражающие статус отдельного молекулярного процесса, пути или механизма, не могут фиксировать прогноз болезни для полной популяции DN. В альтернативных подходах панели мультимаркеров были включены в классификаторы по диагностике и прогнозу заболеваний, демонстрируя улучшение показателей также при слепой проверке.В Roscioni et al. сигнатура из 273 пептидов, определенная в моче, была включена в машинный классификатор опорных векторов (Roscioni et al., 2013). Сигнатура содержала фрагменты коллагена, в конечном итоге отражающие изменения в обмене внеклеточного матрикса и фиброзе вместе с маркерами воспаления, такими как, например, провоспалительный белок S100-A9, а также уромодулин, который, как было показано, связан с интерстициальным фиброзом и атрофией канальцев (Nkuipou- Kenfack et al., 2014).
Одним из факторов, влияющих на потребность в панелях мультимаркеров, может быть индивидуальная дисперсия базовых уровней биомаркеров, когда включение нескольких маркеров специально в методы нелинейной классификации повышает надежность.Однако вторым фактором может быть общая неоднородность популяции пациентов. Конкретные проявления заболевания могут значительно различаться не только на разных стадиях прогрессирования заболевания, в конечном итоге наблюдая переход от защитных к повреждающим механизмам, но даже в рамках конкретной категории хронических заболеваний почек, как это определено в настоящей клинической классификации, представленной в рекомендациях KDIGO (члены Совета KDIGO, 2013).
Улучшенные прогностические характеристики панелей мультимаркеров в дополнение к строгой функциональной классификации переходов стадий в DN с использованием альбуминурии, а также рСКФ в качестве клинически используемых параметров прогрессирования явно подтверждают случай патофизиологической гетерогенности, в нынешних клинических условиях однородной популяции пациентов.Однако конкретно для альбуминурии обсуждается роль функционального маркера по сравнению с фактором заболевания (Roscioni et al., 2014).
Получение надежных диагностических или прогностических классификаторов, например, на основе профилей протеомики или метаболомики, может улучшить клиническое ведение пациента в отношении начала, а также интенсивности терапевтических мер (Roscioni et al., 2013; Pena et al., 2014). Также в дизайне клинических испытаний такие стратегии обогащения можно использовать, например, путем выявления лиц, склонных к быстрому прогрессированию заболевания, и рандомизации в этой когорте высокого риска в группу лечения и плацебо (например,g., Priority study, ссылка на Clinicaltrials.gov NCT02040441).
Прогностические биомаркеры, в отличие от диагностических параметров с известным назначением молекулярных процессов и путей, дополнительно позволяют приблизительно определить, какие конкретные механизмы связаны с прогрессированием заболевания. Пейзаж путей DN, обсуждаемый в этой работе, является исключительно поперечным представлением болезни, в первую очередь не позволяя расшифровать, какой из 27 отдельных путей ведет к прогрессированию болезни, а какие другие пути являются просто сторонними наблюдателями или последующими последствиями механистических факторов болезнь.Следовательно, оценка кандидатов в биомаркеры на предмет их связи с прогрессирующим заболеванием, в свою очередь, позволяет определить механизмы, связанные с прогрессирующим заболеванием. Такие знания жизненно важны, например, для определения новых лекарственных средств, требующих включения в механизмы заболевания, являющиеся факторами прогрессирования заболевания. Однако остается вопрос, актуальны ли такие механизмы в той же степени или вообще для конкретного пациента, отнесенного к клиническому фенотипу.
Набор прогностических биомаркеров, охватывающий все потенциально значимые процессы, позволяет проводить специфическое молекулярное фенотипирование отдельных пациентов, однако этого недостаточно с точки зрения прогнозирования ответа на лекарственный препарат, поскольку механизм действия лекарственного средства не учитывается.Здесь системная фармакология направлена на идентификацию лекарственного ответа также на уровне молекулярных процессов и путей. Обоснование состоит в том, чтобы не только сосредоточиться на конкретной мишени лекарственного средства и ее назначении конкретным механизмам, но и включить системные молекулярные изменения, запускаемые лекарством, включая эффекты вне мишени, а также последующие молекулярные изменения. Наличие механизма действия лекарства, а также клинического фенотипа, представленного на уровне молекулярного процесса или пути, позволяет пересекать оба молекулярных состояния.Если из прогностического профиля биомаркера пациента идентифицированы специфические молекулярные механизмы заболевания, связанные с прогрессированием, и лекарство проявляет функциональное вмешательство в такие специфические механизмы, такой пациент может быть более склонным к проявлению ответа на лекарство. С такой настройкой, включая знания о молекулярном фенотипическом составе, значимости молекулярных процессов в прогрессирующем заболевании и знаниях о влиянии лекарственного механизма действия-кандидаты в биомаркеры, первоначально служащие прогностической цели, можно превратить в прогностические биомаркеры на лекарственный ответ.
