красота мира в каждом кадре
Не стоит рассматривать простые зрительные органы лишь как одну из промежуточных ступеней эволюции на пути к сложным устройствам: все они исправно служили своим обладателям. У морских звезд глаза, расположенные на кончиках лучей, не различают цвет и мелкие детали и не могут заметить быстро движущиеся объекты. С таким зрением летящий орел врезался бы в первое попавшееся дерево. Но ведь морские звезды не выслеживают бегущего по полю зайца с высоты небоскреба, им и нужно-то всего ничего: разобраться, где находится коралловый риф — огромное незыблемое подводное сооружение, — чтобы неторопливо добраться до дома. Для этого звездам достаточно тех глаз, какие есть, и ничего лучшего им не надо. Для морской звезды орлиный глаз — бессмысленная трата жизненных ресурсов.
«Глаза не эволюционировали от плохих к идеальным, — объясняет Нильсон. — На первых порах они удовлетворяли несколько простейших зрительных потребностей, чтобы по мере развития начать выполнять многочисленные и невероятно сложные функции».
В своей модели Нильсон выделил четыре этапа эволюции глаза. Характерными особенностями каждого из этапов он выбрал не физические параметры зрительных органов, а те новые возможности, которые появлялись у их обладателей. На первом этапе зрение использовалось для измерения интенсивности падающего света, определения времени суток или оценки глубины вод, на которой находится животное. Тут достаточно одного фоторецептора. У гидры — небольшой сидячей родственницы медузы — глаз вообще нет, но на щупальцах есть фоторецепторы. Как показали Тод Окли и Дэвид Плачетски из Калифорнийского университета (Санта-Барбара), эти фоторецепторы связаны с ядовитыми стрекательными клетками, что, возможно, помогает гидре реагировать на тени проплывающих мимо потенциальных жертв (и выстреливать в них ядовитыми гарпунами) или ждать прихода ночи, когда добыча гидру не видит и сама плывет в щупальца. На второй стадии животные получают способность определять, откуда падает свет, — их фоторецепторы приобретают экранирующую выстилку (слой пигментных клеток), блокирующую часть световых лучей. Рецепторы позволяют своему хозяину увидеть однопиксельную картину мира, но и этого достаточно, чтобы выбрать направление движения к источнику света или, наоборот, — в тень убежища. Так ориентируются личинки многих морских животных. На третьем этапе фоторецепторы с пигментной выстилкой группируются в «глазки», каждый из которых смотрит в свою сторону. Обладатели таких глаз способны различать информацию, поступающую с разных направлений, и сводить ее в единую «картинку» — довольно размытую, но все же позволяющую составить представление об окружающем мире. Это поворотный момент, когда животные не просто улавливают световые сигналы, а начинают воспринимать зрительные образы, — момент появления настоящих глаз. Существа с таким зрением способны найти убежище (как морская звезда) или избежать столкновения с препятствием (как кубомедузы). Но настоящая эволюция глаз начинается на четвертом этапе. С появлением линзы-хрусталика, способной фокусировать световые лучи, зрение обретает резкость и остроту. «На этой стадии список зрительных функций разрастается до бесконечности», — подытоживает Нильсон.
Рецепторы позволяют своему хозяину увидеть однопиксельную картину мира, но и этого достаточно, чтобы выбрать направление движения к источнику света или, наоборот, — в тень убежища. Так ориентируются личинки многих морских животных. На третьем этапе фоторецепторы с пигментной выстилкой группируются в «глазки», каждый из которых смотрит в свою сторону. Обладатели таких глаз способны различать информацию, поступающую с разных направлений, и сводить ее в единую «картинку» — довольно размытую, но все же позволяющую составить представление об окружающем мире. Это поворотный момент, когда животные не просто улавливают световые сигналы, а начинают воспринимать зрительные образы, — момент появления настоящих глаз. Существа с таким зрением способны найти убежище (как морская звезда) или избежать столкновения с препятствием (как кубомедузы). Но настоящая эволюция глаз начинается на четвертом этапе. С появлением линзы-хрусталика, способной фокусировать световые лучи, зрение обретает резкость и остроту. «На этой стадии список зрительных функций разрастается до бесконечности», — подытоживает Нильсон.
Такое разнообразие новых возможностей для восприятия окружающей среды вполне могло быть толчком к бурному видообразованию, то есть к кембрийскому взрыву. Внезапно в системе «хищник — жертва» появляется новый элемент — и, вместо того чтобы вынюхивать, пробовать на вкус или искать на ощупь возможную добычу, хищники обрели способность выслеживать ее на расстоянии. Началась «гонка вооружений», в результате которой животные резко увеличились в размерах, стали подвижнее и обзавелись защитными панцирями и шипами.
Развитие органов зрения тем временем продолжалось. Все основные типы глаз, встречающиеся в наши дни, появились в кембрийском периоде, хотя и в примитивном виде. Конечно, с тех пор они приобрели множество новых функций. Скажем, глаза самцов насекомых поденок выглядят так, будто на небольшой фасеточный глаз «наклеили» еще один, огромный, основная задача которого — неустанно всматриваться в небо в поисках самок. У рыб-четырехглазок, нетрудно догадаться, каждый глаз разделен на две части, одна из которых находится над водной гладью и следит за происходящим в небе, а другая смотрит вниз, выискивая добычу и отслеживая хищников. Человеческий глаз отлично приспособлен для выполнения разных задач, он достаточно быстро воспринимает информацию, отлично улавливает контрасты, а по остроте зрения человека превосходят лишь хищные птицы — недаром говорят «глаз как у орла».
У рыб-четырехглазок, нетрудно догадаться, каждый глаз разделен на две части, одна из которых находится над водной гладью и следит за происходящим в небе, а другая смотрит вниз, выискивая добычу и отслеживая хищников. Человеческий глаз отлично приспособлен для выполнения разных задач, он достаточно быстро воспринимает информацию, отлично улавливает контрасты, а по остроте зрения человека превосходят лишь хищные птицы — недаром говорят «глаз как у орла».
Эволюция. Почему глаза у нас спереди?
- Джейсон Г. Голдман
- BBC Future
Автор фото, Thinkstock
Почему глаза у нас расположены не по бокам головы, а смотрят вперед? Отчасти это связано с необходимостью воспринимать трехмерные изображения, но корреспондент
BBC Future обнаружил и другие причины.
Вы когда-нибудь обращали внимание, что большинство животных в зоопарке можно отнести к одной из двух групп? У одних глаза находятся по бокам головы (это куры, коровы, лошади, зебры), а у других они посажены ближе и расположены спереди (в эту группу входят обезьяны, тигры, совы и волки). Сами посетители зоопарка — люди — очевидно, относятся ко второй группе. С чем же связано это различие?
Расположение глаз — это всегда некий компромисс. Когда глаза находятся спереди, каждый из них посылает в мозг изображение со своего угла зрения, и за счет наложения этих изображений друг на друга человек воспринимает глубину. Животные, глаза у которых расположены по бокам, не способны видеть третье измерение, зато обзор у них гораздо шире.
Автор фото, Thinkstock
Подпись к фото,
У некоторых черепах глаза находятся по бокам, но мозг обрабатывает зрительную информацию так, как если бы глаза у них смотрели вперед
Вероятно, расположение глаз формировалось у разных животных по-разному. К примеру, у некоторых черепах глаза находятся по бокам, но мозг обрабатывает зрительную информацию так, как если бы глаза у них смотрели вперед, — возможно, это связано с тем, что когда черепахи втягивают голову под панцирь, их глаза воспринимают свет только спереди, как будто они расположены в передней части головы. Но почему у нашей ветви эволюционного древа — у приматов — глаза оказались спереди? Тому есть множество объяснений.
Но почему у нашей ветви эволюционного древа — у приматов — глаза оказались спереди? Тому есть множество объяснений.
В 1922 году британский офтальмолог Эдвард Тричер Коллинз писал о том, что ранним приматам требовалось такое зрение, которое “позволяло бы им раскачиваться и точно перепрыгивать с ветки на ветку… хватать пищу руками и подносить ее ко рту”. Поэтому, решил ученый, в процессе эволюции у них развилась способность оценивать расстояние.
В последующие десятилетия гипотеза Коллинза неоднократно пересматривалась и уточнялась, но суть ее в течение долгого времени оставалась неизменной: в процессе эволюции глаза у наших предков переместились вперед, чтобы точно оценивать дистанцию при перепрыгивании с дерева на дерево. Цена ошибки при определении расстояния между деревьями действительно была немалой. “Расплатой за просчет было падение с высоты в несколько метров на землю, кишащую плотоядными зверями”, — написал в 1991 году специалист по визуальной психотерапии Кристофер Тайлер.
Автор фото, Thinkstock
Подпись к фото,
Попугаи обладают панорамным зрением
Слабое место гипотезы Коллинза состоит в том, что у многих животных, которые селятся на деревьях, — например, у белок, — глаза расположены по бокам. Поэтому в 2005 году американский биолог и антрополог Мэтт Картмилл предложил другую гипотезу, исходя из особенностей зрения хищников, способных очень хорошо оценивать расстояние. По мнению Картмилла, это позволяет им выслеживать и ловить добычу, будь то леопард, крадущийся за газелью, ястреб, цепляющий когтями зайца, или один из приматов, хватающий с ветки какое-нибудь насекомое. Ученый счел это объяснение весьма изящным, поскольку оно позволяло понять и другие эволюционные изменения, характерные для приматов. К примеру, ранние приматы в охоте полагались на зрение, а не на обоняние. Картмилл решил, что ухудшение обоняния было побочным эффектом сближения глаз: просто для носа и для нервов, связывающих его с мозгом, осталось не так много места — все пространство было занято глазами.
Американский нейробиолог Джон Оллман подхватил гипотезу Картмилла и доработал ее на основе сведений о ночных хищниках — ведь не у всех хищных животных глаза расположены спереди. У кошек, приматов и сов они действительно находятся в передней части головы, а у мангустов, тупай и мухоловок — по бокам. Вклад Оллмана в развитие этой гипотезы состоит в предположении о том, что такое зрение необходимо тем, кто охотится ночью, — например, кошкам и совам, — потому что впереди глаза воспринимают свет лучше, чем по бокам. Ранние приматы как раз охотились по ночам и, возможно, именно благодаря этому пристрастию к ночной охоте у всех их потомков, в том числе у людей, глаза расположены спереди.
Автор фото, Thinkstock
Подпись к фото,
У хищников, таких как этот леопард, глаза расположены спереди, чтобы лучше видеть добычу
У американского нейробиолога-теоретика Марка Чангизи возникло еще одно объяснение. В 2008 году он опубликовал в “Журнале теоретической биологии” (США) статью о “рентгеновском зрении”, предположив, что расположенные впереди глаза позволяли нашим предкам, жившим в лесу, видеть сквозь плотную листву и тесное переплетение веток. Громкое название “рентгеновское зрение” происходит от любопытного явления, описанного Чангизи: “Если держать палец перед глазами в вертикальном положении, фиксируя взгляд на каком-нибудь предмете, расположенном позади пальца, в мозг поступят два изображения пальца, и оба они будут прозрачными”. Таким образом, получается, что человек может “видеть сквозь” палец, как с помощью рентгеновских лучей.
Нагромождение деревьев в лесу мешает видеть только крупным животным, таким как приматы. Более мелкие, например, белки, не испытывают таких затруднений, поскольку их небольшая голова может легко протиснуться между ветвями и листьями. Крупным животным, которые живут не в лесу, тоже вполне достаточно глаз, которые расположены по сторонам.
Автор фото, Thinkstock
Подпись к фото,
Расположенные впереди глаза позволяли нашим предкам, жившим в лесу, видеть сквозь плотную листву и тесное переплетение веток
Таким образом, причина того, что глаза у нас находятся спереди, еще не установлена.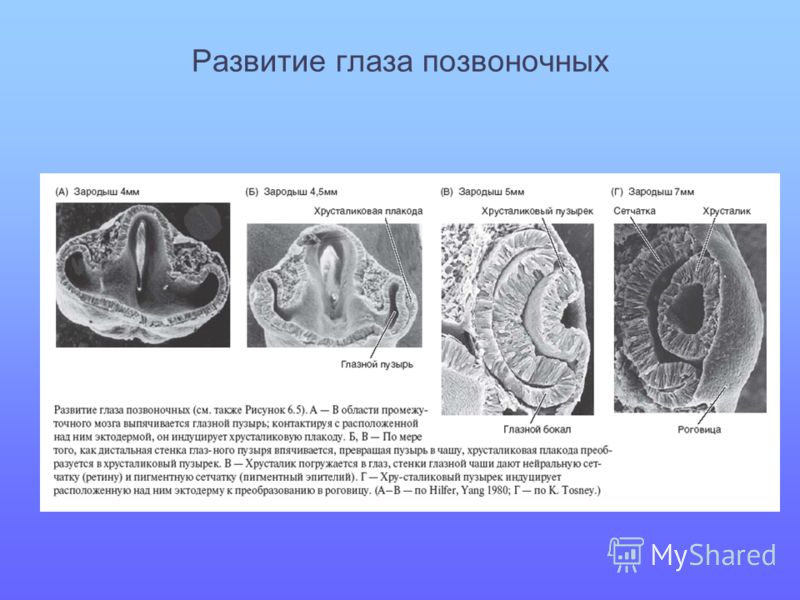 У каждой гипотезы есть свои сильные и слабые стороны. Но независимо от того, зачем нам потребовалось такое зрение — чтобы прыгать с ветки на ветку, ловить вкусных жучков или видеть сквозь листву — очевидно, что такое расположение глаз связано с жизнью среди деревьев.
У каждой гипотезы есть свои сильные и слабые стороны. Но независимо от того, зачем нам потребовалось такое зрение — чтобы прыгать с ветки на ветку, ловить вкусных жучков или видеть сквозь листву — очевидно, что такое расположение глаз связано с жизнью среди деревьев.
эволюция сложного органа Государственный Дарвиновский музей
29 июля 2017 — 1 октября 2017
Расположение: Выставочные залы музея / 3 этаж / Выставочный зал №7
Новый проект Дарвиновского музея «Эволюция сложных органов»
стартует с выставки о самом совершенном в мире оптическом приборе – глазе. Из
витрин на зрителя устремлены сотни пар глаз животных: от острого взора орла до подслеповатых
глазков медузы. Посмотрите на один и тот же пейзаж глазами человека, лошади,
вальдшнепа, сокола и стрекозы, обманите свой мозг оптическими иллюзиями и
начните завидовать осьминогам и кальмарам – ведь их глаза совершеннее наших!
Живые существа обладают богатым ассортиментом зрительных
приборов: от простых светочувствительных пятен на теле дождевого червя
до острого зрения орла. Является ли человеческий глаз вершиной «оптической
пирамиды»? Этому вопросу посвящена первая часть выставки. Здесь наш главный
зрительный орган разобрали на хрусталик, роговицу и сетчатку, разложили на
пиксели и фоторецепторы, измерили поле зрения и… обнаружили, что глаз человека
далёк от идеала! Этот ошеломляющий вывод можно проверить на практике в
интерактивной зоне – обмануть свое зрение при помощи оптической иллюзии или повторить
любимый эксперимент Людовика XIV по обнаружению «ахиллесовой пяты» глаза – слепого пятна.
Если мы используем глаза для изучения окружающей среды, то
для чего нужно зрение остальным представителям царства животных? Обратите
внимание на наглядную ленту эволюции зрения: от простейших «световых датчиков» медузы до сложных фасеточных глаз стрекозы. Каждый
этап – новая возможность для восприятия мира и новый инструмент для выживания. В
В
этом разделе выставки вы познакомитесь со «зрячими» простейшими, посмотрите в
глаза членистоногим, изучите особенное зрение птиц. Знали ли вы, например, что
глаза мухи фиксируют до 300 «кадров» в секунду, а человек лишь 50? Или что
чомга, пернатая любительница подводной охоты, видит одинаково хорошо как в
воздухе, так и под водой? Но самый большой удар по человеческому самолюбию
нанес… кальмар! Зрение этого головоногого моллюска примерно в 10 раз
чувствительней нашего.
Животные способны удивить не только разнообразием строения
глаз, но и их числом, внешним видом и расположением.
Многочисленные лазурные глазки атлантических гребешков, горизонтальные зрачки
альпийской горной козы и водяные глаза наутилуса – вот неполный перечень
эпатажных оптических приборов, представленных на выставке.
Актуальные мероприятия
Развитие лазерной коррекции зрения
О том, какой путь лазерная коррекция зрения прошла за последние годы и каких успехов достигла сегодня.
Когда в медицине возникла идея корректировать зрение манипуляциями на глазу, а не внешними приборами?
Этот вопрос изучали давно — с тех пор, как стал понятен принцип работы глаза. Роговица — это линза. Причем очень сильная. Ее оптическая сила 40-45 диоптрий. Вторая линза в глазу — это хрусталик. У него оптическая сила в два раза меньше. Соответственно, изображение после прохождения этих основных линз должно точно фокусироваться на сетчатке. При несовпадении каких-либо параметров получается размытое изображение, человек видит нечетко.
То, что в таких случаях для коррекции надо менять геометрию глаза, было понятно давно. В начале двадцатого века пытались уменьшить длину глаз (фокусное расстояние) делая совершенно варварские (по нынешним меркам) операции, когда швами стягивали глаз. От таких экспериментов очень быстро отказались из-за сопутствующих проблем. Более разумный и продуктивным, оказался путь по изменению оптических параметров.
От таких экспериментов очень быстро отказались из-за сопутствующих проблем. Более разумный и продуктивным, оказался путь по изменению оптических параметров.
Например, если заменить хрусталик (вторая линза) и на его место поставить искусственный с другой оптической силой, то можно изменить оптическую систему глаза до нужных параметров и получить четкое изображение на сетчатке.
Еще проще оказалось корректировать оптическую силу первой линзы — роговицы. Например, если отклонение зрения от нормы -3, а оптическая сила имеющейся роговицы 45, то подкорректировать надо лишь немного: от 45 до 42. Самый простой способ — сделать очки допустим со стеклами -3.0, более высокотехнологично, контактные линзы и наконец сделать операцию которая раз и навсегда уменьшит оптическую силу роговицы. Как бы “встроить” туда контактную линзу, но при этом физически в роговицу ничего инородного не прибавляется.
С изобретением лазеров это удалось осуществить?
Эксперименты с роговицей велись давно. Советский союз— родина широкого применения Передней Радиальной Кератотомии. Это операция, когда на периферии роговицы делают насечки алмазным ножом, центр роговицы при этом уплощается как раз так как будто туда “встраивается” отрицательная контактная лиза. Этот метод помог множеству людей жить полноценной жизнью без очков и контактных линз. Но метод так же имел и много нареканий: основные из них это: долгое восстановление, недостаточная предсказуемость, сложное заживление рубцов, вторичный астигматизм и др.
Первые эксимерные лазеры появились в 80-х годах прошлого века и были очень простые — они испаряли ткань. Использовались как японские Nidek, так и советские лазеры «Профиль-500». Ими уже можно было обточить роговицу глаза, придав ее новые оптические параметры. Этот метод назывался ФРК — фоторефракционная кератэктомия. Но оказалось, что и он имеет очень много минусов.
Роговица покрыта эпителием, где много нервных окончаний, это вызывает достаточно долгое (2-3 дня) заживление и не самые приятные ощущения в виде слезотечения и светобоязни. Кроме того возможно появление помутнений в роговице при операциях с большой степенью близорукости ( более 5 диоптрий).
Кроме того возможно появление помутнений в роговице при операциях с большой степенью близорукости ( более 5 диоптрий).
Сейчас ФРК не используют?
Редко. Но все же существуют ситуации, когда именно ФРК имеет преимущества, а именно в случаях так называемой “тонкой роговицы”. Нужно только понимать, что восстановление зрения займет время.
Как дальше шла эволюция методов?
Как и везде, в сторону увеличения точности и уменьшения инвазивности. Следующим методом стал LASIK (Laser-Assisted in Situ Keratomileusis). Предложен в США в конце 90-х годов двадцатого века.
Суть метода в том, что поверхностные самые чувствительные слои роговицы почти исключаются из воздействия. Специальным механическим кератомом делается неполный срез поверхностных слоев роговицы. Затем этот верхний слой приподнимается и эксимерный лазер всю свою процедуру (обтачивание за счет выпаривания) проводит на нижних слоях. Затем лоскут поверхностных слоев роговицы укладывают на место, где он достаточно быстро прирастает на свое место. Восстановление после такой операции значительно быстрее (день, два) и неприятных ощущений по сравнению с ФРК на порядок меньше.
Какие нарекания к данному методу?
Механическое срезание кератомом оказалось не очень точной процедурой. Допустим, средняя толщина роговицы около 500 микрон. Нам для безопасности глаза необходимо оставить 300 микрон ткани роговицы интактными. Еще около 100 микрон оставляют на крышку (верхний слой роговицы). Остается самые важные 100 микрон рабочей толщины в которой и вытачивается невидимая контактная линза. По расчетам, минимум 15 микрон нужно для коррекции на каждую диоптрию. Так вот когда счет идет на микроны механика дает большие погрешности. Какие бы ни делали прекрасные ножи кератомы — толщина среза все время разная.
Допустим выставляем 100 микрон толщину среза, а на деле он может получиться от 80 до 120 микрон. Во многих ситуациях сегодня это уже неприемлемо. Профиль среза микрокератомом тоже не идеален, кроме того, есть и другие отрицательные факторы.
В методах следующего поколения удалось эту трудность преодолеть?
Следующим этапом эволюции стал ФемтоLASIK. Принцип действия такого лазера — это микро-взрыв. Наверняка, все видели сувениры из оргстекла, где внутри куба или шара сделана объемная картинка — это сделано фемто-лазером. Фемто лазер проходит через прозрачную среду и на заданном расстоянии взрывает микро пузырек. Каждый пузырек меньше микрона, но общая сеть микровзрывов образует срез.
Начало внедрении технологии относится к 2005-2010 годам. Как и многое в офтальмо хирургии, эти технологии пришли из Германии. Условия использования импортных фемто лазеров построены так, что за каждую процедуру мы должны платить производителю оборудования. То есть каждую процедуру есть одна специальная насадка (стоимость которой сама по себе ничтожна), но на нее надо купить лицензию.
Получается, что фемтоLASIK выходит дороже из-за особых условий лицензии?
Сегодня это касается только фемтолазеров, эксимер-лазеры можно использовать без такой лицензии.
ФемтоLASIK дал значительное увеличение точности?
Да. Погрешность стала 2-3 микрона. То есть теперь, если мы хотим сделать разрез 100 микрон — мы именно его и получим. Кроме того, форма среза фемтолазером тоже поменялась и теперь ее стало значительно легче поставить на место и труднее сместить куда-либо.
Принцип «крышечки» из верхнего слоя роговицы остался. Первым работает фемтолазер делает крышечку, затем она на время поднимается. После этого на внутренних слоях роговицы, уже другим лазером, эксимерным, формируется как бы корригирующая контактная линза. Потом крышечку ставят на место, где она и прирастает. Заживление достаточно быстрое. Через сутки уже нет особых ощущений.
Какая технология по лазерной коррекции зрения на сегодняшний день, в 2020 году, считается самой прогрессивной?
Новейшая и безопасная – Relex Smile. Это технология только фемтосекундного лазера, но срезы проводятся по-другому. При Relex Smile в слоях роговицы сразу формируется отрицательная корригирующая линза. Она в какой-то мере похожа на настоящую контактную линзу, сделанную из ткани роговицы, но значительно тоньше ее.
При Relex Smile в слоях роговицы сразу формируется отрицательная корригирующая линза. Она в какой-то мере похожа на настоящую контактную линзу, сделанную из ткани роговицы, но значительно тоньше ее.
После того линза сформирована, лазер делает микроразрез (около 2 мм) через который мы и достаем этот лишнюю ткань роговицы в виде линзы. То есть получается один фемтолазер делает все то же самое что и при ФемтоLASIK, но это происходит под крышечной которую теперь не надо поднимать. За счет того, что микроразрез теперь 2 мм., а не 20 мм как при ФемтоLASIK процесс восстановления еще более ускоряется.
Метод Relex Smile позволяет корригировать близорукость до -11.0 диоптрий и астигматизм до – 6.0.
Какая реабилитация после коррекции близорукости по методу Smile?
Восстановление после процедуры занимает всего пару часов. Даже периода слезотечения, по сути, нет.
Обычно после эксимер-лазерных операций некоторое время (иногда до месяца) восстанавливается нормальный уровень увлажнения глаза, нужно капать специальные капли по типу искусственной слезы. При Smile эта необходимость как правило значительно меньше. К тому же здесь отсутствует возможность смещения лоскута в период первичного восстановления., то есть ограничения полностью отсутствуют, можно хоть боксом сразу заниматься. Ну и самое главное, острота зрения восстанавливается тоже очень быстро. Обычно больные нам рассказывают, что уже через 1-2 часа зрение становится резким.
У подавляющего числа наших пациентов, на следующий день зрение единица.
Говорят, что нормальная острота зрения — это единица. А можно ли сделать зрение выше единицы?
Здесь понадобится экскурс в физику зрительного аппарата. Но если вкратце, зрение «единица» — это предельный угол, под которым нормальный человеческий глаз может различить две точки. Связано это с тем, что сетчатка работает, как матрица у фотоаппарата, так вот, различимость предметов – зависит от расстояния между пикселями на матрице или еще от концентрации из на единицу площади. Сетчатка человеческого глаза физически насыщенна так этими пикселями что 1.0 “единица” по проверочной таблице — это норма.
Сетчатка человеческого глаза физически насыщенна так этими пикселями что 1.0 “единица” по проверочной таблице — это норма.
Хотя известны уникальные случаи, когда некоторые люди без оптики могли различать спутники у планет, острота зрения у них была значительно больше единицы.
Что касается операций, то при создании идеальной глазной оптики (исправления лазером всех природных неровностей роговицы) острота может стать 1.2, и даже иногда 1.5.
Часто после хирургической коррекции зрения от пациентов можно услышать вопрос: «А у меня зрение больше не упадет?» Скажите, может ли близорукость снова прогрессировать?
Такая вероятность существует. Ведь мы никоим образом не меняем ничего в глазу, мы лишь встраиваем внутрь глаза как бы контактную линзу. При этом прогрессирование близорукости (увеличение длины глаза) в принципе возможно. Глаз живет своей жизнью. Он может, например, увеличиваться. Собственно, поэтому мы делаем операции операцию по коррекции зрения после 18 лет, когда прогрессирование уже закончилось.
Лазерная коррекция предпочтительна на стабилизированной близорукости, но в некоторых случаях, через годы, если произошло прогрессирование мы можем провести докоррекцию и снова получить прекрасное зрение.
Тем, кто сделал операцию Smile, нужно потом в течение жизни регулярно проверяться у офтальмолога?
Раз в год желательно осматривать глазное дно у специалиста. Это рекомендация актуальна для любого человека, имеющего близорукость, делал он лазерную коррекцию зрения или нет – не имеет значения. Так как миопический глаз растянут, на периферии сетчатки могут появляться участки истончения, и дистрофии. Обработка этих участков лазером по необходимости, значительно снижает риски каких-либо неприятностей с сетчаткой.
Можно ли делать коррекцию зрения во время беременности или периода лактации?
Необходимо определятся по срокам беременности. Но сегодня подход такой что почти во время всего периода беременности и лактации можно делать Smile.
Осложнения на Smile возможны?
Теоретически, да, как и после любой манипуляции. Поэтому важно выбирать опытных специалистов. В нашей клинике уже сделано сотни операций Smile, без каких либо осложнений. Бывают редчайшие случаи, когда есть противопоказания к Smile. Это выясняется на этапе диагностики, который обязательно проводится перед лазерной коррекцией зрения. Если есть противопоказания к Smile, тогда используем ФемтоLasik или даже ФРК.
Какой диапазон коррекции у метода Смайл?
Если говорить о близорукости, то от -1 до -10. Астигматизм до -5.0.
Для более сильных отклонений есть другие технологии: например, факичные линзы. Но это предмет отдельного разговора. Главное, что следует знать пациенту, сегодня существуют методики, позволяющие решить большой спектр проблем со зрением.
Мы все время говорим о близорукости. А как быть с дальнозоркостью? Лазерная коррекция зрения применима только для миопического глаза?
Дальнозоркость — более сложно технологически решаемая проблема. Smile здесь, к сожалению, пока не работает. Можем использовать Femto Lasik или ФРК. Он позволяет корректировать до 6 диоптрий.
Что такое дальнозоркость? Это значит, глаз по длине меньше, чем необходимо для четкого фокуса на сетчатку. С дальнозоркостью еще такой физиологический момент: хрусталик настолько гибкое тело, что в молодости он может компенсировать до 3-4 диоптрий дальнозоркости. А после 40 лет, когда теряется гибкость связок хрусталика, появляется дальнозоркость. Иногда в таких случаях мы меняем хрусталик на искусственный и получаем прекрасные результаты.
Много ли сейчас в Москве клиник, предлагающих технику Smile?
Около 6 клиник, включая государственные. Но уверен, что эта технология скоро будет везде. Тем более, что на подходе лазеры-конкуренты.
Сколько фирм сейчас выпускают такие лазеры?
Сегодня лазеры для проведения Smile делает только одна фирма – Carl Zeiss. Эта немецкая фирма пока является монополистом и из-за этого ставит свои финансовые условия.
Но сейчас на подходе уже лазеры других фирм, тоже из Германии: Zimmer и Schwind. Когда они выйдут на рынок, им придется выбирать другие названия для своих методов, так как Smile – уже запатентованная технология. Но принцип работы у них будет аналогичный.
Что будет дальше? Есть ли недостатки у Smile? В какую сторону будет развиваться лазерная коррекция зрения?
За эти годы много раз улучшалось компьютерное обеспечение, но сама процедура остается неизменна. Smile показала себя, как лучшая технология, к которой нет особых нареканий. Когда изобрели первый лазер Smile, производители сделали исследование на референтной группе: набрали добровольцев (60 человек по всей Германии) протестировали и сделали им коррекцию Smile. Это было 10 лет назад.
В этом году опубликовали их нынешнее состояние — у всех рефракция роговицы никак не изменилась со времени проведения операции.
Эволюция конечно будет, еще точнее, еще удобнее, еще более предсказуемо, но Smile это здорово и это надолго.
Эволюция Органа Зрения Человека | Подольский центр зрения
Чарльз Дарвин подарил всему миру уникальнейшую теорию эволюции человеческого организма. Объяснив изменения всего организма в целом, он никогда не переставал удивляться функционированию и сложности строения человеческого органа зрения. В данной статье мы расскажем вам об уникальности и эволюции глаз.
Человеческий глаз — система поистине уникальная. Человек с легкостью может разглядывать расположенный вдали предмет и тут же моментально фокусироваться на близлежащих объектах, познавать окружающий мир, различать огромное количество цветов и еще большее количество их оттенков. Ведь именно благодаря своему зрению человек может воспринимать от 80 до 90 % информации.
Однако человеческий глаз не всегда был таким как сейчас — он эволюционировал. А началось все это более чем пятьсот миллионов лет назад. Маленькое светочувствительное пятнышко начало свое развитие на теле одноклеточных. Это пятнышко было концентрацией белков, которые были восприимчивы к свету. Они оживлялись, когда им встречались источники света, а стало быть, пища или тепло. У плоского червя планарии можно наблюдать более развитый механизм подобного глазка, который расположен изолированно в эпидермисе, он способен различать свет и еще его направление.
Это пятнышко было концентрацией белков, которые были восприимчивы к свету. Они оживлялись, когда им встречались источники света, а стало быть, пища или тепло. У плоского червя планарии можно наблюдать более развитый механизм подобного глазка, который расположен изолированно в эпидермисе, он способен различать свет и еще его направление.
После многих тысячелетий, у многих животных такое скопление белков погрузилось внутрь, а их просвет стал более узким. В результате появился так называемый «пинхольный эффект», с помощью которого уменьшилось искажение и увеличилось разрешение видимой информации. Поскольку теперь в глаз стал попадать только тонкий луч света.
Несмотря на то, что пинхольный глаз уже позволял воспринимать изображение, началом эволюции человеческого глаза все же считается линза. Офтальмология предполагает, что сама линза — это сильное скопление прозрачных клеток, защищающих глаз от различных инфекций. Из-за этого сам глаз немного позже наполнился жидкостью, что в результате улучшило резкость изображения и его чувствительность. Особый механизм сформировали эти прозрачные белки, благодаря которому на сетчатке свет фокусировался в одной точке. Адаптивность глазу обеспечивает линза, а именно она изменяет свою кривизну для дальнего и ближнего изображения. Такая система пинхольной камеры с линзой и есть прародителем человеческого глаза. Намного позже появились склера, радужка и слезные железы.
Огромное значение в развитии человеческого глаза имеет эволюция головного мозга, именно с его помощью человек способен обрабатывать миллионы цветных и резких изображений.
На сегодняшний день ученым-офтальмологам очень многое известно о функционировании и развитии глаз. Благодаря этим знаниям врачи ведут непрерывную борьбу с различными глазными проблемами и дефектами. Также ведется постоянная работа по разработке синтетических имплантов, ведь они смогли бы подарить зрение слепым и слабовидящим людям. Но сможет ли человеческий ум превзойти саму Природу? Время покажет.
Эволюция глаз в аниме и их особенности
Все началось в 60-х
Человек, которому мы обязаны этим фирменным аниме стилем, является не кто иной, как Осаму Тедзука, коего в принципе величают крестным отцом аниме. Во многом Тедзука вдохновлялся творчеством Уолта Диснея и такими работами как «Бэмби» и дизайном персонажей из Мики Мауса, например, Дональдом Даком.
Однако если проводить параллели между дизайном персонажей, японцы ушли дальше своих американских коллег и в аниме Тедзуки не только глаза, но и причёска, черты лица и одежда во многом описывали архетип.
В целом дизайн персонажей играет роль в передаче эмоций любой конкретной сцены. Такой фокус и тщательное внимание к дизайну глаз само по себе оказывается эффективным. Тедзука и многие его последователи понимали, что глаза могут создать настроение для эмоциональных сцен, чем и запустили цепочку эволюции глаз в аниме.
Архетипы по глазам
Так, глаза стали полностью олицетворять того или иного персонажа, выступая идентификатором. Там, где в американской анимации главным был внешний вид, восточная анимация концентрировалась в первую очередь на глазах. Для начала давайте пройдемся по основным моментам того, как на сегодня аниме глаза стали дословно зеркалом души.
Персонаж с очень большими глазами и ртом в виде пиалы, если можно так выразиться, является невинным и смотрит на мир с восхищением, и неподдельной радостью. Очень часто такое можно встретить в моэ, так как этот жанр возводит милоту персонажей в апогей.
В противовес им выступают персонажи с зауженными глазами с острыми контурами. Чаще всего такие персонажи злые или как минимум циничные. Но за этими суровыми аниме глазами они скрывают частичку нежности или любви, как было, например, с Кёном из «Меланхолии Харухи Судзумии».
Однако у мужских персонажей в целом глаза могут меняться с возрастом. У женских персонажей также, но с героями мужчинами так происходит чаще в силу японской культуры. С возрастом большие и круглые глаза мальчика могу заостряться, как некий символизм, что он вырос. Например, вам просто стоит сравнить глаза Саске до того как Итачи устроил резню в клане Учиха и уже после того, как он перешел на сторону Орочимару.
С возрастом большие и круглые глаза мальчика могу заостряться, как некий символизм, что он вырос. Например, вам просто стоит сравнить глаза Саске до того как Итачи устроил резню в клане Учиха и уже после того, как он перешел на сторону Орочимару.
При этом подобные персонажи, сохраняя в себе ту самую частицу добра, могут с возрастом опять стать добрее или как минимум мудрее. Их глаза хоть также заужены, уже не так остры, а вокруг них можно увидеть складки.
Итог следующий – чем круглее становятся глаза персонажа, тем больше добрых качеств в нем проявляется. Также практика создания глаз полезна в выражениях реакции, помимо увеличения или уменьшения в размерах. Например, когда персонаж влюбляется или видит что-то, что ему нравится и он хочет это получить. Это особенно видно в Sailor Moon.
Чтобы показать абсолютную злость или даже безумие персонажа часто используют прием, когда глаза неподдельно большие, но сами зрачки сужены. И в некоторых случаях так олицетворяют еще и страх. В этом случае все зависит от выражения рта.
В противовес этому выступает прием, когда зрачки большие – такое используют, чтобы показать грусть и это самый коварный прием, заставляющий тебя сочувствовать.
И мой самый любимый момент – пустые зрачки, показывающие либо моральное истощение персонажа, подавленное состояние или же его полное изменение внутри.
Непосредственно эволюция глаз в аниме
Наглядно эволюцию глаз в аниме одним рисунком показал Наоки Сайто, аниматор и художник, занимающийся на протяжении многих лет играми серии Pokemon, и кому как не ему разбираться в том, как эволюционирует такой важный элемент дизайна персонажей как глаза.
По словам иллюстратора, изображение слева показывает, как глаза были нарисованы давно, с вертикальной длинной овальной формой и множеством пустого пространства вокруг зрачков. Портрет в середине показывает переход к более угловатым формам глаз, которые в целом больше, с акцентом на сильные темные ресницы. Справа у нас есть изображение, которое обычно используется сегодня, с меньшим количеством пустого пространства внутри глаза и функциями, которые являются более консервативными и сдержанными по сравнению с прошлым.
Справа у нас есть изображение, которое обычно используется сегодня, с меньшим количеством пустого пространства внутри глаза и функциями, которые являются более консервативными и сдержанными по сравнению с прошлым.
По наблюдениям людей из его твиттера изображение слева напоминает персонажей из аниме-франшизы Slayers, которая началась в 1989 году. Среднее изображение напоминает K-ON! Впервые персонажи появились в манге в 2007 году, а картинка справа выглядит как персонажи из Sound! Euphonium 2013-го.
И да, как по мне эти приведенные пользователями твиттера под его постом примеры идеально описывают переход. Особенно K-On!. Как по мне, жанр моэ во многом определил сегодняшний дизайн многих персонажей и их глаз, так как нашел некий баланс между безумными размерами прошлого и тем, что они приставляют сегодня.
Так что условно мы можем делить виды аниме глаз на три эпохи:
- Эпоха больших глаз и узких зрачков
- Эпоха больших глаз и больших зрачков
- Эпоха баланса между размером глаз и зрачков
Однако это в целом, и если рассматривать единичные случаи – это не так. Хочу сказать, что мы говорим в общих чертах.
Нарратив
Но независимо от того, как все сложилось, глаза в аниме рассказывают потрясающую историю эволюции персонажа, и мой любимый пример – это Саске, как я уже упоминал ранее. И он не единственный пример такой эволюции. Вспомните Кирицугу Эмия, из Fate/ или Кензо Тенму из Monster и Акане Цунемори из Psycho-Pass.
Некоторые аниме могут иметь более тонкий дизайн для аниме глаз их главных героев, но они по-прежнему включают в себя выразительность, которую можно встретить только в такой среде, как анимация, что объясняет, почему такие концепции в первую очередь анимированы.
Революция в мире дисплеев. Создана безопасная для глаз матрица тоньше LCD и контрастнее E Ink
, Текст: Эльяс Касми
TCL представила новый стандарт экранов NXTPAPER, превосходящий по многим параметрам LCD и Е Ink, но при этом не нуждающийся в подсветке и безвредный для глаз. Он предназначен для мобильных устройств, особенно для ридеров и планшетов, но TCL не говорит о сроках начала массового производства таких панелей.
Китайцы всех опередили
Компания TCL разработала новый тип дисплея, который, по ее заверению, существенно лучше популярных форматов панелей. Он получил название NXTPAPER, и TCL предлагает использовать его в мобильных устройствах.
В рамках премьеры NXTPAPER, прошедшей на международной выставке IFA 2020, представители TCL заявили, что такие экраны оптимально подойдут для использования в планшетных компьютерах и электронных книгах. Про применение их в смартфонах они не упоминали.
TCL из Китая выпускает широкий спектр устройств как под собственным брендом, так и под дочерними. Так, за устройства марки Alcatel отвечает именно она, плюс до недавнего времени ей принадлежала лицензия на выпуск смартфонов BlackBerry. О планах по отказу от бренда TCL заявила в феврале 2020 г., а в августе 2020 г., как сообщал CNews, лицензия была передана американскому стартапу OnwardMobility.