Профилирование Omics вносит большой вклад в характеристику как клинических фенотипов, так и механизма действия лекарства. Интеграция результатов профилирования клинических выборок часто приводит к незначительному совпадению отдельных исследований, отчасти из-за недостаточного размера выборки в сочетании с разными критериями включения и использованным материалом выборки (Fechete et al., 2011). В представленном здесь примере 1010 признаков в общей сложности идентифицированы как дифференциально регулируемые в транскриптомике или присваиваются DN согласно литературному анализу, с 880 уникальными признаками.Эквивалентный дисбаланс в согласованности характеристик в исследованиях также обнаружен для данных транскриптомики ингибиторов АПФ. Все эти препараты относятся к одному и тому же функциональному контексту, но из всего 3152 признаков, идентифицированных для шести включенных препаратов, общее количество уникальных признаков все еще составляет 2058, причем 661 идентифицируется как минимум в двух сигнатурах лекарств.
Следующее расхождение становится очевидным — ограниченное совпадение анализа обогащения, основанного на сигнатурах профилирования и анализа литературы на основе признаков, по сравнению с явным анализом литературы для молекулярных путей.Из 27 путей, извлеченных из научных источников, подтверждены только семь, однако другие пути обогащения не были обнаружены с помощью литературных источников. Кроме того, основным недостатком является ограниченное представление генов, кодирующих белок, на таких картах путей, например, для генов, кодирующих белок 6491, и для генов, кодирующих белок Panther 2163, соответственно. Это ограничение влияет не только на обогащение пути, но также на назначение биомаркера и целевых кандидатов. Из общего количества 104 лекарственных мишеней и 54 кандидатов в биомаркеры 29 не относятся ни к какому пути KEGG или Panther.
Здесь можно использовать другой подход, а именно сегментацию сетей взаимодействия белков, демонстрирующую улучшенный охват набора генов, кодирующих белок. Объединение INTACT, Reactome и BioGRID позволяет представить в общей сложности 13 907 генов, кодирующих белки, что явно выходит за рамки общедоступных баз данных о путях. В альтернативных подходах гибридные сети взаимодействия используются для дальнейшего расширения охвата генов, кодирующих белок, а также для улучшения ложноотрицательных показателей в отношении белок-белковых взаимодействий и отношений (Fechete et al., 2013).
Вычисление подграфа, специфичного для DN, а также специфичного для ингибитора АПФ с последующей сегментацией на основе топологии позволяет альтернативное представление ландшафта молекулярных процессов для клинической картины, а также механизма действия лекарства. Анализ интерференции на уровне перекрывающихся генов, кодирующих белок, привел к четырем сегментам процесса, содержащим центральные аспекты патофизиологии DN. В этих мешающих молекулярных процессах было идентифицировано семь кандидатов в биомаркеры.Было показано, что CCL5 (RANTES), участвующий в рекрутировании моноцитов и макрофагов в почечную кору, подавляется ингибированием АПФ, указывая на то, что экспрессия RANTES опосредуется через рецептор ангиотензина II типа 2 (Kashiwagi et al., 2002). Аналогичным образом было показано, что на животных моделях экспрессия TGFB1 снижается ингибиторами АПФ. Активация NFKB1 ангиотензином II была показана в гладких мышцах сосудов и мезангиальных клетках (Hernández-Presa et al., 1997). В исследовании Dong et al. Анализ экономической эффективности лечения ингибиторами АПФ пациентов с сахарным диабетом 1 типа показал, что уровень гликозилированного HbA1c явно влияет на экономическую эффективность использования лекарств в расчете на год жизни с поправкой на качество (QALY) (Dong et al., 2004). Авторы пришли к выводу, что, помимо возраста пациента, при рассмотрении терапии необходимо учитывать и другие факторы.
Очевидно, механизм действия лекарства влияет на многочисленные молекулярные процессы, например, ингибиторы АПФ, многие из которых также влияют на прогрессирование DN. Анализ взаимодействия молекулярных процессов патофизиологии, связанной с прогрессированием заболевания, и механизма действия лекарств позволяет предложить прогностические маркеры. Тестирование таких кандидатов в прогностические биомаркеры может дать представление о значимости отдельных процессов на уровне пациента, что напрямую связано с вероятностью реакции на лекарство.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Исследование, приведшее к этим результатам, получило финансирование от Седьмой рамочной программы Европейского сообщества в соответствии с соглашением о гранте No. 241544 (SysKid).
Список литературы
Адлер, А.И., Стивенс, Р.Дж., Мэнли, С. Е., Билоус, Р. У., Калл, К. А., и Холман, Р. Р. (2003). Развитие и прогрессирование нефропатии при диабете 2 типа: проспективное исследование диабета Соединенного Королевства (UKPDS 64). Почки Инт . 63, 225–232. DOI: 10.1046 / j.1523-1755.2003.00712.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Адлер С.Г., Шварц С., Уильямс М.Э., Арауз-Пачеко К., Болтон В.К., Ли Т. и др. (2010). Исследование фазы 1 моноклональных антител против CTGF у пациентов с диабетом и микроальбуминурией. Clin. Варенье. Soc. Нефрол . 5, 1420–1428. DOI: 10.2215 / CJN.09321209
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Баелде, Х. Дж., Эйкманс, М., Доран, П. П., Лаппин, Д. У. П., де Хеер, Э. и Брейн, Дж. А. (2004). Профили экспрессии генов в клубочках почек человека с диабетической нефропатией. г. Дж. Почки Дис . 43, 636–650. DOI: 10.1053 / j.ajkd.2003.12.028
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Бертье, К.К., Чжан, Х., Шин, М., Хенгер, А., Нельсон, Р. Г., Йи, Б. и др. (2009). Повышенная экспрессия сигнального преобразователя киназы Janus и активатора участников пути транскрипции при диабетической нефропатии человека. Диабет 58, 469–477. DOI: 10.2337 / db08-1328
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Бреннер, Б. М., Купер, М. Э., де Зеу, Д., Кин, В. Ф., Митч, В. Е., Парвинг, Х. Х., и др. (2001). Влияние лозартана на почечные и сердечно-сосудистые исходы у пациентов с диабетом 2 типа и нефропатией. N. Engl. J. Med . 345, 861–869. DOI: 10.1056 / NEJMoa011161
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Buyse, M., Loi, S., van’t Veer, L., Viale, G., Delorenzi, M., Glas, A.M, et al. (2006). Подтверждение и клиническая полезность прогностической сигнатуры из 70 генов для женщин с раком молочной железы без лимфоузлов. J. Natl. Институт рака . 98, 1183–1192. DOI: 10.1093 / jnci / djj329
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Коэн, К.Д., Линденмейер, М. Т., Эйхингер, Ф., Хан, А., Зейферт, М., Молл, А. Г. и др. (2008). Улучшенное выяснение биологических процессов, связанных с диабетической нефропатией, с помощью анализа данных микроматрицы на основе одного зонда. PLoS ONE 3: e2937. DOI: 10.1371 / journal.pone.0002937
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Крофт Д., Мундо А. Ф., Хоу Р., Милачич М., Вейзер Дж., Ву Г. и др. (2014). База знаний Reactome pathway. Nucleic Acids Res .42, D472 – D477. DOI: 10.1093 / nar / gkt1102
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Дешмук, Х. А., Палмер, К. Н. А., Моррис, А. Д., Колхун, Х. М. (2013). Исследование известных локусов скорости клубочковой фильтрации у пациентов с диабетом 2 типа. Диабет. Мед . 30, 1230–1235. DOI: 10.1111 / dme.12211
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
De Zeeuw, D., Akizawa, T., Audhya, P., Bakris, G.L., Chin, M., Christ-Schmidt, H., et al. (2013). Бардоксолон метил при диабете 2 типа и хронической болезни почек 4 стадии. N. Engl. J. Med . 369, 2492–2503. DOI: 10.1056 / NEJMoa1306033