Лучше, чем Е Ink
По заявлению разработчиков, экраны NXTPAPER не мерцают во время работы и не излучают синий свет. За счет этих особенностей они не создают нагрузку на зрительный аппарат, однако этими же преимуществами обладают и панели на электронных чернилах, в том числе и новая E Ink Kaleido, способная отображать 4096 оттенков цветов.
TCL уверена в превосходстве их разработки над LCD и даже над Е Ink
TCL сравнила NXTPAPER с такими экранами, заявив, что их разработка обладает на 25% более высокой контрастностью. Также она отметила, что NXTPAPER могут использоваться для просмотра видео, чем экраны на электронных чернилах ввиду невысокой скорости обновления похвастаться не могут. Также TCL заявила, что экраны нового типа смогут формировать полноцветное изображение в разрешении Full HD (1920х1080 точек). Количество отображаемых ими оттенков она предпочла не раскрывать.
Также TCL заявила, что экраны нового типа смогут формировать полноцветное изображение в разрешении Full HD (1920х1080 точек). Количество отображаемых ими оттенков она предпочла не раскрывать.
LCD тоже не конкурент
Без сравнения ЖК-панелями тоже не обошлось – по официальным данным, матрицы NXTPAPER на 35% тоньше и на 65% более энергоэффективны.
Прототип планшета с экраном NXTPAPER
Определенное сходство у новинки TCL есть и с дисплеями OLED. Им, как известно, не требуется дополнительная подсветка в отличие от ЖК-матриц, и NXTPAPER тоже обходится без нее.
Независимость от подсветки здесь реализована путем использования специальных материалов с высокой отражающей способностью. По словам производителей, он «повторно использует естественный свет», то есть он отражает свет от внешних источников.
Годы разработки
В TCL заявляют, что специалисты компании потратили несколько лет на разработку данной технологии. Как долго они работали над ней на момент публикации материала известны не были.
Как виртуализация улучшает бизнес-показатели ЦОД
Инфраструктура
Нет информации и о том, на какой именно стадии разработки эта технология находится. Рассказав об устройствах, в которых экраны NXTPAPER могут применяться, представители TCL не рассказали в итоге, когда именно первые гаджеты с такими дисплеями поступят в продажу, как они будут отличаться по цене от тех же книжек с Е Ink и планшетов с IPS-матрицами и, главное, кто из производителей проявил интерес к этой технологии.
Существует вероятность, что TCL будет выпускать устройства с экранами NXTPAPER под имеющимися в ее распоряжении брендами, в том числе и под собственным. К примеру, вместе с этой технологией она показала планшеты TCL 10 TabMax и 10 TabMid, продажи которых начнутся в IV квартале 2020 г., но обе эти модели оснащены классическими IPS-дисплеями с подсветкой и определенной долей излучаемого синего света.
Потенциальный недостаток устройств NXTPAPER
Способность экранов NXTPAPER работать без подсветки может побудить производителей начать выпускать гаджеты, в которых она не предусмотрена вовсе. Таким образом, электронную книгу с таким дисплеем и тем более планшетный компьютер нельзя будет использовать в полной темноте, например, при чтении перед сном.
Таким образом, электронную книгу с таким дисплеем и тем более планшетный компьютер нельзя будет использовать в полной темноте, например, при чтении перед сном.
Первые ридеры с дисплеями на электронных чернилах тоже имели этот недостаток. Современные же модели, в подавляющем своем большинстве, поставляются с подсветкой, в некоторых случаях, даже с двумя оттенками.
Эволюция глаза | New Scientist
Они появились в мгновение ока и навсегда изменили правила жизни. На глазах жизнь была спокойнее и покорнее, в ней преобладали вялые, мягкотелые существа, валявшиеся в море. Изобретение глаза открыло путь к более жестокому и конкурентному миру. Vision позволил животным стать активными охотниками и спровоцировал эволюционную гонку вооружений, которая изменила планету.
Когда появились глаза?
Первые глаза появились около 541 миллиона лет назад — в самом начале кембрийского периода, когда действительно возникла сложная многоклеточная жизнь — у группы ныне вымерших животных, называемых трилобитами, которые были немного похожи на крупных морских мокриц.Их глаза были сложными, как у современных насекомых. И их появление в летописи окаменелостей поразительно внезапно. Предки трилобитов, жившие 544 миллиона лет назад, не имели глаз. Так что же произошло за этот волшебный миллион лет? Неужели глаза с их взаимосвязанным набором сетчатки, хрусталика, зрачка и зрительного нерва слишком сложны, чтобы появиться внезапно?
Как эволюционировали глаза?
Реклама
Сложность глаза долгое время была полем битвы эволюционистов.С тех пор, как Уильям Пейли в 1802 году придумал аналогию с часовщиком, в которой утверждалось, что такая сложная вещь, как часы, должна иметь своего производителя, креационисты использовали ее, чтобы привести «аргумент, основанный на дизайне». Они говорят, что глаза настолько замысловаты, что внушает доверие предположение, что они эволюционировали в результате отбора и накопления случайных мутаций.
Чарльз Дарвин был хорошо осведомлен об этом аргументе. В книге «О происхождении видов» он признал, что глаза были настолько сложными, что их эволюция казалась «в высшей степени абсурдной».Но он продолжал убедительно утверждать, что это только казалось абсурдом. Сложные глаза могли развиться из очень простых путем естественного отбора, если каждая градация была полезной. По словам Дарвина, ключ к разгадке состоит в том, чтобы найти в животном мире глаза средней сложности, которые продемонстрируют возможный путь от простого к сложному.
Эти промежуточные формы теперь найдены. По мнению биологов-эволюционистов, самому элементарному глазу потребовалось менее полумиллиона лет, чтобы превратиться в сложный глаз-«камеру», подобный нашему.
Первым шагом является развитие светочувствительных клеток. Это кажется тривиальным делом. У многих одноклеточных организмов есть пятна на глазах, сделанные из светочувствительных пигментов. Некоторые могут даже плыть к свету или от него. Такие рудиментарные светочувствительные способности дают очевидное преимущество в выживании.
Следующим шагом для многоклеточных организмов было сосредоточение своих светочувствительных клеток в одном месте. Участки светочувствительных клеток, вероятно, были обычным явлением задолго до кембрия, что позволяло ранним животным обнаруживать свет и ощущать, с какого направления он исходит.Такие рудиментарные органы зрения до сих пор используются медузами, плоскими червями и другими примитивными группами, и они явно лучше, чем ничего.
Первые глаза в природе
Простейшими организмами со светочувствительными пятнами являются гидры — пресноводные существа, родственные медузам. У них нет глаз, но они сжимаются в шар под ярким светом. Гидры интересны с эволюционной точки зрения, потому что их базовое светочувствительное оборудование очень похоже на то, что наблюдается у других эволюционных линий, включая млекопитающих.Он основан на двух типах белков: опсинах, которые меняют форму при попадании на них света, и ионных каналах, которые реагируют на изменение формы, генерируя электрический сигнал. Генетические исследования показывают, что все системы опсин-ионных каналов произошли от общего предка, подобного гидрам, что указывает на единое эволюционное происхождение всех зрительных систем.
Генетические исследования показывают, что все системы опсин-ионных каналов произошли от общего предка, подобного гидрам, что указывает на единое эволюционное происхождение всех зрительных систем.
Следующим шагом будет создание небольшого углубления, содержащего светочувствительные клетки. Это упрощает определение направления, откуда исходит свет, и, следовательно, распознавание движения.Чем глубже яма, тем резче дискриминация.
Дальнейшее улучшение может быть достигнуто за счет сужения отверстия ямы, чтобы свет попадал через маленькое отверстие, как в камере-обскуре. С помощью такого рода оборудования сетчатка глаза может разрешать изображения — значительное улучшение по сравнению с предыдущими моделями. Глаза с камерой-обскурой, лишенные линзы и роговицы, встречаются сегодня у наутилуса.
Последнее большое изменение — это усовершенствование линз. Вероятно, это началось с защитного слоя кожи, который вырос над отверстием.Но он превратился в оптический инструмент, способный фокусировать свет на сетчатке. Как только это произошло, эффективность глаза как системы визуализации резко возросла с 1 до 100 процентов.
Глаза такого типа до сих пор встречаются у кубозойных животных, очень подвижных и ядовитых морских хищников, похожих на медуз. У них 24 глаза, расположенные в четыре группы; 16 — это просто светочувствительные ямки, но одна пара в каждом кластере сложна, со сложной линзой, сетчаткой, радужной оболочкой и роговицей.
Трилобиты пошли несколько другим путем, развив сложные глаза с несколькими линзами. Но основная последовательность событий была той же.
Трилобиты были не единственными животными, которые наткнулись на это изобретение, хотя они были первыми. Биологи считают, что глаза эволюционировали независимо во многих, а возможно, и в сотнях случаев.
И какая разница. В слепом мире раннего кембрия видение было равносильно сверхдержаве. Трилобиты стали первыми активными хищниками, способными искать и преследовать добычу, как ни одно животное прежде. Неудивительно, что их жертвы эволюционировали. Всего несколько миллионов лет спустя глаза были повсюду, а животные стали более активными и ощетинились доспехами. Этот всплеск эволюционных инноваций — это то, что мы теперь знаем как кембрийский взрыв.
Неудивительно, что их жертвы эволюционировали. Всего несколько миллионов лет спустя глаза были повсюду, а животные стали более активными и ощетинились доспехами. Этот всплеск эволюционных инноваций — это то, что мы теперь знаем как кембрийский взрыв.
Однако прицел не универсален. Из 37 типов многоклеточных животных только шесть развили его. Но эти шесть, включая наш собственный тип, хордовые, а также членистоногих и моллюсков, являются самыми многочисленными, широко распространенными и успешными животными на планете.
Eye Evolution
Глаз
Глаз всегда был предметом споров для биологов-эволюционистов. Разница в строении глаз между пауком и человеком настолько разительна, что кажется нелогичным, что оба глаза должны быть связаны эволюционно. Поэтому предполагалось, учитывая огромное разнообразие структур глаза, встречающихся в природе, что глаз должен возникать случайно более 40 раз в течение эволюции. Чарльз Дарвин знал, что с помощью своей теории эволюции он смог объяснить, что если глаз был создан случайным образом, то давление отбора приведет к появлению разнообразия, наблюдаемого у разных видов.Но в этой логике была основная проблема. Противники теории Дарвина справедливо указывали, что кажется невозможным, чтобы такой совершенный орган, как глаз, мог эволюционировать случайно более 40 раз. Решение Дарвина заключалось в том, чтобы постулировать, что простой прототип глаза эволюционировал только один раз, очень рано в эволюции, который затем посредством естественного отбора мог дать начало разнообразию, присутствующему в природе. Современная молекулярная биология предоставила убедительные доказательства такой идеи.Гены, определяющие развитие глаз, по-видимому, сохраняются во всем животном царстве, что позволяет предположить, что рождение глаза было единичным событием в эволюции.
Глаза позволяют животным улавливать свет и преобразовывать его в электрический сигнал. Однако, пока свет улавливается нашими глазами, процесс «видения» изображения происходит в нашем мозгу. Глаз позвоночного называется глазом типа «перевернутая камера», поскольку он устроен так же, как фотоаппарат. Как и в фотоаппарате, линза служит для фокусировки света на сетчатке, которая представляет собой светочувствительную пленку в задней части наших глаз, в то время как радужная оболочка контролирует количество попадающего света.Сетчатка — это тонкая нейроэпителиальная ткань, состоящая из 3 клеточных слоев. Однако перевернутое имя не относится к тому факту, что изображение в конечном итоге проецируется на сетчатку вверх ногами, а скорее относится к расположению различных нейронов внутри сетчатки. Светочувствительные нейроны сетчатки, которые улавливают свет, находятся дальше всего от входящего источника света и, следовательно, направлены от света. Следовательно, свет должен сначала пройти через всю сетчатку, прежде чем он сможет быть захвачен фоторецепторами, светочувствительными нейронами, которые служат для запуска процесса зрения.Примеры не перевернутых (то есть «перевернутых») глаз типа камеры — это глаз головоногих моллюсков.
Сетчатка
Сетчатка состоит из трех клеточных слоев: внешнего ядерного слоя, внутреннего ядерного слоя и слоя ганглиозных клеток. Внешний ядерный слой содержит только фоторецепторы. В сетчатке позвоночных находятся два типа фоторецепторов: палочки и колбочки. Конические фоторецепторы отвечают за зрение при более ярком дневном свете и опосредуют цветовое зрение.Стержневые фоторецепторы в 1000 раз более чувствительны к свету и вызывают зрение при тусклом свете. Свет, который улавливается PR, преобразуется в электрический сигнал, который передается на биполярные клетки. Биполярные клетки расположены во внутреннем ядерном слое и передают сигнал ганглиозным клеткам, расположенным в слое ганглиозных клеток. Ганглиозные клетки — выходные нейроны сетчатки, которые проецируются в мозг. Другие типы клеток, обнаруженные во внутреннем ядерном слое, — это амакриновые клетки, горизонтальные клетки и клетки глии Мюллера (перевёрнутое изображение).Клетки глии Мюллера являются основным типом клеток глии в сетчатке. Они охватывают всю толщину сетчатки и играют важную вспомогательную роль для различных нейронов сетчатки. Клетки глии Мюллера также являются частью гематоэнцефалического барьера сетчатки. Амакриновые и горизонтальные клетки играют важную роль в модуляции первичного сигнала от биполярных клеток.
В человеческом глазу свет фокусируется на небольшом участке сетчатки, ямке, где возникает высокая острота зрения. Эта область хорошо приспособлена для дневного зрения и содержит только фоторецепторы колбочек.Примерно 95% фоторецепторов в остальной части сетчатки представляют собой палочки с центральным кольцом, богатым палочками, которое почти не содержит колбочек. За исключением ямки, отсутствующей у мышей, сетчатка человека и мыши очень похожа. Таким образом, мыши служат отличной моделью для изучения развития сетчатки и заболеваний.
Эволюция глаз и визуально управляемое поведение
Реферат
Морфология и молекулярные механизмы фоторецепторных клеток и глаз животных обнаруживают сложную структуру дупликаций и кооптации генетических модулей, что приводит к появлению ряда различных светочувствительных систем, которые имеют много общих компонентов, в которых явная гомология встречается редко.На основе молекулярных и морфологических открытий я обсуждаю функциональные требования к зрению и то, как они ограничили эволюцию глаз. Тот факт, что естественный отбор на глазах действует через последствия визуально управляемого поведения, приводит к концепции эволюции с разделением задач, когда сенсорные системы развиваются путем последовательного овладения сенсорными задачами. Я выделяю четыре ключевых нововведения, которые одна за другой проложили путь к развитию эффективных глаз. Эти инновации включают (i) эффективные фотопигменты, (ii) направленность через экранирующий пигмент, (iii) складывание мембраны фоторецептора и (iv) фокусирующую оптику.Предлагается соответствующая эволюционная последовательность, начинающаяся с ненаправленного мониторинга окружающей яркости и ведущая к сравнению яркостей в пределах сцены, сначала в режиме сканирования, а затем в параллельных пространственных каналах в глазах, формирующих изображения.
Ключевые слова: зрение, эволюция, глаз, зрительная задача, визуальная информация, фоторецептор
1. Предпосылки
(a) Введение
Новые молекулярные и генетические данные об эволюции глаза генерируются постоянно растущими темпами.Это привело к гораздо лучшему пониманию эволюции глаз, а также к частым конфликтам между данными разных типов или разных видов. В частности, морфологические данные иногда указывают в одном направлении, а молекулярные сигналы — в другом. Но оба типа данных в принципе одинаково достоверны и верны. Причина того, что разные правильные данные иногда кажутся противоречащими друг другу, конечно же, заключается в том, что наши интерпретации неверны или, по крайней мере, не полны. В этой статье я кратко рассмотрю информацию, наиболее актуальную для ранней эволюции глаза, и обсуждаю возможные сценарии в свете функциональных аргументов.
Без понимания того, как отбор руководил процессом эволюции, любой эволюционный сценарий будет неполным. Филогения генов или белков может сказать нам, что могло произойти на молекулярном уровне, а другие подходы, такие как исследования экспрессии и эксперименты с нокаутом, могут связать эти молекулярные изменения с функциями. Но чтобы понять, почему эволюционировали особенности или функции, нам нужны правдоподобные идеи об их выборе, чтобы мы могли объяснить, почему предполагаемым эволюционным путям отдавалось предпочтение в ущерб другим возможностям.В процессе эволюции сенсорных функций прием внешней информации будет только в том случае, если она будет использоваться для управления поведением или физиологией таким образом, чтобы оказывать положительное влияние на приспособленность.
Для анализа преимуществ сенсорных систем полезно ввести термин «сенсорная задача», в широком смысле определяемый как систематическая поведенческая или физиологическая реакция на конкретную полученную информацию. Самый простой смысл включает в себя единственную сенсорную задачу, связывающую конкретную полученную информацию с одним конкретным ответом.В эволюции сенсорной системы одна сенсорная задача также является очевидной отправной точкой. Помимо более поздних модификаций сенсорной задачи, последующее эволюционное развитие сенсорной системы будет связано с появлением новых задач. Таким образом, сенсорную эволюцию можно понимать как последовательное добавление сенсорных задач. Базовый эволюционный принцип дублирования с последующей модификацией был бы типичным механизмом для добавления новых задач к сенсорной системе.
Сенсорные задачи могут быть основаны на простых цепях рецептор-эффектор без промежуточной обработки, или они могут включать сравнения или память, требуя различной степени промежуточной обработки от простой до чрезвычайно сложной.При реконструкции сенсорной эволюции естественно предположить, что простые и менее сложные задачи возникли первыми, потому что они требуют наименьшего количества компонентов. По мере накопления сенсорных задач сенсорные органы должны будут перейти от предоставления конкретной информации, настроенной для одной или нескольких аналогичных задач, на предоставление более общей информации, охватывающей потребности широкого круга различных задач. Этот процесс увеличивает нагрузку на нервную систему, потому что, когда полученная информация постепенно становится более общей, возрастает потребность в сортировке и фильтрации информации для решения каждой отдельной задачи.
Каждая сенсорная задача требует, чтобы определенный стимул был связан с определенной реакцией. Чтобы это работало, необходимо, чтобы сенсорная система была построена так, чтобы она надежно реагировала при наличии определенного стимула, но игнорировала стимулы, не соответствующие конкретным требованиям. Таким образом, сенсорная система должна действовать как фильтр, пропускающий определенный стимул, но блокирующий всю остальную информацию. Эта концепция сенсорной фильтрации была разработана Венером (1987), который также ввел термин «согласованные фильтры», чтобы в совокупности включать сенсорные и нейронные свойства, которые вместе образуют фильтр, который соответствует конкретному информационному шаблону, вызывающему поведенческую реакцию.Таким образом, каждая сенсорная задача потребует определенного согласованного фильтра, и в простых сенсорных системах, поддерживающих только одну или несколько подобных сенсорных задач, эти фильтры могут быть реализованы в свойствах рецепторных клеток или конструкции сенсорного органа (например, по зрению см. Nilsson и др. .2005; Land & Nilsson 2006). Когда сенсорный орган эволюционирует, чтобы поддерживать большое количество сенсорных задач, согласованные фильтры должны постепенно переходить от сенсорного органа к нервной системе.В области зрения это означает, что можно ожидать эволюции от одноцелевых фоторецептивных систем с периферической фильтрацией к глазам общего назначения с центральной нейронной фильтрацией (Land & Nilsson 2006).
Сенсорные системы могут улучшить физическую форму только за счет реакции, которую они вызывают. И наоборот, поведение и физиологические реакции требуют, чтобы сенсорный контроль осуществлялся именно в условиях, когда реакция улучшает ситуацию. Таким образом, сенсоры и эффекторы имеют смысл только в сочетании, а эволюция чувств тесно связана с эволюцией передвижения и поведения.Способность приобретать, обрабатывать и по-разному реагировать на большие объемы информации — это также то, что отличает животных от всех других форм жизни, и история эволюции животных в значительной степени является историей эволюции сенсоров. Фактически, большинство экосистем было бы совсем другим без животных и их органов чувств. Следовательно, каждая попытка понять сенсорную эволюцию — это также попытка понять эволюцию животных и изменяющихся экосистем, частью которых они были.