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Донг, Ф. Б., Соренсен, С. В., Маннинен, Д. Л., Томпсон, Т. Дж., Нараян, В., Орианс, К. Э. и др. (2004). Экономическая эффективность лечения пациентов с сахарным диабетом 1 типа ингибиторами АПФ. Фармакоэкономика 22, 1015–1027.DOI: 10.2165 / 00019053-200422150-00005
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Fechete, R., Heinzel, A., Perco, P., Mönks, K., Söllner, J., Stelzer, G., et al. (2011). Картирование молекулярных путей, биомаркеров и мишеней лекарств при диабетической нефропатии. Proteomics Clin. Заявление . 5, 354–366. DOI: 10.1002 / prca.201000136
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Fechete, R., Heinzel, A., Soellner, J., Perco, P., Лукас, А., Майер, Б. (2013). Использование информационного содержания для расширения сетей взаимодействия генов, кодирующих человеческий белок. J. Comput. Sci. Syst. Биол . 6, 73–82. DOI: 10.4172 / jcsb.1000102
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Фрид, Л. Ф., Эмануэле, Н., Чжан, Дж. Х., Брофи, М., Коннер, Т. А., Дакворт, В. и др. (2013). Комбинированное ингибирование ангиотензина для лечения диабетической нефропатии. N. Engl. J. Med . 369, 1892–1903.DOI: 10.1056 / NEJMoa1303154
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Хайнцель А., Фешете Р., Мюльбергер И., Перко П., Майер Б. и Лукас А. (2013). Молекулярные модели кардиоренального синдрома. Электрофорез 34, 1649–1656. DOI: 10.1002 / elps.201200642
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Heinzel, A., Fechete, R., Söllner, J., Perco, P., Heinze, G., Oberbauer, R., et al. (2012). Графики данных для связи клинического фенотипа и пространства молекулярных признаков. Внутр. J. Syst. Биол. Биомед. Технол . 1, 11–25. DOI: 10.4018 / ijsbbt.2012010102
CrossRef Полный текст
Хайнцель А., Мюльбергер И., Фешете Р., Майер Б. и Перко П. (2014). Функциональные молекулярные единицы для управления дизайном панели биомаркеров. Methods Mol. Биол . 1159, 109–133. DOI: 10.1007 / 978-1-4939-0709-0_7
Физиология сенсорных систем
ВОПРОСЫ ДЛЯ ЭКЗАМЕНА ПО КУРСУ НОРМАЛЬНОЙ ФИЗИОЛОГИИ
1 семестр
Объект и цель физиологии.Методы исследования в физиологии. История физиологии. Развитие физиологии в России: Сеченов, Павлов, Введенский, Ухтомский. Казанская физиологическая школа: Овсянников, Ковалевский, Миславский, Самойлов. Современный этап развития физиологии.
Регуляция физиологических функций. Понятие гомеостаза. Система регулирования. Нервная и гуморальная регуляция. Положительные и отрицательные отзывы.
Физиология возбудимых систем
Строение клеточной мембраны возбудимых клеток, основные функции.Ионные каналы, классификация и физиологическая роль. Механизмы активации ионных каналов (лиганд-зависимый, потенциал-зависимый, механочувствительный).
Характеристика внутри- и внеклеточной жидкости возбудимых клеток. Транспорт через клеточную мембрану. Ионные насосы, их разновидности. В первую очередь и во вторую очередь активный транспорт. Ионотропные и метаботропные рецепторы клеточной мембраны.
Возбудимость. История открытия электрических явлений. Мембранный потенциал.Факторы, обеспечивающие его внешний вид и содержание. Параметры возбудимости. Пороговое раздражение, хронаксия, лабильность. Потенциал действия, ионные механизмы его возникновения. Анализ фаз потенциала действия. Регенеративная деполяризация. Огнеупор, его фаза. Механизмы и физиологическое значение инактивации натрия. Закон «все или ничего».
Местный отклик, механизм возникновения. Сравнение местного отклика со свойствами потенциала действия.Действие ДК на возбудимые мембраны. Закон полярного раздражения (Закон Пфлюгера). Феномен аккомодации.
Особенности возбуждения миелинизированных и немиелинизированных нервных волокон. Скорость проведения импульса. Классификация нервных волокон по скорости проведения. Законы проведения возбуждения по нервным волокнам.