В этой статье я сосредоточусь на ранних стадиях эволюции фоторецепторов, которые привели к появлению первых глаз.Существующие знания и теории из морфологии, физиологии и молекулярной биологии сформируют основу, и я исследую, как это можно сочетать с ограничениями и возможностями последовательной эволюции визуальных задач. Вот некоторые из вопросов, которые будут исследованы: какие именно сенсорные задачи привели к эволюции первых глаз; в какой последовательности развивались эти сенсорные задачи и что для этих задач требовалось с точки зрения сенсорных структур и функций. Цель статьи — расширить обсуждение ранней эволюции глаза от вопросов «как» до вопросов «почему».
(b) Молекулярные и морфологические признаки эволюции глаза
Теории происхождения и развития глаз, конечно, должны основываться на известном разнообразии современных зрительных систем и других светочувствительных систем у животных. Здесь я не буду делать подробный обзор этого огромного разнообразия, но кратко упомяну некоторые из наиболее важных аспектов (последние обзоры см. В Arendt & Wittbrodt 2001; Arendt 2003; Plachetzki et al .2005; Gregory 2008) .Самым фундаментальным и, вероятно, самым древним свойством зрительных систем является способность ощущать свет. У всех организмов есть несколько различных типов органических молекул, которые участвуют в восприятии света (Björn 2007), и они обычно связаны с определенными белками. За исключением, возможно, светочувствительности цитохром с оксидазой у личинок губок (Björn & Rasmusson 2009), светочувствительность животных основана либо на криптохромах, либо на опсинах. В то время как для невизуальной фоторецепции у животных могут использоваться криптохромы или опсины, зрение основано исключительно на опсиновых фотопигментах.Опсины животных (опсины типа II) уникальны для животных и, по-видимому, не связаны с опсинами бактерий и водорослей (опсины типа I; Larusso et al .2008).
Опсины животных принадлежат к семейству рецепторов, сопряженных с G-белками (GPCR), где подавляющее большинство белков связываются и реагируют на химические лиганды и участвуют в синаптической передаче, рецепции гормонов, обонятельности или вкусовых ощущениях (Pierce et al 2002 г.). Разница между GPCR, обнаруживающими химические стимулы, и светочувствительными опсинами заключается в том, что опсины связывают светочувствительный хромофор и сигнализируют, когда хромофор был изменен светом.В опсинах хромофор представляет собой производное витамина А, чаще всего альдегид витамина А, также известный как сетчатка, который реагирует на свет изменением конформации 11- цис на полностью транс . Таким образом, опсины по-прежнему действуют как хеморецепторные белки, но для определенного химического стимула, индуцированного светом, и, возможно, это отражает хеморецепторное происхождение животных опсинов.
Существуют разные классы опсинов, которые по-разному ведут себя при изомеризации хромофора.Ко-опсины (первоначально известные из палочек и колбочек позвоночных) высвобождают хромофор после того, как он был преобразован в изоформу all- trans (Lamb & Pugh 2004), тогда как r-опсины сохраняют и цис- 11-, и цис-. изоформы all- trans прочно связаны (r-опсины известны из фоторецепторов глаз беспозвоночных, а также представлены меланопсинами у позвоночных). В r-опсинах хромофор может быть преобразован обратно в форму 11- цис путем поглощения еще одного фотона, и эта фоторегенерация служит для пополнения чувствительного фотопигмента (Hamdorf 1977).C-opsins не могут сами регенерировать хромофор, и для этой цели требуется отдельная ферментная система (Lamb & Pugh 2004). Функционально аберрантный класс опсинов действует не как рецепторные белки, а как фотоизомеразы, которые используют свет для преобразования хромофора из формы all- trans в форму 11- cis , которая затем высвобождается и готова к включению в обычный опсин ( Сперлинг и Хаббард 1975; Смит и Голдсмит 1991; Макби и др. .2001; Гонсалес-Фернандес 2003).
Опсины из разных классов также связываются с разными G-белками, которые связаны с разными каскадами трансдукции (). Считается, что билатерии обычно обладают по крайней мере тремя классами опсинов — ц-опсинами, р-опсинами и G или -опсинами / фотоизомеразами (Terakita 2005; Raible и др. , 2006), тогда как книдарии имеют ц-опсины и уникальный class, класс книдопсов, который, по-видимому, является книжным аналогом как r-опсинов, так и G o -опсинов / фотоизомераз (Plachetzki et al .2007; Коянаги и др. . 2008; Kozmik и др. . 2008; Шуга и др. . 2008 г.). Поскольку гены опсинов отсутствуют у губок, плакозоа и хоанофлагеллят, но у губок есть другие GPCR (Suga et al .1999; Plachetzki et al .2007), похоже, что опсины произошли от ранних эуметазоанов и что разные классы частично излучали до и частично после раскола между Cnidaria и Bilateria.
Таблица 1.
Опсины и каскады трансдукции в фоторецепторах животных.ФДЭ, фосфодиэстераза; PLC, фосфолипаза C; GC, гуанилатциклаза; AC; аденилатциклаза; цГМФ, циклический гуанозинфосфат; цАМФ, циклический аденозинфосфат; IP 3 , инозитолтрифосфат; ДАГ, диацилглицерин. Для получения дополнительной информации см. Pierce et al . (2002) и Теракита (2005).
| опсин | G-белок | контролируемый фермент | второй мессенджер | полярность ответа | |||
|---|---|---|---|---|---|---|---|
| c-opsin | G t | G t | r-opsin | G q | PLC | IP 3 , DAG | деполяризация |
| G o -опсин | G o| c | GC деполяризация | | ||||
| класс книдопсов | G s | AC | цАМФ | деполяризация |
Другой общей чертой глаз животных является то, что они используют гомологичные гены развития, в частности, Pax. -6 ген (Quiring et al .1994; Гальдер и др. . 1995; Геринг и Икео 1999). Это было воспринято как доказательство того, что все глаза являются строго гомологичными органами (Gehring & Ikeo 1999; Gehring 2005), но поскольку Pax-6 и другие гены развития многократно активируются во время онтогенетического развития, неясно, была ли первоначальная роль из этих генов должен был определять сенсорную / нервную систему, фоторецепторную клетку или глаз (Nilsson 1996, 2004; Fernald 2000, 2004; Kozmik 2005). Следовательно, гены развития менее информативны в отношении эволюции глаза, чем опсины.
Почти все без исключения фоторецепторные клетки глаз животных имеют явную мембранную специализацию для размещения большого количества фотопигмента опсина, который связан с мембраной. Два наиболее распространенных типа таких специализаций мембран — это те, которые основаны на модифицированных ресничках, и те, которые основаны на микроворсинках, выступающих из тела клетки. Полученное в результате различие между цилиарными и рабдомерными фоторецепторными клетками оказало большое влияние на наше понимание эволюции глаз (Land & Fernald 1992; Arendt & Wittbrodt 2001; Nilsson 2004).Большинство глаз беспозвоночных основаны на рабдомерных рецепторных клетках, тогда как глаза позвоночных и книдарий основаны на ресничных рецепторных клетках. Различие между ресничными и рабдомерными рецепторными клетками подчеркивается тем фактом, что они строго используют разные классы опсина, c-opsin и r-opsin, соответственно, и, как следствие, они также используют разные каскады трансдукции (). Теперь ясно, что у двустворчатых животных обычно есть оба типа фоторецепторных клеток, но это зависит от типа, какой тип используется для зрения (Arendt 2003; Arendt et al .2004; Plachetzki и др. . 2005). Ситуация осложняется существованием более двух типов рецепторных клеток, промежуточных типов и клеток, которые изменяются от одного типа к другому в процессе онтогенетического развития (Eakin & Westfall 1965; Eakin 1972; Blumer 1995, 1996). Таким образом, разумным выводом является то, что различные типы фоторецепторных клеток были установлены на ранних этапах эволюции эуметазоа, но что более поздние модификации в клеточной структуре, молекулярных механизмах и использовании вместе с генетической ко-опцией в разной степени размыли картину в разных уголках мира. царство животных.
Более ранние различия между глазами с обратной или обратной сетчаткой, определяемые ориентацией рецепторных клеток (Salvini-Plawen & Mayr 1977; Arendt & Wittbrodt 2001), также имеют промежуточные или трудно интерпретируемые иным образом случаи, и в настоящее время кажется, что строение и дизайн глаз настолько различаются между типами животных, что по большей части эволюция глаз протекала независимо у разных групп животных. Для понимания ранних фаз эволюции глаза более интересно рассмотреть молекулярно определенные типы фоторецепторных клеток (Arendt 2003, 2008), а также их положение и роль у различных современных животных.В этой статье я делаю различие между невизуальной фоторецепцией и зрением. Первые включают ненейронные клетки, которые экспрессируют опсины для своего физиологического контроля, а также фоторецепторные нейроны, которые не встроены в глаза и не связаны со скрининговым пигментом или другими скрининговыми структурами. Такие клетки не будут получать никакой информации о направлении света, но будут реагировать на общую интенсивность окружающей среды. Предпосылкой для визуального фоторецепции является то, что он предоставляет информацию о направлении света либо с помощью частично экранированных фоторецепторов и сканирующих движений тела ( a — c ), либо путем сравнения сигналов от нескольких фоторецепторов в глазах (истинное пространственное зрение; ). d , и ).
Полусхематические изображения глазков и простых глаз: ( a ) одноклеточный глазок личинки коробчатой медузы; ( b ) глазок личинки полихеты; ( c ) глазок плоского червя acoel, с отражающими тромбоцитами в пигментной клетке, но без мембранных стэков в двух рецепторных клетках; ( d ) глаз с обратной чашечкой плоского червя планарии и ( e ) глаз с обратной линзой ювенильной коробчатой медузы. Фоторецепторные клетки обозначены зеленой штриховкой. pr — скрининг пигмента в рецепторной клетке; mc, подвижная ресничка; пк — специализированная пигментная клетка; rh, рабдом; r — светоотражающие кристаллы; rc, рецептивная ресничка.
(c) Типы процессов в эволюции глаза
Глаза, как и другие сложные органы, состоят из различных специализированных клеток, которые имеют клеточную морфологию и биохимические механизмы, необходимые для их конкретной функции. Очевидно, что некоторые структуры и молекулярные механизмы должны быть на месте, прежде чем появится смысл для развития других. С точки зрения эволюции глаза ясно, что светочувствительность должна развиться до некоторой степени до того, как структуры рецептор-клетка начнут развиваться, а рецепторные клетки, в свою очередь, должны существовать до того, как может быть собран многоклеточный глаз.В этой последовательности событий вступают в игру четыре различных эволюционных процесса: (i) эволюция молекулярных компонентов, (ii) эволюция клеточных структур, (iii) эволюция типов клеток и (iv) эволюция формы органов (обозначена как a). — d дюйма). Эти четыре процесса пересекаются и взаимодействуют, но каждый процесс вносит изменения по-своему.
Четыре процесса ( a , молекулярные компоненты; b , клеточные структуры; c , типы клеток; d , форма органов), участвующих в эволюции глаза.Процессы перекрываются во времени, но инициируются в порядке возрастания.
Молекулярные компоненты, такие как опсины и трансдукционные белки, подвержены изменениям в последовательности белка, введенным случайными мутациями. Отбор воздействует на изменения последовательности, и, поскольку подавляющее большинство изменений являются нейтральными или вредными, небольшой выбор, доступный для отбора, заставит направление эволюции последовательности в значительной степени зависеть от случайной природы мутаций, и процесс генерирует дискретные, а не непрерывные изменения.Гены могут играть более чем одну роль, и это совместное использование генов (Пятигорский 2007) имеет важные последствия для изменений. Классическими примерами совместного использования генов в зрении являются стрессовые белки или шапероны, которые также функционируют как кристаллины в хрусталике или роговице (Пятигорский, 1998). Чем шире генетическое совместное использование, тем больше вероятность того, что изменение последовательности имеет некоторые положительные эффекты, но также увеличивается риск отрицательных последствий (Пятигорский 2007). Лекарством от этого гордиева узла является дупликация генов, при которой разные копии могут свободно специализироваться для разных ролей.Дупликация генов без предварительного совместного использования генов, конечно, также возможна, но это менее мощный эволюционный механизм. Дублирование генов и последующая модификация ответственны за множество различных типов опсинов (Terakita 2005).
Эволюция клеточных структур, таких как рабдомы, модифицированные реснички или аксоны, необходима для определенных функций или, по крайней мере, для повышения эффективности функций. Изменения генов развития в конечном итоге ответственны за скоординированную экспрессию белков, которые образуют клеточные структуры.Постепенный и количественный характер таких модификаций обычно является результатом взаимодействий между многими генами (Futuyma 1986), и поэтому можно ожидать, что случайность мутаций будет иметь лишь незначительное влияние на эволюцию этих характеристик.
Эволюция типов клеток, которую чаще всего можно описать как клеточную специализацию, обычно требует растущего числа отдельных типов клеток. Дупликация клеточного типа с последующей сегрегацией считается центральным механизмом в эволюции сложности многоклеточных животных (Arendt 2008; Arendt et al. 2009). Генетическая основа — это подспецификация типов клеток посредством модификаций генов, формирующих паттерн, с последующим более специализированным профилем экспрессии каждого нового типа клеток. Считается, что многие специализированные клетки сетчатки глаза животных произошли от предковых клеток с множеством функций, подобно тому, как они превратились из оркестра одного человека в большой оркестр. Считается, что такие принципы ответственны за эволюцию различных типов рецепторных клеток, пигментных клеток и интернейронов из предковых рецепторных клеток (Plachetzki & Oakley 2007; Arendt 2008; Erclik et al .2008 г.). Несколько иной способ дивергенции клеточного типа очевиден в сетчатке многих животных и приводит к постепенному изменению морфологии клеток от одной части сетчатки к другой. Яркий пример — хрусталик глаз коробчатой медузы, где все ресничные рецепторные клетки содержат темный экранирующий пигмент. По направлению к периферии сетчатки сегменты ресничек становятся меньше, и происходит постепенный переход от пигментированных рецепторных клеток к чистым пигментным клеткам, полностью лишенным ресничек, которые образуют радужную оболочку глаза (O’Connor et al .2009 г.).
Форма и размер глаза имеют решающее значение для его оптических характеристик. Параметры конструкции органов контролируются пролиферацией и ростом клеток, и по аналогии с эволюцией сложных клеточных структур, многие различные гены, вероятно, будут вовлечены в морфологию органов. Это делает изменение внутри популяции практически непрерывным (Футуйма, 1986). Выбор из непрерывного (не дискретного) изменения является причиной того, почему глаза могут быть так точно настроены для работы, близкой к физическим пределам (Nilsson & Pelger 1994; Land & Nilsson 2002).
Plachetzki & Oakley (2007) подчеркивают «древовидное мышление» с дублированием и дивергенцией как дополнение к традиционным линейным моделям эволюции глаза. Как должно быть очевидно из рассуждений, упомянутых ранее, обе концептуальные основы необходимы для понимания эволюции глаза. Процессы ( a ) и ( c ) следуют концепции древовидного мышления, тогда как процессы ( b ) и ( d ) следуют линейной модели эволюции. Как я буду доказывать в следующем разделе, эволюция сенсорных систем — это всегда прерывистый процесс, в котором поведенческие задачи определяют сущности, на которые в конечном итоге воздействует отбор.
Большая часть литературы по эволюции глаза посвящена вопросам гомологии (Arendt 2003, 2008; Gehring 2005; Plachetzki et al . 2005; Plachetzki & Oakley 2007; Gregory 2008). Основная причина того, почему гомология является такой сложной концепцией в эволюции глаз, состоит в том, что генетические сети развития, вероятно, были кооптированы клетками в новых местах (Nilsson 2004; Vopalensky & Kozmik 2009). Истинная гомология требует, чтобы признак, разделяемый разными видами, имел непрерывную историю, восходящую к одному и тому же признаку у общего предка.Это означает, что гомологичная структура должна развиваться из клеток в одном и том же онтогенетическом составе клеток или из соответствующих линий в других сегментах тела (последовательная гомология). Структуры, которые развиваются из клеток с разными родословными клеточного деления, не гомологичны, но вполне могут быть кооптированы гомологичными контрольными генами (True & Carroll 2002; McLennan 2008). Эктопические глаза были экспериментально индуцированы в различных положениях из-за неправильной экспрессии генов развития как у Drosophila , так и у Xenopus (Halder et al .1995; Onuma и др. . 2002). Внематочные глаза — это искусственно вызванные случаи кооптации, и количество различных генов развития, способных вызывать эктопические глаза, указывает на то, что кооптации часто встречаются в эволюции.
Ко-вариант в соседних клетках с похожими, но не идентичными родословными, вероятно, будет легче индуцировать, и такой ко-вариант также будет особенно трудно отличить от гомологии. Кроме того, экспрессия генов, специфичных для определенного типа клеток, будет вводить в заблуждение в тех случаях, когда целые наборы генов кооптируются как сборки, и то, что кажется множеством независимых сходств, на самом деле является только одним сходством с определенной вероятностью ложного указания. гомология.Проблемы еще больше усугубляются модульной природой генетических сетей развития (Oakley 2003). Совместное использование таких компонентов, как опсины, G-белки и ионные каналы из разных систем, может создавать новые системы с бриколажем гомологий генов развития. Взгляд на край мантии моллюсков и на жаберные щупальца веерных червей, вероятно, является результатом обширной кооптации и генетического бриколажа (Nilsson 1994; Oakley 2003). Коопцию и бриколаж следует понимать не как исключения, а как стандартные эволюционные механизмы, которые в значительной степени способствуют эволюционируемости (Lenski et al .2006 г.).
На основе существующих морфологических и молекулярных данных, а также обсуждения эволюционных процессов, упомянутых ранее, можно подвести некоторые итоги по основным ветвям эволюции глаза. Кажется очевидным, что светочувствительность на основе опсина имеет единственное происхождение и была продублирована во множестве систем на очень раннем этапе эволюции животных (Plachetzki et al .2007). Парные головные глаза беспозвоночных обнаруживают многие признаки гомологии, такие как r-opsins, каскад трансдукции G q и рецепторы с рабдомерной морфологией (Arendt 2003, 2008).Отдельные личиночные и взрослые головные глаза у многих типов беспозвоночных (Arendt & Wittbrodt 2001) можно рассматривать как случай последовательной гомологии, но какой из них является исходным глазом, остается открытым вопросом, ожидающим более глубоких знаний об историях жизни обычных животных. предки. Глаза позвоночных, вероятно, лучше всего понимать как бриколаж компонентов (Arendt et al .2004; Arendt 2008), возможно потому, что ранние позвоночные прошли фазу, в которой предковые системы были упрощены, прежде чем они снова стали сложными (см. Также Lamb et al. .2007). Глаза без головного мозга, как правило, могут быть результатом кооптации контрольных генов или набора и развития невизуальных фоторецептивных систем. Глаза книдарий обнаруживают связи с разными классами фоторецепторов у билатерий, но еще слишком рано, чтобы быть уверенным в уровне гомологии между глазами у этих двух основных ветвей животных. Затем я исследую, могут ли селективные преимущества, которыми руководствовалась ранняя эволюция глаза, помочь рассеять туман над некоторыми из проблем, обсужденных ранее.
2. Ранние сенсорные задачи и их функциональные потребности
Широкое распространение глаз или глазков позволяет сделать два связанных вывода: во-первых, глаза или глазки предлагают значительное функциональное преимущество для подавляющего большинства образов жизни животных, и во-вторых, что глаза или ocelli появились очень рано в эволюции животных. Это одна общая роль или много разных ролей? Чтобы ответить на этот вопрос, необходимо определить типы информации, которую потенциально может предоставить свет.Общая интенсивность окружающего освещения зависит от времени суток, времени года, лунного цикла, погоды, глубины воды и положения относительно затеняющих структур. Разумно предположить, что эволюция сенсорных задач, основанных на светочувствительности, началась как ненаправленный мониторинг внешней освещенности, и что это легло в основу, на которой развились направленная чувствительность и истинное зрение. Ненаправленный мониторинг интенсивности света может использоваться в качестве увлечения для суточных часов; он может выполнять задачи фотокинеза, такие как реакция тени или контроль глубины в воде, или он может выступать в качестве индикатора поверхности роющего образа жизни.Как только рецепторная клетка приобретает некоторую направленность, она может управлять прямым фототаксисом и действовать как оптическая статоциста для управления ориентацией тела. Множественные фоторецепторные клетки, направленные в разные стороны, являются основой истинного зрения, и это позволяет выполнять огромное количество зрительных задач, выполняемых членистоногими, головоногими и позвоночными. Краткое изложение наиболее примитивных ролей, которые играют световые рецепторы животных, дается ниже. Список более сложных визуальных задач см. В Land & Nilsson (2006).
Таблица 2.
Свойства фоторецепторных систем и возможные сенсорные задачи. Обнаружение теней не включено, потому что, хотя оно не требует направленности, оно требует быстрого отклика и адаптации.
| свойства сенсорной системы | сенсорные задачи |
|---|---|
| ненаправленная светочувствительность: нет экранирующего пигмента, медленная реакция, большой динамический диапазон, без адаптации | циркадный захват |
| УФ предупреждение | |
| обнаружение поверхности: роющий образ жизни | |
| чувствительность к направленному свету: экранирующий пигмент, умеренная скорость отклика, ограниченный динамический диапазон, адаптация | фототаксис |
| оптическая статоциста | |
| рецепторы, пигментная чашка, низкое разрешение, умеренная скорость реакции, ограниченный динамический диапазон, адаптация | навигация по отношению к неодушевленным структурам |
| пространственное видение: множественные рецепторы, фокусирующая оптика, высокое разрешение, быстрая реакция, ограниченный динамический диапазон, адаптация | интерактив вместе с другими животными |
Для каждой из примитивных ролей светочувствительности животных существуют функциональные требования с точки зрения абсолютной чувствительности, спектральной чувствительности, скорости и адаптации к интенсивности.Эти требования имеют здесь центральное значение, потому что они должны были вызвать отбор, управляющий эволюцией клеточных структур и молекулярных механизмов в фоторецепторных и поддерживающих клетках. Ниже приводится анализ этих требований, начиная с основных требований к светочувствительности и прорабатывая сенсорные задачи, основанные на уровнях внешней освещенности, посредством направленного приема света и, наконец, до истинного (пространственного) зрения.
(a) Светочувствительность
Обнаружение света опсинами животных означает, что потребляется хромофор 11- цис .Чтобы поддерживать чувствительность, активные хромофоры должны пополняться со скоростью, сопоставимой с потреблением. Прямой солнечный свет соответствует 10 миллионам фотонов видимого света в секунду, достигающим каждого квадратного микрометра земной поверхности, и это потребляет хромофор 11- цис с экстремальной скоростью в облученных фоторецепторных клетках. Однако спонтанная (термическая) регенерация от полностью транс к сетчатке 11 цис происходит только с очень низкой скоростью (Deng et al .1991). Использование света также для обратного переключения хромофора из конфигурации полностью транс в конфигурацию 11- цис решает проблему, но для свободного хромофора это требует ультрафиолетового (УФ) излучения, которое отсутствует даже на умеренных глубинах моря. Фотоизомеразы, которые не являются сенсорными опсинами, обладают свойством регенерировать хромофор в присутствии видимого света, и, как таковые, они комплементарны к-опсинам, которые лишены этой способности. Р-опсины отличаются тем, что они действуют как рецепторные белки, так и фотоизомеразы, так что хромофор может оставаться связанным с опсином все время.Если предположить, что сложный цикл фотопигментации позвоночных с ферментативной темновой регенерацией (Lamb & Pugh 2004; Kusakabe et al. 2009) является более поздним приобретением, то функциональная эволюция зрительных пигментов может быть предварительно реконструирована.
Древний опсин, из которого произошли к-опсины, р-опсины и фотоизомеразы / G o -опсины, мог быть чистым рецепторным белком (например, к-опсином), чистой фотоизомеразой или белком с комбинированными свойствами. (как р-опсин).Из этих трех возможностей предковый опсин вряд ли был фотоизомеразой, потому что трудно увидеть потребность в таком ферменте, если нет рецепторного белка, продуцирующего его субстрат. Происхождение в виде комбинированного (бистабильного) белка приведет к двум альтернативным сценариям, в которых фотоизомераза или чистый рецепторный белок могут развиться из одной из копий после дупликации гена. Последний вариант маловероятен, потому что чистый рецепторный белок (тип c-opsin) будет уступать исходному бистабильному белку до того, как появится фотоизомераза.Если, с другой стороны, фотоизомераза произошла из копии гена исходного бистабильного пигмента, то класс c-opsin эволюционировал бы последним и образовал кладу с r-опсинами или G o -опсинами / фотоизомеразами, и ни одна из этих филогений не поддерживается (Plachetzki et al , 2007;). Таким образом, единственный жизнеспособный вариант состоит в том, что предковый опсин был чистой рецепторной молекулой, подобной современным к-опсинам. Если на втором месте будет бистабильный пигмент, это, вероятно, приведет к потере исходного гена, потому что чистый рецепторный белок без дополнения фотоизомеразы будет значительно хуже бистабильного опсина.Единственный оставшийся сценарий состоит в том, что предковый опсин обладал свойствами к-опсина и что первая дупликация гена привела к эволюции фотоизомеразы, которая вместе с предковым опсином сформировала эффективную функциональную пару. Класс р-опсинов затем развился бы после дупликации и модификации фотоизомеразы, и подобное событие произошло бы снова с образованием G o -опсинов. Такой сценарий согласуется как с филогенией опсинов (Plachetzki et al .2007) и с функциональными аргументами ().
Гипотетическая эволюция функции опсина. Основные аргументы см. В тексте. Филогенетическое древо составлено согласно Plachetzki et al . (2007).
Первоначальное расщепление для генерации фотоизомеразы вместе с рецепторным белком должно было произойти очень рано, до расщепления на книдарий и билатерий. Если современные книдарии используют к-опсины в своих глазах, как было предложено Kozmik et al . (2008), можно ожидать, что клады книдопсов содержат фотоизомеразы, которые работают вместе с цопсинами.Наблюдение за обесцвечиванием опсинов медуз (O’Connor et al , 2009) приводит к гипотезе о том, что фоторецепция книдарий основана на отдельных рецепторных и ферментных белках.
Связанный с этим вопрос заключается в том, почему опсины стали единственным типом зрительного фотопигмента, в то время как животные используют как опсины, так и криптохромы для невизуальной фоторецепции. Это примечательно, потому что криптохромы были унаследованы от одноклеточных предшественников животных (Rubin et al .2006), но опсины типа II уникальны для животных (Larusso et al , 2008) и, следовательно, должны были развиться от предка, который уже обладал криптохромами. Спонтанная регенерация криптохромов происходит с разумной скоростью (Kennis et al .2004; Bouly et al .2007; Losi 2007; Kao et al .2008), и в этом смысле криптохромы были бы лучше, чем древний отбеливающийся опсин. Таким образом, вероятно, есть и другие причины, которые сделали опсины жизнеспособной альтернативой криптохромам.Возможно, что скорость и усиление сигнального пути дали опсинам преимущество для зрительных систем, в которых скорость и чувствительность более важны, чем для невизуальных ролей. Альтернативное объяснение заключается в том, что спектральная чувствительность опсинов больше подходит для зрения в зеленом свете, который лучше всего проникает в прибрежную воду (500–550 нм), тогда как синяя и УФ-чувствительность криптохромов (менее 500 нм) адекватна для не- визуальные задачи (Геринг, Росбаш, 2003).
(b) Задачи рассеянного света
Яркость, создаваемая естественными источниками света, варьируется на впечатляющие 8 логарифмических единиц между солнечным светом и звездным светом (Land & Nilsson 2002).Сильно пасмурное небо снизит яркость примерно на логарифмическую единицу, в результате чего средняя яркость сцены изменится примерно на 9 логарифмических единиц с солнечного дня на пасмурную безлунную ночь (). Разница в яркости между различными частями сцены намного меньше, обычно 1,5–2 логарифмических единицы (без источника света). Глубина в воде сильно влияет на силу света. Даже самая чистая океанская вода ослабляет свет примерно на 1,5 логарифмических единиц на каждые 100 м, а в прибрежной воде ослабление во много раз выше.
Естественное сияние и чувствительность глаз. ( a ) Часть цикла дневного света, нанесенная на логарифмическую шкалу интенсивности. Диапазон яркости внутри сцены постоянен, но скользит вверх и вниз по шкале журнала между днем и ночью. В пасмурных условиях вся функция яркости смещается вниз максимум на 1 логарифмическую единицу. ( b ) Рабочий диапазон фоторецепторов был смоделирован для ненаправленного мониторинга окружающей яркости, направленного сканирования фототаксиса и пространственного видения (расчеты и данные см. В дополнительных электронных материалах). Светло-зеленый указывает рабочий диапазон для фоторецептора без расширений мембраны, а темно-зеленый — для соответствующего фоторецептора с реалистичным количеством мембран, наложенных друг на друга. Ненаправленный фоторецептор работает до уровней ниже звездного без какой-либо наложения мембран, но визуализирующие глаза не могут различать яркость в пределах сцены даже при ярком солнечном свете, если мембрана фоторецептора не сложена сильно. Комбинируя большие линзы, среднее разрешение и слабое зрение, ночные и глубоководные животные могут использовать свои глаза при более низкой яркости, чем указано в крайней правой зеленой полосе (Warrant 1999).( c ) Глубина чистой океанской воды нанесена на ту же шкалу яркости, предполагая, что солнечный свет у поверхности.
24-часовой световой цикл имеет очевидное отношение к большинству животных. Даже первые многоклеточные животные выиграли бы, если бы узнали, когда их пищевые организмы участвуют в фотосинтезе. Современные планктонные организмы, как животные, так и их пищевые организмы, участвуют в суточных вертикальных миграциях, чтобы найти оптимальные условия для питания / фотосинтеза, избежать поедания, доступа к питательным веществам, расселения и ряда других факторов (Harris 1953; Putzeys & Hernández- Леон 2005).Многие из этих факторов могли бы стимулировать вертикальные миграции также во времена первых многоклеточных животных, до того как появились хищники, управляемые визуально, и макроскопические животные. Ненаправленный мониторинг интенсивности света достаточен для управления вертикальным положением водных организмов, но для этого требуется сочетание фотокинеза и ориентации по гравитации (пассивное или посредством геотаксиса; Häder et al. .2005; Roberts 2006; Richter et al. .2007). Свет является не менее важным сигналом для эпибентосных или частично роющих животных, а циркадные часы и оптические глубиномеры, содержащие свет, могли бы предложить селективные преимущества для урметазоа или урбилатерийских предков, независимо от того, плавали они или ползали.
Использование света в качестве меры глубины в воде осложняется тем фактом, что яркость определяется как временем дня, так и глубиной. Если организм строго не следует изолямам, необходимо разделять причины любых изменений яркости, чтобы можно было распутать время суток и глубину в воде. Есть два очевидных способа для вертикально мигрирующего планктона решить неоднозначность яркости. Во-первых, циркадный ритм может предвосхищать время суток, и установка часов может выполняться на протяжении многих световых циклов, так что влияние колебаний глубины может быть минимизировано.Другая возможность — использовать тот факт, что спектральный состав света меняется по-разному в зависимости от времени и глубины (Lythgoe 1979). Последнее оказывает гораздо более сильное влияние на спектральный состав: чистая океаническая вода с глубиной постепенно становится голубее, а прибрежная вода — зеленее. Два спектрально разных фотопигмента могут образовывать очень надежный глубиномер, который в значительной степени не зависит от цикла диэлектрического света. Еще одна причина эволюции более чем одного фотопигмента заключается в том, что для высокой чувствительности требуется спектральный пик, близкий к максимальному пропусканию воды, тогда как обнаружение УФ-света может быть полезно для предотвращения радиационного повреждения белков и генетического материала, которое может происходить вблизи поверхность в течение дня.
Таким образом, у ранних многоклеточных животных были бы веские причины приобрести световосстановленные циркадные часы и спектрально разные фотопигменты для измерения глубины воды и предупреждения о воздействии УФ-излучения. Во всех этих задачах информацию несут абсолютные интенсивности. Это означает, что рецепторы должны охватывать очень большой динамический диапазон и что адаптация нежелательна. Изменения интенсивности также медленные, что требует медленных реакций, интегрирующихся в течение длительного времени.Это типичные свойства невизуальных фоторецепторов (Gotow & Nishi 2008). Для предупреждения об УФ-излучении фоторецепторы могут иметь меньший динамический диапазон и более быструю реакцию, но для этой задачи также должна отсутствовать адаптация.
Глубиномер, основанный на сравнении сигналов от двух спектрально разных опсинов, мог быть ранней причиной не только спектрально различных опсинов, но также хроматического антагонизма и различных каскадов трансдукции. Рецепторы в теменном глазу ящериц, хотя и не используются в качестве измерителя глубины, обладают механизмом хроматического антагонизма в отдельной клетке (Solessio & Engbretson, 1993), и два опсина действуют антагонистически через разные каскады трансдукции с противоположными эффектами на ионные каналы. (Су и др. .2006 г.). У ранних многоклеточных животных эволюция сходных хроматических механизмов могла приводить к расхождению как в опсинах, так и в механизмах трансдукции, а позже они были разделены на разные типы клеток.
(c) Задачи направленности
Направление света содержит огромное количество информации об окружающем мире. Первым эволюционным шагом для начала изучения этой информации является получение некоторой формы частичного экранирования фоторецепторных клеток. Чувствительность к направленному свету могла возникнуть как побочный продукт защитной пигментации тела.Несмотря на то, что скрининговый пигмент является типичной частью глаз и глазков, потребность в пигментации не возникла с развитием зрения. Пигменты присутствуют во всех организмах, которые подвергаются воздействию света, и они защищают клетки от повреждения коротковолновым светом. Причина, по которой молекулы биологических пигментов, таких как меланины и птерины, могут поглощать свет, не повреждая себя, заключается в том, что сильно сопряженные двойные связи позволяют энергии распределяться по молекуле и легко рассеиваться в виде тепла.По той же причине биологические пигменты удаляют вредные свободные радикалы и защищают клетку от окислительного повреждения (McGraw 2005). Коротковолновый свет является одной из причин появления свободных радикалов, и поэтому биологические пигменты играют двойную роль в защите от неблагоприятного воздействия света. Это означает, что скрининговые пигменты должны быть обычными клеточными составляющими задолго до того, как у животных появилось зрение.
Если пигментация тела обеспечивает некоторую рудиментарную направленность фоторецепторной клетки, легко увидеть, как отбор может уловить ее и улучшить направленность путем синтеза скринингового пигмента в рецепторной клетке или в соседних клетках.Как только появляется некоторая направленность ( a — c ), собственные движения животного позволяют сравнивать интенсивность света в разных направлениях. Эта информация может использоваться для фототактической ориентации к свету или от него (обсуждение фототаксиса у планктонных организмов см. Jékely 2009) или может функционировать как оптическая статоциста для управления положением тела по отношению к общему направлению света. Переход от невизуальных задач к фототаксису требует серьезных изменений в свойствах рецепторных клеток.Актуальной информацией являются уже не абсолютные интенсивности, изменяющиеся на 8 логарифмических единиц между днем и ночью, а быстрые изменения (секунды), охватывающие 1,5–2 логарифмических единицы, наложенные на 24-часовой световой цикл. Чтобы извлечь информацию для фототаксиса, рецепторным клеткам требуется быстрый отклик, соответствующий образцу движения животного, и высокая контрастная чувствительность для обнаружения сравнительно небольших различий яркости в разных направлениях. Также необходим механизм адаптации, чтобы исключить 24-часовой световой цикл из ответа.
Эти свойства требуют как количественных, так и качественных модификаций механизмов обнаружения и передачи сигналов в рецепторных клетках; но, что более важно, уменьшенный угол, под которым принимается свет, вместе с гораздо более коротким временем интегрирования приводит к резкому сокращению количества фотонов, обнаруживаемых в течение каждого времени интегрирования. Необходимость различать меньшие различия в интенсивности одновременно требует больших выборок фотонов (Land & Nilsson 2002).Чтобы оценить способность различать соответствующие различия в интенсивности, я рассчитал необходимую минимальную интенсивность для выполнения конкретных задач, таких как определение ритма интенсивности, простой фототаксис или истинное пространственное (изображение) видение (подробности см. В дополнительных электронных материалах). Расчеты были основаны на пяти реалистичных моделях: одной из невизуальных фоторецепторов, двух глазков с разным направлением, используемых для фототаксиса, и двух глаз с пространственным разрешением (глаз без линзы и маленький глаз типа камеры).Центральный вопрос, побудивший к моделированию, заключался в том, необходима ли укладка мембран, таких как микроворсинки, ламели или диски, для различных задач.
Результаты оказались неожиданными (см. Дополнительные электронные материалы). Ненаправленный фоторецептор для определения цикла диэфирного света может обеспечивать полный диапазон яркости в 8 логарифмов без каких-либо морфологических специализаций мембран. Даже если плотность фотопигмента на несколько логарифмических единиц ниже, чем в зрительных клетках позвоночных или насекомых, чувствительность будет достаточно высокой для обнаружения лунного света.Таким образом, для невизуального мониторинга яркости нет необходимости в ресничках или микроворсинках. Действительно, невизуальные фоторецепторы часто вообще лишены этих заметных структур (Gooley et al . 2003; Gotow & Nishi 2007, 2008; Van Gelder 2008). Моделирование также демонстрирует, что фототаксис, который требует направленности с большой угловой чувствительностью и промежуточным временем интегрирования, может функционировать без наложения мембран, но только в течение дня и на довольно мелкой воде (см. Таблицу S1, дополнительный электронный материал).Тот факт, что у плоских червей acoel есть глазки без ресничек или рабдомеров, подтверждает вывод, что направленная фоторецепция (предположительно для фототаксиса) не требует укладки мембран (Yamasu 1991). Для использования при низкой сумеречной интенсивности в более глубокой воде или в очень мутной воде умеренное наложение мембран поможет расширить диапазон интенсивностей, в которых можно использовать фототаксис ().
Направленность, быстрый отклик и адаптация, действующие как временный фильтр верхних частот, являются важными свойствами фоторецептора, который служит фототаксису или функционирует как оптическая статоциста ().Эти свойства передают информацию об угловом распределении света в окружающей среде и удаляют информацию об общей яркости окружающей среды. Поэтому фоторецепторная ячейка, которая играет роль в измерениях направленного света, должна отказаться от любой первоначальной задачи по мониторингу общей окружающей яркости. Но поскольку увлечение биологических часов, вероятно, останется важным, направленная фоторецепция, вероятно, будет развиваться только после дублирования ненаправленного фоторецептора.Эволюция фототаксиса и оптических статоцист, таким образом, вероятно, послужила ранними причинами возникновения множественных фоторецепторных систем у ранних многоклеточных животных.
Ключевые инновации в развитии глаз. Предполагается, что направленная фоторецепция возникла в результате ненаправленного мониторинга окружающей яркости за счет дублирования клеток и дупликации гена опсина, приводящей к парам белков рецептора опсина и фотоизомеразы для эффективной регенерации хромофора. За этим последовало введение скринингового пигмента, а вскоре и укладка мембран, чтобы обеспечить лучшее различение контраста, увеличенную скорость и более направленную фоторецепцию.Множественные рецепторные клетки тогда позволили бы истинное пространственное зрение, и можно было бы отказаться от режима сканирования. Однокамерные и сложные глаза должны развиваться независимо от направленных глазков. Чтобы собрать достаточно фотонов для пространственного зрения с более высоким разрешением, необходимо ввести линзы, но ожидается, что новые задачи с высоким разрешением дополнят, а не заменят старые задачи с низким разрешением.
(d) Задачи визуализации
Однонаправленный фоторецептор полагается на движение тела для получения информации об угловом (пространственном) распределении света, и это мешает способности обнаруживать изменения в окружающей среде.Когда разные фоторецепторы направлены в разные стороны, пространственная информация может собираться одновременно без движения тела. Когда всего два рецептора направлены в разные стороны, в принципе есть изображение, а добавляя больше рецепторов, глаз может увеличивать объем информации практически без ограничений. Пространственное зрение также является наиболее информативным из всех органов чувств, и оно предлагает сенсорное руководство для самых изощренных форм поведения животных. Не может быть никаких сомнений в том, что шаг от направленного фоторецептора к первой ямке или чашечке глаза имел решающее значение для эволюции животных.