Химические и электрические синапсы. Проведение возбуждения в нервно-мышечном соединении. Строение синапса. Квантовая секреция нейромедиатора.Спонтанная квантовая секреция. Экзоцитоз синаптических везикул. Активация никотинового ацетилхолинового рецептора постсинаптической мембраны. Роль ацетилхолинэстеразы. Потенциалы концевой пластинки и генерация потенциала действия в мышечном волокне. Пре- и постсинаптические механизмы действия физиологически активных веществ и фармацевтических препаратов на нервно-мышечную передачу. Нарушение нервно-мышечной передачи при переутомлении.
Типы мышечных волокон. Строение миофибрилл как функциональной единицы мышечного волокна.Механизм сокращения мышц в саркомере поперечно-полосатых мышц. Теория скользящей нити. Роль ионов кальция и АТФ в сокращении мышц. Электромеханическая муфта. Процесс расслабления мышц. Трупное окоченение. Простое подергивание, суммирование и столбняк. Типы тетанических сокращений. Тоническое сокращение мышц. Контрактура. Энергия сокращения мышц. Фазические и тонические мышечные волокна. Особенности функционирования и метаболизма окислительных и гликолитических волокон.
Концепция моторного агрегата.Регулировка мышечной силы. Развитие утомления тела, нервно-мышечной подготовки и отдельных мышц.
Особенности строения, иннервации и сокращения гладких мышц. Самопроизвольные сокращения гладкой мускулатуры и их механизм. Единичная (унитарная) и многоэлементная гладкая мышца
Центральная нервная система
Рефлекс как общий принцип регуляции функций организма. Рефлекторная дуга и ее составляющие.Классификация рефлексов. Соматические и вегетативные рефлексы. Моно- и полисинаптические рефлексы. Основные типы и функции нейрональных и глиальных клеток. Гематоэнцефалический барьер, механизмы и функции. Центральные синапсы. Медиаторы в ЦНС. EPSP и IPSP. Пре- и постсинаптическое торможение. Явление суммирования — временное и пространственное суммирование.
Свойства и характеристики возбуждения нейронных сетей: дивергенция, конвергенция, потенциация, окклюзия.Принцип конвергенции (конечный общий путь). Торможение в ЦНС (И. М. Сеченов), его виды и роль. Современное понимание механизмов центрального торможения. Взаимное, латеральное торможение Реншоу. Понятие о нервном центре, его свойствах. Доминирующий принцип. Особенности доминирующего центра (Ухтомский). Факторы, способствующие появлению доминанты.
Спинной мозг. Двигательная функция спинного мозга. Моносинаптические и полисинаптические рефлексы. Афферентная проприоцептивная система.Экстрафузальные и интрафузальные волокна. Понятие о гамма-петле. Регулировка мышечного тонуса. Сухожильные рефлексы. Соматические полисинаптические рефлексы (сгибатель, выпрямитель). Автоматические программы спинного мозга. Спинальные рефлексы вызывают мышечный спазм. Спинальные вегетативные рефлексы. Передача информации по спинному мозгу. Спинальный шок.
Двигательная функция ствола головного мозга. Статические и статокинетические рефлексы. Моторные ядра ствола головного мозга: красные, вестибулярные, ядра ретикулярной формации. Функции красных ядер, их влияние на альфа и гамма мотонейроны спинного мозга.Децеребрационная жесткость. Постуральные рефлексы продолговатого мозга. Важнейшие вегетативные рефлексы продолговатого мозга. Роль среднего мозга в реализации статокинетических рефлексов. Зрительные и слуховые ориентировочные рефлексы. Спастичность и жесткость.
Мозжечок и его основные функции. Связь мозжечка с другими отделами центральной нервной системы ниже и выше по течению. Последствия удаления мозжечка. Симптомы, возникшие в результате отказа мозжечка, и их причины.
Ретикулярная формация, ее структура и функции нервной организации. Ретикулярные нейроны. Полисенсорные ретикулярные нейроны. Ретикулярная активирующая система.
Промежуточный мозг. Таламус как сборщик конфиденциальной информации. Ядра таламуса. Гипоталамус и его функции. Характеристики нейронов и ядер гипоталамуса.
Базальные ядра. Значение базальных ганглиев в координации двигательной активности как посредника между ассоциативной и моторной областями коры.Значение «черного вещества», его связь с базальными ядрами. Болезнь Паркинсона.