Развитие глаза с пространственным разрешением из направленного фоторецептора означает, что угол, видимый каждым рецептором, должен будет уменьшиться, и это резко снижает скорость обнаружения фотонов. Переход от поля 180 ° личиночного глаза Platynereis dumerilii ( b ; Jékely и др. , 2008) к полю 25 ° в маленьком глазе с пигментной чашкой ( d ) включает 42 раза снижение скорости регистрации фотонов. Эта потеря чувствительности — главное препятствие в развитии пространственного зрения.Проблема усугубляется тем фактом, что время интегрирования должно быть сокращено вместе с полем обзора рецептора, чтобы поддерживать размытие при движении на допустимом уровне. Кроме того, интересующие нас контрасты меньше для пространственного зрения, чем для фототаксиса, который требует больших выборок фотонов за время интегрирования (см. Дополнительные электронные материалы для обсуждения размера фотонных выборок). Следствием этой быстро растущей потребности в фотонах является то, что наложение фоторецепторной мембраны становится абсолютной предпосылкой для эволюции пространственного зрения.Даже дневной свет был бы слишком тусклым для глаза с пигментной чашкой без какой-либо укладки светочувствительной мембраны (; таблица S1, дополнительный электронный материал). Таким образом, рабдомы и ресничные специализации должны были существовать, по крайней мере до некоторой степени, до того, как появились первые пространственно разрешающие глаза.
Поскольку каждый двойной слой мембраны может поглощать не более 0,1% света (см. Дополнительные материалы по электронике), существует большой потенциал при складывании и укладке мембраны.Но когда мембранных слоев становится много, прирост, создаваемый каждым новым слоем, уменьшается. У человеческого жезла около 1300 дисков, а у рабдома мясной мухи примерно в два раза больше микровиллярных слоев. По сравнению с развернутой клеточной мембраной такое наложение включает увеличение чувствительности на 2–3 логарифмических единицы (см. Уравнения в дополнительном электронном материале). Если бы такое усиление чувствительности было потрачено на компенсацию потери чувствительности, вызванной уменьшением угловой чувствительности, сколько пространственного разрешения можно было бы купить? Ответ несколько неутешительный.Начиная с поля восприятия 180 ° системы направленного фототаксиса, 2–3 логарифмических единицы дополнительной чувствительности, получаемой при наложении мембран, достаточно только для уменьшения угла примерно до 10 °, и если время интегрирования меньше, а образцы фотонов больше учитываются, разрешение улучшается еще меньше. Угловая чувствительность 10–20 ° достаточно мала для навигации по неодушевленным структурам окружающего мира, но она должна быть значительно уменьшена, чтобы разрешить более сложные визуальные задачи, такие как преследование добычи, обнаружение хищников или распознавание партнера.
После исчерпания преимуществ, связанных с наложением мембран, очевидно, что необходима другая стратегия для постоянного улучшения визуального разрешения. Эта стратегия — введение фокусирующей оптики. Путем введения линзы область обнаружения света может быть перемещена из области рецепторной клетки в область линзы (таблица S1, дополнительный электронный материал). Поскольку клетки животных имеют диаметр всего 5–10 мкм, а линзы животных могут иметь диаметр в диапазоне миллиметров или даже сантиметров, область обнаружения может быть увеличена до 8 логарифмических единиц.Это позволяет значительно увеличить улавливание фотонов, что необходимо для высокого разрешения глаз членистоногих, головоногих и позвоночных, и обеспечивает достаточную чувствительность, чтобы настроить дизайн глаза для ночного или глубоководного использования. Для обсуждения более сложных визуальных задач см. Land & Nilsson (2006).
Линзы можно вводить постепенно, если есть ткань, заполняющая полость глазного пигментного колпачка ( e ). Путем экспрессии подходящих белков показатель преломления может быть увеличен и распределен таким образом, что формируется высококачественная линза (Nilsson & Pelger 1994).Недавняя работа с глазами коробчатой медузы (O’Connor et al , 2009) предполагает, что клетки, заполняющие полость везикулярных глаз, могут выполнять несколько функций и что фокусировка не является основной функцией. До того, как изменятся свойства фокусировки, очевидный способ сформировать изображение — это затенение в пигментной ямке или чашке. Простой способ сформировать чашку для пигмента — заставить рецепторные клетки выровнять внешнюю поверхность растущего комка прозрачных клеток. Такая механическая функция потребовала бы некоторой жесткости прозрачных ячеек, и это, естественно, предварительно адаптировало бы глазок чашки для перехода в глазок хрусталика.Другая и, возможно, даже более старая функция клеток стекловидного тела в полости глазных ямок или чашечек — служить УФ-фильтрами для защиты от радиационного повреждения рецепторных клеток. Таким образом, кажется, что эволюция хрусталика не требует каких-либо необычных обстоятельств, и причина, по которой у многих беспозвоночных вместо хрусталика в глазах, скорее всего, является то, что отбор не способствует никакому визуально управляемому поведению, требующему высокого разрешения в этих разновидность.
Пространственное зрение может возникать не только путем умножения рецепторных клеток внутри пигментной ямки, но не менее жизнеспособным вариантом является умножение всей структуры, включая пигментную ямку.У плоских червей представлены обе эти возможности (Kuchiiwa et al , 1991). На ранних этапах эволюции глаза нелегко увидеть, что одна из двух альтернатив была бы намного лучше другой. Однако позже, когда появились линзы, оказалось, что однокамерный раствор значительно превосходит многослойный глаз (Kirschfeld 1974). Одна из причин такой разницы заключается в том, что в глазах сравнимого размера множество линз сложного глаза должно быть намного меньше, чем одиночная линза в глазу камерного типа.
Диапазон яркости приблизительно 1,7 логарифма в естественных сценах будет скользить вверх и вниз по шкале интенсивности между днем и ночью, при изменении облаков и погоды, а также когда животное меняет глубину в воде или перемещается между затененными и открытыми участками. Чтобы обнаружить минимально возможный контраст в пределах диапазона яркости сцены, необходимо для визуализирующих глаз адаптироваться, чтобы функция отклика / интенсивности всегда соответствовала диапазону яркости (Лафлин, 1981). Это означает, что информация об абсолютной интенсивности удаляется из зрительной системы.Тем не менее, некоторые механизмы адаптации, такие как диаметр зрачка и величина нейронного суммирования (Warrant 1999), в идеале должны контролироваться абсолютной интенсивностью. Как следствие, невизуальная фоторецепция выполняет функцию визуализации глаз, и неудивительно, что зрительные интернейроны, такие как некоторые ганглиозные клетки в глазах позвоночных, по своей природе чувствительны к свету, не внося прямого вклада в зрение (Gooley et al . 2003 г.). Эти клетки демонстрируют сходство с другими невизуальными фоторецепторами в медленном ходе реакции (Do et al .2009), но похоже, что они не полностью лишены адаптации (Wong et al . 2005). Для эволюционного понимания внутренних светочувствительных интернейронов в зрительном пути важно отметить, что эта невизуальная функция, вероятно, представляет развитие зрительного пути, и, как таковая, она не обязательно является сохраняющейся функцией датирования. назад к довизуальным предкам.
3. Синтез и мировоззрение
Движущей силой сенсорной эволюции является добавление новых сенсорных задач, которые дают животным новые реакции на внешнюю информацию.Это рассуждение приводит к пониманию сенсорной эволюции как процесса с разделением задач. Для развития каждой новой задачи сенсорная система будет подвергаться отбору, направленному на извлечение определенного подмножества сенсорной информации. Новые сенсорные задачи развиваются путем модификации или доработки систем, которые обслуживают уже существующие сенсорные задачи. В некоторых случаях новая задача служит той же цели, что и более старая задача, что приводит к замене и потере старой задачи. Примером замены является переход от направленных фоторецепторов, используемых для фототаксиса, к пространственному зрению с несколькими фоторецепторами.Здесь вполне вероятно, что древний сканирующий режим сбора данных был просто заменен одновременным получением пространственной информации. В других случаях добавлялась новая задача без устаревания предыдущей. Эволюция направленного фоторецептора для фототаксиса из ненаправленного мониторинга цикла диэлектрического света является очевидным примером, в котором новая сенсорная задача не нарушает ценности старой задачи. В таких случаях событие дублирования должно было инициировать развитие новой задачи.
Исходя из предположения, что эволюция в целом будет развиваться от задач с небольшими требованиями к молекулярным машинам и морфологическим структурам к задачам с постепенно более широкими требованиями, эволюционная последовательность ранних задач, ведущих к истинному зрению, может быть реконструирована с некоторой уверенностью. Эта последовательность начинается с невизуальной фоторецепции для контроля циркадных ритмов или контроля глубины воды, за которой следует направленная фоторецепция для фототаксиса или ориентации тела, которая затем заменяется истинным пространственным зрением.С точки зрения структур, это соответствовало бы последовательности от фоторецепторных клеток без мембранной специализации через направленное затенение путем скрининга пигментных структур либо в самом фоторецепторе, либо в ассоциированной клетке, вплоть до развития сворачивания мембраны, которое могло бы открыться. для достаточной чувствительности, чтобы развить первые настоящие глаза с пространственным зрением ().
События дублирования, которые привели к различным классам опсинов, вероятно, будут соответствовать введению новых или модифицированных сенсорных задач.Как утверждается в этой статье, первоначальное разделение между c-opsins и всеми другими опсинами могло привести к образованию фермента фотоизомеразы для повышения эффективности исходного опсина. Если бы это было так, дупликация гена могла бы привести к улучшению функции существующей сенсорной задачи, а не к дублированию в новую задачу и старую задачу. Поскольку эта дупликация опсина предшествовала расколу между Cnidaria и Bilateria (Plachetzki et al , 2007), сенсорная задача, требующая более эффективной регенерации, существовала бы у последнего общего предка двух групп животных.Трудно понять, что быстрая регенерация была бы решающим свойством невизуального мониторинга яркости, тем более что достаточно эффективные системы на основе криптохрома, вероятно, уже существовали (Рубин и др. , 2006). Вполне возможно, что быстрая регенерация фотопигмента способствовала развитию направленной фоторецепции.
Ожидается, что эволюция укладки мембран, будь то реснички, микроворсинки или дивертикулы, стала предпосылкой для перехода от направленной фоторецепции к первым настоящим глазам.Как следствие, если можно было бы продемонстрировать, что укладка на основе микроворсинок и ресничек имеет независимое происхождение, это будет означать, что пространственное видение возникло более одного раза. К сожалению, этот вопрос не получил особого развития со времени интенсивных дебатов три десятилетия назад (Salvini-Plawen 1982; Vanfleteren 1982). Хотя большинство типов фоторецепторных клеток, которые были исследованы, можно аккуратно классифицировать как рабдомерные или цилиарные, все еще существует ряд промежуточных случаев и клеток, которые изменяются от одного типа к другому в ходе онтогенеза (см. § 1 b ).Существует очевидный риск того, что тип наложения мембран был более эволюционно пластичным, чем предполагалось, и что различию между рабдомерными и ресничными рецепторами придали слишком большое значение. Надеемся, что широкое сравнительное определение типа опсина в фоторецепторных клетках решит эту проблему.
Jékely et al . (2008) предполагают, что рабдомерные глазки, участвующие в фототаксисе у планктонных личинок полихет ( b ), представляют собой наследственное состояние билатеральных животных.Это привлекательная идея, но она не вносит ясности в то, почему у полихет есть цилиарные фоторецепторы в головном мозге, участвующие в контроле циркадных ритмов (Arendt et al . 2004). Укладка цилиарной мембраны этих невизуальных фоторецепторов убедительно свидетельствует об их эволюционной истории как направленных фоторецепторов (см. § 2 c ; см. Дополнительные электронные материалы). Это указывает на гораздо более древнюю роль направленного фоторецепции или указывает на то, что он независимо развился в двух разных системах и что направленность (экранирующий пигмент) была вторично потеряна, когда одна из систем восстановила свою первоначальную роль в мониторинге яркости.Еще одна возможность состоит в том, что ассоциация между фоторецепцией и ресничками изначально возникла не как средство наложения мембран, а что она основана на древней функциональной связи между сенсорным контролем биения ресничек. Самодостаточная сенсомоторная функция глазковых клеток личинок коробчатой медузы ( a ; Nordström et al. , 2003) тогда была бы лучшим представителем первых направленных фоторецепторов у животных.
Многочисленные случаи укладки рабдомерных и цилиарных мембран в клетках, которые не связаны со скринингом пигментных структур (например,грамм. Purschke и др. . 2006) поддерживают идею о том, что ненаправленные фоторецептивные системы неоднократно эволюционировали из древних направленных систем, предположительно в результате дублирования и последующей потери скринингового пигмента в одной из копий. Картина дополнительно усложняется простыми глазками у плоских червей acoel ( c ), которые лишены мембранной укладки, но связаны с пигментной клеткой (Yamasu 1991). Считается, что плоские черви Acoel являются базальными билатериями и имеют прямое развитие без планктонных личинок (Ruiz-Trillo et al .1999, 2002; Телфорд и др. . 2003; Филипп и др. . 2007). Заманчиво предположить, что это восходит к системе фототаксиса у бентосных ползучих Urbilateria, а не у планктонных предков, как это было предложено Jékely et al . (2008).
Исходя из нынешних знаний, кажется, что можно создать армию различных гипотетических эволюционных сценариев, ни один из которых полностью или легко не учитывает различные наборы фоторецепторов у разных групп животных.Проблема в нынешнем виде состоит в том, что морфологические свидетельства демонстрируют полусогласованный паттерн с многочисленными исключениями и случаями, которые трудно классифицировать, и что молекулярные / генетические свидетельства типов опсинов и каскадов трансдукции пока доступны только из ограниченного набора группы и виды животных. Существует очевидная потребность в дополнительных сравнительных знаниях о типах опсинов, каскадах трансдукции и других молекулярных сигналах, прежде чем можно будет понять морфологическое разнообразие. Функциональный контекст и концепция эволюции с разделением на задачи, представленная здесь, необходимы не только для руководства формированием гипотез, но и для понимания, а не просто для описания эволюции светочувствительности и зрения.
Вот как эволюционировали глаза 500 миллионов лет назад
Глаза настолько сложны, что сам Чарльз Дарвин сказал, что идея их развития казалась «абсурдной в высшей степени». Но они это сделали, и это видео под редакцией TED показывает, как это сделать.
Ученые проследили происхождение не столь скромного человеческого глазного яблока не менее 500 миллионов лет назад, когда одноклеточные организмы, такие как крохотная эвглена, образовали простое плоское «световое пятно» на внешней стороне. их тел.Этот кластер светочувствительных белков был связан с удлиненными хвостовидными жгутиками существ, которые использовались, чтобы продвигать их по воде. Это позволяло им реагировать на источники света — и, следовательно, на пищу — и мчаться к ним.
Плоский червь под названием планария развился в несколько более сложной версии, которая работала даже лучше, потому что она была изогнута внутрь, чтобы лучше определять направление падающего света. Мало того, что червь мог использовать этот орган для поиска светоотражающей пищи, он также мог определять, где находятся очаги слабого освещения, и использовать их как укрытие от хищников.
Затем, как говорит Джошуа Харви на видео выше, в течение нескольких тысячелетий эти чашеобразные органы начали перемещаться внутри различных существ, таких как наутилус, пока не осталось только небольшое отверстие. В результате получился «эффект точечного отверстия», который резко увеличил разрешение и уменьшил искажения, потому что только тонкий луч света мог пройти через крошечное отверстие для обработки.
Но мы можем добиться большего, чем простое отверстие. Ключ к зрению в том виде, в каком мы его знаем, — это линза.Происхождение очень важной линзы восходит к слою прозрачных ячеек, которые закрывали отверстие точечного отверстия для защиты от элементов. Теперь, когда глаз закрыт, внутренняя структура глаза заполнилась жидкостью, и это действительно помогает оптимизировать светочувствительность и обработку, — говорит Харви.
За этой разработкой последовала эволюция кристаллической структуры за линзой, которая застряла вокруг, потому что она отлично фокусировала падающий свет на единственную точку на сетчатке сзади.Эта простая структура в конечном итоге стала человеческим глазом, когда начало появляться цветное кольцо, называемое радужной оболочкой, так что количество света, попадающего во внутренние части, можно было регулировать в соответствии с окружающей средой. Внешняя белая область, известная как склера, сформирована, чтобы удерживать структуру глаза на месте, и мы даже развили слезные протоки, чтобы наши глаза были защищены непрерывным запасом прозрачной защитной пленки.
Но нет никакого смысла иметь все эти крутые штуки для обработки света, если мы не знаем, что делать с любой из этой информации.По мере того, как человеческий глаз становился более сложным и эффективным, человеческий мозг также увеличивался, дополняя его. Посмотрите видео на TED выше, чтобы узнать, как это сделать.
А вы когда-нибудь задумывались, как голубые глаза приобрели свой цвет? Оказывается, они не получают свой цвет от пигмента — на самом деле это намного интереснее.
Источники: TED-ed, SPLOID. Изображение в Facebook, сделанное Стефано Гарау на Shutterstock
Эволюция и взгляд: двухсотлетие Дарвина и пятисотлетие происхождения видов | Генетика и геномика | JAMA Офтальмология
Evolution — важная концепция для всех, кто считает науку лучшим способом понять мир природы.Он настолько полно установлен, насколько может быть установлен любой научный принцип, и является великой объединяющей темой всей биологии, столь же неотъемлемой для понимания форм жизни, как гравитация для понимания космоса. К 200-летию со дня рождения Чарльза Дарвина в 1809 году и через 150 лет после публикации О происхождении видов посредством естественного отбора в 1859 году мы должны помнить об основных чертах эволюции глаза и о том важном месте, которое он занимает. в развитии и уточнении эволюционной теории.Несколько основных моментов включают древность родопсина, готовность глаза к эволюции, влияние глаз на разнообразие форм жизни и многообещающее влияние генетики на биологию развития и эволюционную биологию.
ШЕСТАЯ ГЛАВА ПРОИСХОЖДЕНИЕ ВИДОВ
Чарльз Дарвин (рис. 1) прекрасно понимал, что его идеи вызовут бурю критики со стороны широкой публики, если не научного истеблишмента.В хитроумном упреждающем ударе против «естественного богословия», креационистской идеи или «разумного замысла» того времени, Дарвин написал шестую главу первого издания журнала «О происхождении видов» «Трудности теории». это происхождение с модификацией, или то, что позже стало известно как эволюция, было произведено естественным отбором. Один из разделов этой главы озаглавлен «Органы чрезвычайного совершенства и сложности».
Предположить, что глаз со всеми его неподражаемыми приспособлениями для настройки фокуса на разные расстояния, для допуска различных уровней света и для коррекции сферических и хроматических аберраций, мог быть сформирован естественным отбором, кажется, я свободно признаться, абсурд в высшей степени.И все же разум подсказывает мне, что если будет показано, что существует множество градаций от совершенного и сложного глаза до очень несовершенного и простого, причем каждая ступень полезна для своего обладателя; если дальше, глаз действительно немного отличается, и вариации передаются по наследству, что, безусловно, так; и если какое-либо изменение или модификация органа будут когда-либо полезны животному в меняющихся условиях жизни, то трудность поверить в то, что совершенный и сложный глаз может быть сформирован естественным отбором, хотя и непреодолим для нашего воображения, вряд ли можно считать реальной. .То, как нерв становится чувствительным к свету, не волнует нас больше, чем то, как зародилась сама жизнь; но я могу заметить, что некоторые факты заставляют меня подозревать, что любой чувствительный нерв может стать чувствительным к свету, а также к тем более грубым колебаниям воздуха, которые производят звук. 1 (p217)
Первое предложение в этой цитате часто искажается, как будто Дарвин беспокоился об объяснении эволюции глаза, но на самом деле он был совершенно уверен в своей правоте.Можно было бы пожелать, чтобы он не так бесцеремонно отказывался от светочувствительности («как нерв становится чувствительным к свету, нас почти не беспокоит») — эволюция родопсина и архитектура палочек и колбочек заслужили бы его восхищение. . Можно также пожелать, чтобы его викторианская проза была более простой и прямой — последние отрывки из Origin почти поэтичны по своей элегантности — но мысль ясна. Возьмите вариацию, ее «наследуемость» (менделевская генетика в то время еще не родилась) и полезность примитивных глаз, и сложный глаз можно легко развить.