Двигательные области коры головного мозга. Роль пирамидальной и экстрапирамидной систем в организации двигательных актов.
Лимбическая система, ее строение, основная физиологическая функция.
Кора больших полушарий головного мозга, ее строение. Клеточная структура коры. Представление функций в коре. Сенсорная, двигательная и ассоциативная области.Локализация функций в коре головного мозга.
Симпатический и парасимпатический отделы вегетативной нервной системы, их характеристика. Медиаторы симпатической и парасимпатической систем. Механизм действия медиаторов симпатического и парасимпатического отделов на различные рецепторы. Вегетативные рефлексы и центры регуляции вегетативных функций.
Электрические явления в коре головного мозга в состоянии покоя и деятельности организма. Электроэнцефалография.Основные виды электрической активности на ЭЭГ и механизмы их возникновения. Вызванные потенциалы. Клиническое использование ЭЭГ.
Электромиография. Диагностика мышечного аппарата с помощью электромиографии.
Физиология сенсорных систем
Анализаторы. Строение и физиологическое значение. Кодирование информации в сенсорных системах. Понятие ощущения. Общие свойства рецепторов. Классификация рецепторов и их типов.Классификация рецепторов. Из первичных и вторично-сенсорных рецепторных клеток. Механизм возбуждения в рецепторах. Рецепторный потенциал. Адаптация рецепторов. Особенности кодирования в анализаторах. Кодирование поступающей информации, примеры некоторых видов кодирования.
Визуальный анализатор. Оптическая система глаза. Зрачок и зрачковый рефлекс. Размещение глаза. Нарушения рефракции глаза (близорукость, дальнозоркость, астигматизм). Пресбиопия. Строение сетчатки глаза.Фоторецепторы. Слепая зона. Фотохимические реакции в рецепторах сетчатки. Электрические явления в сетчатке и зрительном нерве. Рецептивные поля нейронов. Электроретинограмма. Электрическая активность проводящих путей и центров зрительного анализатора. Темная и светлая адаптация. Цветовое зрение. Теория цветового восприятия. Дальтоник. Двигательный аппарат глаза. Временные характеристики движений глаз (саккады, периоды фиксации и плавность движений). Обработка зрительной информации в таламусе.Преобразование зрительной сенсорной стимуляции в таламус. Анализ зрительной сенсорной стимуляции нейронов зрительной коры.
Анализатор слуха. Строение и функции наружного, среднего и внутреннего уха. Кортиев орган, его строение и механизм возбуждения. Восприятие звуков разной частоты. Механизм генеза рецепторного потенциала волосковых клеток спирального ганглия. Проведение и анализ звука в ЦНС.
Вестибулярный анализатор.Родные раздражители для отолитового аппарата и полукружных каналов. Центральная часть вестибулярного аппарата, поддержание равновесия. Вестибулярные рефлексы, нистагм.
Анализаторы запаха и вкуса. Расположение и строение. Порог чувствительности. Приспособление. Чувствительность рецепторов к разным вкусовым раздражителям. Карта вкуса языка. Централизованная обработка вкусовой и обонятельной информации
Сомато-висцеральная чувствительность. Модальные свойства.Классификация афферентных нервных волокон, рецепторных структур, типов чувствительности. Кожные механорецепторы, их классификация, гистология, функциональное значение. Афферентная иннервация кожи, рецепторного поля.
Проприоцепция и ее качество. Классификация проприорецепторов, гистология, иннервация. Центральная интеграция проприоцептивных стимулов. схема тела.
Терморецепция. Статические и динамические чувства. Ощущение жара и холода.
Ноцицепция.Качества боли. Болезненные раздражители, адаптация к боли. Нейрофизиологические основы боли: теория восприятия боли. Ноцицепторы и их иннервация. Медиаторы боли и антиноцицептивная система.
Обработка сенсорной информации в ЦНС. Уровни обработки сенсорной информации, специфическая и неспецифическая сенсорная система. Афферентные связи в спинном мозге. Соматосенсорная функция ствола головного мозга, роль ретикулярной формации. Специфические и неспецифические ядра таламуса.Соматосенсорные области коры и их проекция тела.
: 2017-01-28; : 500 | |
:
:
:
© 2015-2020 lektsii.org — —
Вступительная глава: Механизмы действия антибиотиков и устойчивость к антибиотикам
2.4.1.1 Репликация ДНК
Предшественники дезоксирибонуклеотидов синтезируются для полимеризации дезоксинуклеотидов.Под действием киназного фермента дезоксинуклеозидтрифосфаты (dNTP: dGTP, dCTP и dATP) синтезируются из дезоксинуклеозиддифосфатов (dNDP: dGDP, dCDP и dADP), которые синтезируются из рибонуклеозидов с помощью рибонуклеотидредуктазы. Но дезокситимидинтрифосфат (дТТФ) синтезируется другим путем. Дезоксиурацилдифосфат (dUDP), который синтезируется из урацилдифосфата (UDP) рибонуклеотидредуктазой, превращается в дезоксиурацилмонофосфат (dUMP). Тимидилатсинтетаза катализирует реакцию, которая превращает dUMP, метилированный тетрагидрофолатом (THF), в dTMP.На этой стадии тетрагидрофолат (THF) синтезируется из дигидрофолата (DHF) дигидрофолатредуктазой. Наконец, dTTP синтезируется киназой из dTMP (рис. 2).
ДНК-гираза открывает цепи ДНК для полимеризации дезоксинуклеотидов ДНК-полимеразой в соответствии с каждой кольцевой матричной цепью хромосомы [5].
2.4.1.2 Подавление репликации и механизмы устойчивости
Кинолоны, такие как налидиксовая кислота и ципрофлоксацин, которые используются для лечения инфекций, вызванных Pseudomonas spp., предотвращают образование репликационной вилки путем ингибирования ДНК-гиразы в результате связывания с субъединицей A gyr. Новобиоцин и кумермицин предотвращают образование репликационной вилки, ингибируя ДНК-гиразу в результате связывания с субъединицей gyr B. Когда гены бактерий gyr A и gyr B мутируют, у бактерий развивается устойчивость к этим антибиотикам [5]. Норфлоксацин, гатифлоксацин, гемифлоксацин и моксифлоксацин — это другие группы флорхинолонов, обладающие более широким спектром действия.Флорхинолоны нетоксичны. Ципрофлоксацин, норфлоксацин, гатифлоксацин и гемифлоксацин используются для лечения инфекции мочевыводящих путей и пневмонии [2].
Синтез предшественников дезоксинуклеотидов, которые используются для репликации ДНК, можно блокировать триметопримом, гидроксимочевиной, 5-фтордезоксиуридином и 5-фторурацилом.
Триметоприм — ингибитор синтеза фолиевой кислоты. Триметоприм, который является структурным аналогом DHF, предотвращает синтез THF, ингибируя дигидрофолатредуктазу.Итак, dTMP, являющийся предшественником дезоксинуклеотидного полимера, не синтезируется. Существует множество механизмов устойчивости к триметоприму. Если ThyA, то в гене микроорганизма происходит мутация, инактивирующая тимидилат, сентетаз dTMP и DHF не синтезируются, в результате невозможности переноса метильной группы от THF к dUMP. Поскольку дигидрофолатредуктаза не синтезируется в микроорганизме, у которого мутирован ген thyA , этот микроорганизм не ингибируется триметопримом.Если микроорганизм изменяет сайт связывания дигидрофолатредуктазы, триметоприм не может связываться с дигидрофолатредуктазой. Итак, бактерии вырабатывают устойчивость к триметоприму. Другой механизм устойчивости к триметоприму — перенос гена, вызывающего устойчивость к триметоприму [5].
Гидроксимочевина ингибирует рибонуклеотидредуктазу, которая катализирует дезоксинуклеозиддифосфат из рибонуклеозиддифосфата (рис. 2). При мутации микроорганизма гена, кодирующего рибонуклеотидредуктазу, у микроорганизма развивается устойчивость к гидроксимочевине [5].
Синтез предшественника может блокироваться 5-фтордезоксиуридином и 5-фторурацилом с конкурентным ингибированием. Монофосфатные формы 5-фтордезоксиуридина и 5-фторурацила являются структурными аналогами dUMP, который является субстратом тимидилатсинтазы. Они подавляют синтез dTMP. В случае мутации микроорганизма гена, кодирующего тимидилатсинтазу, у микроорганизма развивается устойчивость к 5-фтордезоксиуридину и 5-фторурацилу [5].