В идеале, продолжает Дарвин, было бы лучше проследить постепенное развитие сложных глаз у предков известных в настоящее время животных. Но поскольку летопись окаменелостей не допускает этого, мы должны взглянуть на весь спектр глаз у различных существующих видов, доступных нам для изучения, особенно у беспозвоночных, потому что они показывают самые широкие различия в глазах, в то время как все позвоночные уже достигли сложный глазок камеры. Если мы сделаем это:
Я не вижу большой трудности (не больше, чем в случае многих других структур) в том, чтобы поверить в то, что естественный отбор превратил простой аппарат зрительного нерва, просто покрытый пигментом и покрытый прозрачной мембраной, в оптический инструмент, как совершенный, как у любого члена большого класса Сочленяющихся [то есть позвоночных]. 1 (p218)
Затем он подчеркивает, что эти изменения, хотя и происходят постепенно в необъятном геологическом времени, могут происходить под действием естественного отбора.
Если мы должны сравнить глаз с оптическим прибором, мы должны в воображении взять толстый слой прозрачной ткани с нервом, чувствительным к свету под ним, а затем предположить, что каждая часть этого слоя постоянно медленно изменяется по плотности, так, чтобы разделиться на слои разной плотности и толщины, расположенные на разном расстоянии друг от друга, причем поверхности каждого слоя медленно меняют форму.Далее мы должны предположить, что существует сила, всегда пристально наблюдающая за каждым незначительным случайным изменением в прозрачных слоях; и тщательный отбор каждого изменения, которое при различных обстоятельствах может каким-либо образом или в любой степени иметь тенденцию к созданию более отчетливого изображения. Мы должны предположить, что каждое новое состояние инструмента умножается на миллион; и каждое должно быть сохранено, пока не будет произведено лучшее, а затем старые — уничтожены. В живых организмах вариации вызовут небольшие изменения, поколение будет умножать их почти бесконечно, а естественный отбор будет безошибочно определять каждое улучшение.Пусть этот процесс продолжается миллионы и миллионы лет; и в течение каждого года на миллионы разного рода особей; и разве мы не можем поверить в то, что живой оптический инструмент мог бы таким образом превзойти стеклянный, как творения Творца по сравнению с творениями человека? 1
Этот раздел, посвященный глазу, цитируется здесь так широко, чтобы показать, что в нем Дарвин сформулировал многие из основных положений всей книги. Кроме того, Дарвин неявно предполагал модульность эволюции, что она может изменить одну часть организма, не нарушая целостность и выживаемость организма в целом.Этим занимался Жорж Кювье, основоположник сравнительной анатомии и палеонтологии на рубеже XIX века. 2
Родопсин — древняя молекула
Термин родопсин в самом широком смысле (также теперь называемый ретинилиденовыми белками ) относится к семейству молекул, которые содержат хромофорный сегмент сетчатки (ретинальдегид или альдегид витамина А) и различные типы белковых сегментов, опсины, все они в той или иной степени связаны.Примечательно, что это конкретное семейство молекул — а их было идентифицировано более 300 — 3 — выступает в качестве светочувствительного химического вещества во всех системах зрения у существ, находящихся вверх и вниз по древу эволюции. Точные аминокислотные последовательности белковых сегментов могут различаться, и фотохимический каскад отличается своими деталями (например, циклический гуанозинмонофосфат в качестве второго мессенджера у позвоночных и инозитолтрифосфат у беспозвоночных). Тем не менее, базовое сочетание альдегида и белка витамина А является постоянной характеристикой.Если он не так распространен, как ДНК, родопсин достаточно повсеместен, что также отражает базовое единство форм жизни.
Родопсин обнаружен не только во всем домене эукариот (настоящие ядросодержащие клетки, из которых произошли простейшие, растения и животные), он также присутствует в прокариотах (микробах без ядер или других внутренних органелл), в доменах бактерий и архей. (микроскопические безъядерные клетки, изначально обнаруженные в экстремальных условиях, таких как вулканические горячие источники или гиперсоленые районы, которые отличаются от бактерий по своей биохимии).
Поскольку родопсин необходим для зрения, его обнаружение в подвижных организмах от простейших до человека неудивительно. Эти организмы находят легкое чутье настолько полезным, потому что они могут реагировать некоторым адаптивным движением на то, что они ощущают в окружающей среде. Даже самый простой светочувствительный орган позволит обнаружить тень.
Что удивительно, так это то, что прокариоты содержат также родопсин. Какую возможную пользу может иметь видение любого вида для таких примитивных организмов?
Во-первых, мы склонны забывать, что бактерии (и археи) часто подвижны, используя жгутики или волокнистые структуры, называемые пили . 4 Движение может наблюдаться у таких патогенов, как Pseudomonas aeruginosa и Neisseria gonorrhoeae , и, вероятно, способствует их патогенности. Любой тип подвижной способности позволяет организму проявлять фототаксис, способность ориентироваться и двигаться к свету или от него. Эта способность учитывает циркадные ритмы и вертикальную миграцию в глубинах океана в соответствии с циклами день-ночь. 5 В самом широком смысле это тип зрения, и родопсин делает это возможным.
Во-вторых, молекула родопсина в этих примитивных формах жизни не обязательно используется для зрения — из-за ее феноменальной способности захватывать фотоны, запускающие биохимический каскад и преобразование света в нервный импульс. Здесь родопсин может служить не в качестве молекулы , запускающей механизм , а скорее как протонный насос . Это белок, который присутствует внутри внутриклеточных мембран и служит для перемещения протонов через мембрану из одного субклеточного компартмента в другой.Это устройство для захвата энергии, используемое для передачи энергии внутри клетки и для обеспечения ее метаболизма. Так обстоит дело с родопсинами, обнаруженными в микробах, из которых наиболее изучен бактериородопсин. Кроме того, галородопсин, обнаруженный у некоторых видов, служит насосом для ионов хлора внутри клетки. 6
Бактерии и археи существовали до эукариот и животных. Эти более поздние формы жизни взяли родопсин, который, скорее всего, уже был доступен в качестве протонного насоса, и использовали его в качестве основы для зрения.Взаимодействие молекулы, служащей одной цели, и использование ее для другой цели — повторяющаяся тема в эволюции. Молекулы и метаболические пути подвержены такой кооптации, как и анатомические структуры. Evolution использует то, что доступно. Это непревзойденный переработчик.
Неясно, как именно изначально возник родопсин. Белок подобен белку, обнаруженному в сенсорных и химических рецепторах у некоторых примитивных существ, и, возможно, был заимствован из этих других чувственных модальностей.Мы можем быть уверены, что промежуточные формы молекулы имеют некоторую полезность для клетки и что эта адаптивная полезность обеспечивает удержание и развитие молекулы. В конце концов, молекулярная эволюция подчиняется тем же эволюционным принципам, которые применимы к анатомическим структурам. Такая система была постулирована при эволюции протонного насоса цитохрома c оксидазы, другой древней молекулы, которая у человека состоит из 6 белков, что даже более сложно, чем ситуация с родопсином. 7
Молекулярная эволюция — относительно новая область исследований. Это показывает, что задолго до появления многоклеточных животных и растений, которые мы знаем как ископаемые, задолго до появления микробов-эукариот, более примитивные формы жизни эволюционировали, молекулы и метаболические пути, которые обеспечивали развитие более сложной жизни. . Родопсин был там с самого начала, так хорошо выполняя свою работу, что его сохраняли на протяжении более миллиарда лет жизни на Земле.
Хотя чувствительность к свету присутствует у микробов, настоящие глаза или структуры, которые предоставляют информацию о направлении света, исходящего из различных частей окружающей среды, ограничены животными. Предсказание Дарвина об обнаружении промежуточных форм глаза полностью сбылось. Переход от светочувствительных глазных пятен, обнаруженных у некоторых простейших и ямчатых глаз червей и моллюсков, к сложным глазам типа камеры у позвоночных хорошо задокументирован у животных, живущих сегодня, и, соответственно, у их предков.Доступно несколько отличных обзоров, в которых это подробно обсуждается. 8 -15
В целом глаза различаются как минимум по 3 категориям:
1. Тип конструкции
a. «Простые» (неправильное название) или однокамерные глаза, наблюдаемые у некоторых моллюсков, пауков, морских и наземных позвоночных, сетчатка которых вогнута, а изображение перевернуто.
б. «Составные» или многокамерные глаза, которые состоят из отдельных покрытых пигментом канальцев («омматидий») в различных массивах, как это наблюдается у ракообразных и насекомых, и где эквивалент сетчатки выпуклый, а изображение прямое.
Каждый тип глаз показывает переходные формы. Представлено по крайней мере 11 оптических систем, включая зеркальную оптику, а также преломляющую оптику и, недавно обнаруженный, телеобъектив 9 (но без зум-объектива и без призмы Френеля). British Journal of Ophthalmology изображает многие из наиболее интересных глаз на обложке с забавными пояснительными эссе офтальмолога Ивана Шваба. Некоторые из этих глаз лишают воображение.
(Здесь я должен признаться в тайной любви к участку фоторецепторов на спине так называемой «безглазой» креветки, извлеченной из глубин океанской бездны, недалеко от гидротермальных жерл в срединно-океаническом хребте в центральной Атлантике.Там, вдали от любых следов солнечного света, креветки используют свою светочувствительность, чтобы ориентироваться на тусклый свет, исходящий из вентиляционного отверстия, из которого выделяются необходимые ей питательные вещества на основе серы. 16 )
2. Тип фоторецептора. Разработано как минимум 2 решения проблемы организации молекул родопсина внутри клетки таким образом, чтобы оптимизировать вероятность попадания фотона в молекулу. Оба включают изменение конформации мембраны, окружающей клетку, чтобы наложить множество слоев фотопигмента на пути фотона.
а. Мембрана рабдомерной клетки содержит множество микроворсинок, в основном присутствует у беспозвоночных и деполяризуется во время разряда.
б. Мембрана цилиарной клетки, вдавленная внутрь инвагинациями, присутствует в основном у позвоночных и гиперполяризуется при выписке.
3. Способ развития эмбриона: наиболее отчетливо это видно при сравнении глаза у позвоночных с внешне похожим глазом типа камеры, наблюдаемым у осьминога, головоногого моллюска.
а. Глаз беспозвоночного осьминога развивается в результате серии складок из эпидермальной плакоды. Фоторецепторы обращены в полость стекловидного тела.
б. Глаз позвоночного формируется из нервной пластинки и побуждает вышележащий эпидермис формировать хрусталик. Инвагинация нервной чашечки образует двухслойную сетчатку, в которой фоторецепторы обращены в сторону от полости стекловидного тела.
Линзы показывают молекулярный оппортунизм
Линзы у разных видов могут происходить из разных тканей эмбриона.Но независимо от того, какая ткань является источником, вещество, из которого состоит тело линзы, должно показывать постепенную разницу в плотности: больше в центре, что приводит к более высокому показателю преломления, менее плотное на периферии с меньшим показателем преломления. Это был эволюционный способ борьбы со сферической аберрацией, особенно актуальной проблемой для сферических линз морских позвоночных. Эту проблему предположительно признал британский ученый Джеймс Клерк Максвелл (1831-1879). Однажды за завтраком он посмотрел на поданную ему селедку и решил тут же вскрыть ей глаз.Он увидел, что линза имеет сферическую форму, и быстро понял, что сферическая аберрация ограничит эффективность глаза, если внутри линзы не будет градации показателей преломления, и так оно и было. 17
Фактические вещества, используемые в хрусталике позвоночных — кристаллические белки — когда-то считались уникальными для хрусталика и были разработаны специально для этой цели. В настоящее время признано, что они похожи на белки, используемые для других целей, и занимают свое место с другими белками, которые, по-видимому, были получены из ферментов или других факторов.Фернальд 10 называет этот молекулярный оппортунизм и противопоставляет его широко распространенной зависимости от родопсинов.
Глаз полезен при классификации видов
Поскольку с помощью секвенирования аминокислот теперь можно детально изучить различия в различных белках опсина, присутствующих в фоторецепторах позвоночных, и поскольку эти белки тесно связаны друг с другом, этот метод используется для особенно тонких различий в точном местонахождении животных. на эволюционном дереве. 13 Например, фотопигменты аллигатора более тесно связаны с фотопигментами кур и, предположительно, других птиц, чем с другими рептилиями или млекопитающими. 18
Когда появились первые глаза?
В качестве мягких частей глаза в окаменелостях обычно не сохранились. Тем не менее, глаза, состоящие в основном из линз кальцита, случайно окаменели у некоторых ранних трилобитов, ныне вымершего типа членистоногих, существовавшего почти 300 миллионов лет.Видно, что они присутствуют в виде уже хорошо развитых сложных глаз в самом начале кембрийского периода около 540 миллионов лет назад (рис. 2). Первые настоящие глаза, должно быть, появились незадолго до этого, а окаменелости трилобитов стали кульминацией их предшествующей эволюции. Но поскольку трилобиты (или большинство других окаменелостей животных) не встречаются до этого периода, эти первые глаза нельзя увидеть напрямую. 17 , 20
Еще одно свидетельство важности зрения в то время — это то, что внешне эти древние существа были цветными.Parker 17 изучил окаменелости из знаменитых сланцевых пород Берджесс в Канадских Скалистых горах, которые ясно показывают сохранившиеся мягкие части у существ более позднего кембрия. У многих видны окаменелые составные глаза, некоторые из них до крайности причудливы — у одного существа длиной всего несколько миллиметров глаз в два раза меньше его всего тела. Электронные микрофотографии показывают наслоение твердых частей, сохранившихся в этих окаменелостях. Эти слои могли действовать как дифракционная решетка, придавая существу радужный металлический блеск во всех цветах видимого спектра.Паркер действительно визуализировал такой цвет; он мог предупреждать хищников, которых, по-видимому, тоже нужно было заметить.
Vision предоставил такой актив для выживания, что он, возможно, привел к «гонке вооружений» между хищником и жертвой, с разработкой панцирей для защиты и модифицированных придатков и кусающих ртов для нападения. Заманчиво думать о зрении как о главном среди всех факторов, влияющих на выживание этих ранних существ и тем самым ускоряющих их эволюцию. 17
Что можно сказать, так это то, что вскоре после появления глаз животная жизнь, кажется, разлилась во множество различных форм. (До сих пор остается вопросом, было ли такое кажущееся разнообразие действительно новым или просто могло быть оценено по-новому из-за развития ископаемых твердых частей.) Большинство из 35 типов, признанных в настоящее время у современных форм жизни, произошли от трех или около того. присутствует в начале кембрия. Из этих 35 типов у 6 есть глаза: книдарии (медузы), моллюски (улитки, моллюски, кальмары и осьминоги), кольчатые черви (сегментированные черви), онихофоры (бархатные черви), членистоногие (насекомые, пауки и ракообразные) и хордовые (позвоночные).Эти 6 типов содержат около 96% известных видов, живущих сегодня. Возможно, действительно был «оптический взлет» для видов с глазами. 9 , 10
Сколько времени потребуется, чтобы сложный глаз развился?
Для тех, кто сомневается, что естественный отбор мог развить такую сложную структуру, как глаз, испытанный и верный ответ подчеркнул длительный период геологического времени и короткую продолжительность жизни каждого поколения ранних форм жизни.Но в этом долгом периоде беременности, возможно, не было необходимости.
В 1994 году Дэн-Эрик Нильссон и Сюзанна Пелгер опубликовали математическую модель перехода от ямочного глаза к однокамерному глазу позвоночного типа. 20 Названный «Пессимистическая оценка времени, необходимого для развития глаза», он поразил читателей, поскольку показал, насколько быстро может гипотетически развиваться сложный глаз. Их модель стоит обсудить на мгновение, потому что она поднимает некоторые соображения — анатомические ограничения — связанные с происхождением целых органов.
Авторы сделали консервативные, а следовательно, «пессимистические» оценки последовательных шагов, необходимых для трансформации плоского участка фоторецепторов, расположенного между прозрачным защитным слоем с одной стороны и темным пигментированным слоем ткани с другой (Рисунок 3). По мере того, как слои фоторецепторного пигмента изгибаются назад, образуя ямку, способность просто замечать присутствие тени улучшается, давая некоторое ощущение направления света. По мере продолжения этого процесса передний конец начинает сужаться, превращаясь в отверстие с радужной оболочкой.С появлением линзы разрешение еще больше улучшается, особенно когда улучшается градуированная плотность материала линзы.
Предполагая, что ткани будут различаться не более чем на 1% по ширине, длине, показателю преломления и т.д. во время каждого шага, Нильссон и Пелгер подсчитали, что в целом потребуется менее 2000 шагов. (Вас не впечатлило изменение на 1%? Подумайте об этом: 2000 таких изменений могут протянуть достаточно пальца, чтобы дотянуться до Атлантики. 12 ) Одновременно может произойти изменение только одного объекта, еще одна «пессимистическая» особенность, которая может иметь тенденцию к переоценить необходимое время.Еще один важный момент: каждый шаг не будет мешать работе промежуточной формы.
Далее они предположили медленную скорость эволюции, то есть, что у некоторых представителей вида уже есть вариация, передающая дальнейшую благоприятную вариацию следующему поколению. Это изменение немного улучшит зрение и, таким образом, повысит выживаемость, так что оно будет распространяться в популяции вида с каждым последующим поколением. Фактические наблюдения натуралистов в этой области дали значения для таких соображений, как наследуемость, вероятность изменчивости и интенсивность отбора. 21 Оказалось, что менее 400 000 поколений будет более чем достаточно для развития сложного глаза. Поскольку аналогичные существа, живущие сегодня, имеют короткую продолжительность жизни, можно с уверенностью предположить, что поколение займет год. Таким образом, можно разумно оценить весь ход изменения, который займет менее полумиллиона лет — всего лишь мимолетный момент в геологической шкале времени.
Обратите внимание, что модель начинается с фоторецепторов, уже установленных в простой глазковой точке. Оценка относится к однокамерному глазу, но первый глаз был составным.Может ли эта схема применяться и к сложным глазам? Лэнд и Нильссон 12 задумывают рыхлую совокупность глазных пятен у предка трилобитов, изменяющуюся, чтобы сформировать их сложные глаза аналогичным образом. Таким образом, все это могло произойти незадолго до появления самых ранних окаменелостей кембрия или около 540 миллионов лет назад.
Глаз развивался один или несколько раз?
Это создает вопрос с подвохом, потому что и на «один раз», и на , и на «много раз» может быть да, в некотором роде.В классической работе 1977 года прослеживается разновидность глаз у разных линий животных и делается вывод о том, что глаза эволюционировали отдельно по крайней мере 40 раз, а возможно, даже 60 раз в истории форм жизни животных. 22 Эта так называемая «полифилетическая» концепция происхождения глаз теперь является предметом результатов экспериментальной генетики, которые заставляют некоторых ученых отстаивать «монофилетическое» происхождение под влиянием так называемых основных контрольных генов. Эта загадка расширяет наши представления о том, как именно генетические программы влияют на развитие органов в целом.
Экспериментальные генетики в 1980-х определили гомеобокс, последовательности ДНК в генах, которые были особенно важны в регуляции развития формы в эмбрионе. Эти гены гомеобокса кодируют факторы транскрипции, которые включают каскады программ из других генов.
В 1995 году Хальдер и др. Сообщили об экспериментах на надежном лабораторном животном Drosophila melanogaster , плодовой мушке. 23 Им удалось выделить его гомеобоксный ген eyeless. ( Drosophila биологи традиционно называют ген из-за эффекта, который его инактивация оказывает на развитый организм; полная деактивация этого гена на ранней стадии развития плодовой мухи вызывает рождение человека без глаз.) Но на этот раз они намеренно активировали безглазый Ген в соматических клетках эмбриона, у которых глаза обычно не развиваются. Эта манипуляция вызвала развитие эктопических глаз на ногах, крыльях и даже на кончиках усиков.Глаза были анатомически законченными, и фоторецепторы могли активироваться светом. Затем они превзошли свой результат, введя эквивалентный ген Sey («маленький глаз»), взятый у мыши, включив его и вызвав тот же эффект — эктопическое развитие сложного глаза мухи с геном позвоночного животного. 24
Эти поразительные эксперименты сделали ученых похожими на колдунов. Было обнаружено, что гены мыши и мухи имеют высокую степень сходства по аминокислотным последовательностям.Их назвали «главными управляющими генами» для развития глаз, и аналогичные гены были обнаружены во всем спектре животных у червей, кальмаров и людей, у которых его имя — Pax-6 . (Мутационное изменение в Pax-6 связано с различными аномалиями переднего сегмента. 25 -27 Гомозиготная потеря вызывает анофтальмию с пороком развития мозга, достаточно серьезным, чтобы убить новорожденного, показывая, что Pax-6 не только важен для развития глаз, но и для развития самого мозга. 28 )
Одно дело — знать, что ДНК объединяет все живые существа способом, неизвестным Дарвину. Еще более унизительно было узнать, что настоящие наборы генных инструментов для построения глаз и других органов были настолько интенсивно сохранены в ходе эволюции, что люди разделили их с мышами и, mirabile dictu , плодовой мушкой.
Однако концепция Pax-6 как главного управляющего гена, который однажды эволюционировал, а затем включил вторичные программы, которые фактически конструируют самые разные глаза, вероятно, слишком проста. Pax-6 выполняет и другие важные функции, в частности, при формировании головы. Он присутствует у некоторых существ без глаз, в том числе у некоторых медуз. Другие гены также могут включать развитие глаз, и они могут находиться выше Pax-6 . По словам Фернальда:
Генетический контроль строения глаза ошеломляюще сложен, и кажется более вероятным, что интерактивная «генная сеть» регулирует развитие всех сложных органов, включая глаз. . . . Поскольку Pax-6 существовал перед глазами, как и другие важные для развития гены, он, возможно, был задействован в формировании глаз, как и опсин. 10 (п448)
К 1996 году было нанесено на карту по крайней мере 72 гена, участвующих в развитии глаз мыши, и, несомненно, сейчас их стало больше. 29
Другими словами, Pax-6 мог быть набран в разное время для разных животных, так что даже если сложные генетические программы подстерегали, они не всегда включались, а сами глаза эволюционировали много раз. Весь этот вопрос заставляет переосмыслить классическую эволюционную концепцию гомологии, идею о том, что сходные структуры у разных видов могут быть идентифицированы как связанные благодаря общему происхождению. 29 -32
Онтогенез повторяет филогению
Я впервые столкнулся с этой загадочной фразой, отображаемой над доской в классе естественных наук, как введение в биологию в средней школе. Он обладал торжественной магической силой религиозного изречения. В то время он уже был в немилости, если не полностью устарел, в течение 50 лет.
Это выражение было придумано Эрнстом Геккелем (1834-1919), который хотел объявить «биогенетический закон», гласящий, что стадии развития эмбриона позвоночных напоминают взрослые формы существ, которые появились раньше в ходе эволюции. Я полагаю, он надеялся, что такие великие принципы объяснят мириады реалий биологических форм, и был слишком впечатлен физиками того времени, которые унаследовали стабильные математические «законы» и были заняты в то время формулированием новых единицы.Но эволюция сделала биологию исторической наукой, и закон — неправильный термин для описания закономерностей, с которыми он работает.
По словам Стивена Джея Гулда, у которого был тот же опыт, что и у меня с фразой Геккеля, «биогенетический закон Геккеля был настолько экстремальным, а его коллапс — настолько впечатляющим, что вся тема стала табу». 33 (p2) Он держался в разбавленной форме, что привлекло внимание, например, к временному появлению жаберных щелей у эмбриона млекопитающих, не считаясь при этом много объяснимым ни в эмбриогенезе, ни в эволюции.До недавнего времени эмбриологи и эволюционисты не разговаривали друг с другом. Это было очень плохо, потому что эволюцию можно рассматривать как в основном взаимодействие среды с развитием. Игнорирование эмбриогенеза было ошибкой, особенно с учетом того, что сам Дарвин часто ссылался на эмбриологию в «Происхождении » и в частных письмах. 34 (стр.282)
Здесь мы должны помнить, что глаз был важен для ранней эмбриологии, поскольку он помог сформулировать концепцию индукции, определяемую как ткань, влияющая на последующее развитие другой.Именно на развивающемся глазу лягушки Ганс Спеманн (1869-1941) провел свои знаменитые эксперименты в 1901 году. Он увидел, что обычно хрусталик развивается из вышележащей эктодермы. Но если до развития хрусталика он вырезал нижележащий зачаток глаза, хрусталик не смог бы сформироваться. Трансплантация зачатка глаза в другие части эмбриона вызвала развитие хрусталика в вышележащей эктодерме, где бы он ни находился. Более поздние эксперименты, стимулирующие развитие нервной трубки, привели к концепции организатора. Организатор явился прорывом в понимании эмбриогенеза и произвел фурор в свое время. 35 (p163)
Однако, помимо этих ключей, эмбриология описала развитие, но не могла сказать, почему оно происходило определенными способами, а не другими. Эмбриологи не присоединились к современному синтезу классического дарвинизма и популяционной генетики, когда он был сформулирован в 1930-х и 1940-х годах. Потребовалась экспериментальная генетика, чтобы показать, что важны не только гены, но и эта форма определяется тем, когда и где они выключаются, включаются и как долго.Большим сюрпризом стало обнаружение того, что аналогичные подходы к формированию паттерна эмбриона применимы для видов, которые так далеко разошлись по эволюционной шкале. Нынешнее сближение между эволюционными биологами, молекулярными генетиками и биологами, занимающимися вопросами развития (известное как «evo-DevO»), проясняет, как никогда ранее не было возможности, как могут происходить как маленькие, так и большие изменения формы. 34 , 36 , 37 Очевидно, что существуют основные процессы в развитии, посредством которых изменения генотипа — будь то мутация, рекомбинация генов или перегруппировка хромосом во время полового размножения — могут иметь глубокое влияние на фенотип и ускорять эволюцию большие изменения.Генетические переключатели, которые включают или выключают гены, настолько важны для определения фенотипа, что являются особенно влиятельными участками эволюции. Понимание этого может облегчить нам понимание того факта, что в геноме человека всего 22 500 генов, что всего в 6 раз больше, чем в геноме бактерии. 37
Дарвин построил свою теорию на трех основных идеях: произошли изменения, что наследственность передала некоторые из них следующему поколению, и этот естественный отбор определил, какие из них останутся и будут развиваться с течением времени.Он не знал задействованных генетических и клеточных механизмов, и с тех пор наука активно ставит под сомнение, исследует и изменяет его идеи. Теперь, через 150 лет после рождения О происхождении видов , то, что он назвал , продолжается один длинный аргумент , обещающий еще больше понимания природы жизни на Земле.
Для тех читателей, которые зашли так далеко и все еще интересуются этим элегантным заключительным отрывком в Origin , вот он:
В этом взгляде на жизнь есть величие, с его несколькими силами, первоначально вдохнувшимися в несколько форм или в одну; и что, пока эта планета двигалась в соответствии с фиксированным законом всемирного тяготения, с самого начала развивались и развиваются бесконечные формы, самые прекрасные и самые чудесные. 1 (p459-460)
Для корреспонденции: Ronald S. Fishman, MD, 47880 Cross Manor Rd, Saint Inigos, MD 20684 ([email protected]).
Допущено к публикации: 6 апреля 2008 г .; окончательная доработка получена 22 апреля 2008 г .; принята к печати 22 апреля 2008 г.
Раскрытие финансовой информации: Не сообщалось.
1.
Дарвин
C О происхождении видов путем естественного отбора или сохранении благоприятных рас в борьбе за жизнь [первоначально опубликовано в Лондоне, Англия: J Murray; 1859].Нью-Йорк, Нью-Йорк Краун Паблишерс Авенел Букс, 1979;
2.
Гулд
SJ Восемь поросят: размышления в естествознании. Нью-Йорк, NY Norton1993; 254
3. Спудич
JLYang
C-HJung
КХСпудич
RU Ретинилиденовые белки: структуры и функции от архей до человека. Annu Rev Cell Dev Biol 2000; 16365-392 PubMedGoogle ScholarCrossref 5.Armitage
JPHellingwerf
KJ Поведенческие реакции, индуцированные светом («фототаксис») у прокариот. Photosynth Res 2003; 76.
(1-3)
145-155PubMedGoogle ScholarCrossref 6.Oesterhelt
DStoeckenius
W Функции новой фоторецепторной мембраны. Proc Natl Acad Sci U S A 1973; 70
(10)
2853–2857PubMedGoogle ScholarCrossref 8. Кронли-Диллон
JR Грегори
RL Эволюция глаза и зрительной системы. Бостон, Массачусетс, CRC Press, 1991; Зрение и зрительная дисфункция том 2Google Scholar12.Land
MFNilsson
D-E Глаза животных. Oxford, England Oxford University Press, 2002; Oxford Animal Biology series Google Scholar14.Lamb
TDCollin
SPPugh
EN
Jr Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и наглазник. Nat Rev Neurosci 2007; 8
(12)
960- 975PubMedGoogle ScholarCrossref 16. Ван Довер
CLSzuts
EZChamberlain
SCCann
JR Новый глаз у «безглазых» креветок из гидротермальных источников Срединно-Атлантического хребта. Природа 1989; 337
(6206)
458-460PubMedGoogle ScholarCrossref 17.
Parker
A В мгновение ока: как видение спровоцировало большой взрыв эволюции . Нью-Йорк, NY Basic Books, 2004; 194224
19.
Форти
R Трилобит! Свидетель эволюции . Нью-Йорк, Нью-Йорк Альфред А. Кнопф2000; Рис. 10 (рядом с стр. 80)
20. Нильссон
DEPelger
S Пессимистическая оценка времени, необходимого для развития глаза. Proc Biol Sci 1994; 256
(1345)
53- 58PubMedGoogle ScholarCrossref 22. Сальвини-Плавен
LVMayr
E Об эволюции фоторецепторов и глаз. Evol Biol 1977; 10207-269Google Scholar 23.Halder
GCallaerts
PGehring
WJ Индукция эктопических глаз путем направленной экспрессии гена eyeless у Drosophila . Наука 1995; 267
(5205)
1788-1792PubMedGoogle ScholarCrossref 25. Мирзаянс
FPearce
WGMacDonald
IMWalter
MA Мутация гена PAX6 у пациентов с аутосомно-доминантным кератитом. Am J Hum Genet 1995; 57
(3)
539-548PubMedGoogle Scholar26.Glaser
TJepeal
LEdwards
JG
и другие. PAX6 Эффект дозировки гена в семье с врожденной катарактой, аниридией, анофтальмией и дефектами центральной нервной системы. Нат Генет 1994; 7
(4)
463-471PubMedGoogle ScholarCrossref 27.Hanson
IMFletcher
JMJordan
Т
и другие. Мутации в локусе PAX6 обнаруживаются при гетерогенных пороках развития переднего сегмента, включая аномалию Петерса. Нат Генет 1994; 6
(2)
168-173PubMedGoogle ScholarCrossref 28.Hever
AMУильямсон
Каван Хейнинген
V Пороки развития глаза: роль PAX6 , SOX2 и OTX2. Clin Genet 2006; 69
(6)
459-470PubMedGoogle ScholarCrossref 31.Wray
GA Эволюционные диссоциации между гомологичными генами и гомологичными структурами. Бок
GRCardew
G Серия симпозиумов Фонда Новартис. Хобокен, штат Нью-Джерси, John Wiley & Sons, 1999; 189–206, Google Scholar 33.
Гулд
SJ Онтогенез и филогения. Кембридж, MA Harvard University Press, 1977;
34.
Кэрролл
SB Самые красивые бесконечные формы: новая наука Эво-Дево. Нью-Йорк, Нью-Йорк WW Norton2005;
35.
Mayr
E Это биология. Кембридж, MA Harvard University Press, 1997; 169-175
36.
Raff
BA Форма жизни: гены, развитие и эволюция формы животных. Чикаго, Иллинойс, Университет Чикаго, 1996;
37.
Киршнер
MWGerhart
JC Правдоподобие жизни: разрешение дилеммы Дарвина. Нью-Хейвен, CT Yale University Press, 2005;
Кислород сформировал эволюцию глаза
ИЗОБРАЖЕНИЕ: Эволюция размера глаза (A) и сетчатки (B). Эволюция структур, дополняющих снабжение сетчатки кислородом, тесно связано с эволюцией больших глаз…
посмотреть еще
Кредит: Кристиан Дамсгаард, AU
.
Конвергентное происхождение новых механизмов подачи кислорода к сетчатке было напрямую связано с одновременным улучшением функциональной анатомии глаза.
В своей «Происхождении видов» Дарвин использовал сложность глаза, чтобы аргументировать свою теорию естественного отбора, и с тех пор глаз продолжает очаровывать и беспокоить эволюционных биологов.
В статье, опубликованной сегодня в eLife , исследователи из Орхусского университета объединились с учеными из восьми международных организаций, чтобы изучить физиологические требования для развития улучшенного зрения.
Они утверждают, что развитие остроты зрения у древних животных сдерживалось их способностью доставлять достаточное количество кислорода к клеткам сетчатки. Их исследование выявило удивительный паттерн механизмов улучшения кислородного снабжения сетчатки, который эволюционировал вместе с улучшенной морфологией сетчатки для улучшения зрения. Модель подходит для всех костных позвоночных, от рыб до птиц и млекопитающих. Эти открытия добавляют дополнительный компонент к нашему пониманию эволюционного глаза, которое на протяжении веков очаровывало и беспокоило эволюционных биологов.
Повышение и понижение кислородного снабжения сетчатки
В исследовании использовалось разнообразие физиологии и анатомии глаз 87 видов животных, включая рыб, земноводных и млекопитающих. Поместив эти виды на древо жизни, авторы раскрыли эволюционную историю глаза от вымершего предка современных позвоночных, которому 425 миллионов лет, до современных животных. Они определили три различных физиологических механизма подачи кислорода в сетчатку, которые всегда связаны с улучшением зрения.Таким образом, у рыб мутации в гемоглобине были связаны со способностью доставлять кислород к сетчатке при исключительно высоком парциальном давлении кислорода для преодоления значительного расстояния диффузии до клеток сетчатки.
Авторы показывают, что происхождение этого механизма около 280 миллионов лет назад было связано с резким увеличением размера глаза и толщины сетчатки, что напрямую связано с улучшением светочувствительности и пространственного разрешения. Этот механизм в гемоглобине впоследствии несколько раз терялся, возможно, чтобы избежать окислительного повреждения и образования пузырьков газа в глазу.
Теплокровные динозавры сформировали мировоззрение млекопитающих
Авторы показывают, что повышенная зависимость от зрения у млекопитающих была связана с эволюцией капиллярного русла внутри сетчатки, несмотря на потенциальный компромисс для остроты зрения, вызванный отклонением света эритроцитами.
Капилляры сетчатки у млекопитающих возникли около 100 миллионов лет назад, когда динозавры развили эндотермию. Эндотермия позволила этим мезозойским динозаврам охотиться ночью, что вынудило ранее ведущих ночной образ жизни млекопитающих вести дневной образ жизни с повышенной надежностью зрения.
Новая модель эволюции глаза показывает, что эволюция внутриретинальных капилляров точно совпала с улучшением зрения около 100 миллионов лет назад. Кроме того, это показывает, что некоторые млекопитающие потеряли капилляры сетчатки, когда они стали меньше полагаться на зрение (например, эхолокационные летучие мыши).
Кислород и зрение идут рука об руку
В целом, этот анализ показывает, что функциональная морфология глаза динамически менялась на протяжении эволюции животных.Это показывает, что морфология глаза идет рука об руку с параллельными изменениями в снабжении сетчаткой кислородом, и они, вероятно, вызваны различными компромиссами с кислородным снабжением сетчатки. Эти компромиссы кажутся приемлемыми вместо улучшенной остроты зрения, доступной при увеличении толщины сетчатки.
В целом, это исследование показывает, что адаптация, обеспечивающая доставку кислорода к сетчатке, была физиологической предпосылкой функционального развития глаза.
###
Заявление об ограничении ответственности: AAAS и EurekAlert! не несут ответственности за точность выпусков новостей, размещенных на EurekAlert! участвующими учреждениями или для использования любой информации через систему EurekAlert.
Eye Origins: как эволюция смогла создать сложный глаз
Насколько выгодно зрение? Глупый вопрос — спрашивать, насколько это выгодно видеть, не так ли? Очевидно, что иметь возможность видеть вещи намного лучше, чем не иметь представления о мире. Иногда кажущиеся глупыми вопросы могут прояснить некоторые интересные идеи, если мы просто сделаем паузу, чтобы удовлетворить их. Солнечный свет на самом деле является важнейшим фактором естественного отбора. Одним из примеров этого является тот факт, что большинство живых клеток на Земле давным-давно развили ферменты репарации ДНК, чтобы исправить повреждения, нанесенные молекулам ДНК, которые постоянно подвергаются бомбардировке вредным ультрафиолетовым излучением Солнца.
Еще один способ воздействия света на естественный отбор — зрение. Свет позволяет животному развивать новую сенсорную модальность, оценивая окружающую среду на другом расстоянии (вид) и давая ему возможность лучше ориентироваться в окружающей среде, чтобы найти пищу или спастись от хищников. Если бы это было правдой, мы бы ожидали увидеть появление разных точек зрения. Более того, если определенный тип глаза особенно хорош для сбора и обработки визуальной информации, можно ожидать, что он будет развиваться независимо во многих живых организмах.Дело в том, что мы действительно видим это у существующих живых существ.
Недавнее открытие, демонстрирующее это, было сделано доктором Майклом Боком и его коллегами, и они опубликовали его в журнале Current Biology . Ученые изучали вид веерных червей ( Megalomma interrupta ), обитающих на морском дне. Эти черви вытягивают свои щупальца вверх, чтобы брать пробы воды и частицы пищи. Интересно, что они развили сложные глаза на своих щупальцах, которые могут обнаруживать тени и, следовательно, движение, чтобы защитить себя от любых приближающихся угроз.Исследователи использовали метод секвенирования для изучения генов, экспрессируемых в глазах веерного червя, известных как транскриптомика. Примечательно, что они обнаружили, что белки, участвующие в обнаружении света у этого беспозвоночного, больше напоминают белки сетчатки многих позвоночных животных. Их результаты показывают, что необычные глаза этого веерного червя, скорее всего, возникли независимо, чтобы удовлетворить их потребности.
Итак, давайте пройдемся по стадиям эволюции нашего глаза и посмотрим, сколько раз стадия развивалась независимо, как, возможно, глаз веерного червя.
От маленького несовершенного глаза к «идеальному»
Точно так же, как небольшая скорость уже дает вам преимущество перед другими, способность видеть чуть лучше может сделать вас более подходящим. Дарвин предположил в своей книге О происхождении видов , что если вы начнете с «маленького и несовершенного глаза», а затем постепенно добавляете к нему больше функций в течение длительного периода времени, вы можете в конечном итоге получить глаз как «идеальный», как у нас. Итак, какие доказательства в животном мире могут указывать на эти стадии эволюции?
Есть небольшой одноклеточный организм, известный как эвглена.Все это животное состоит из одной клетки и имеет небольшой жгутик (хвост), который позволяет ему плавать. Если вы посмотрите на него под микроскопом, он будет больше похож на зеленую каплю. Этот зеленый цвет — хлоропласт, который он использует для фотосинтеза и питания. Фотосинтез требует сбора света, и вам нужно видеть его светом, чтобы подплыть к нему, но эвглена одноклеточная и у нее нет глаза. Ну, в каком-то смысле это так. Среди всего зеленого, что вы видите под прицелом, есть небольшой красноватый пигмент, называемый глазным пятном, который чувствителен к свету.Эвглена использует свое глазное пятно, чтобы подвергнуться процессу, называемому фототаксисом, с помощью которого он перемещается в направлении источника света. Способность обнаруживать солнечный свет позволяет этому организму подплывать к нему и использовать его для фотосинтеза. Простые глазные пятна, подобные глазам Эвглены, эволюционировали независимо как минимум 40 раз в животном мире.
Итак, это простейшее устройство для обнаружения света, которое вы можете получить — простое светочувствительное пятно, которое может обнаруживать очень элементарную информацию о направлении. Теперь мы можем представить, что наличие листа светочувствительных клеток (фоторецепторов) было бы следующим шагом, потому что он позволил бы нам собирать больше света, тем самым увеличивая чувствительность.Но как эволюция может улучшить лист фоторецепторов? Если бы какие-то случайные мутации каким-то образом вызвали вмятину на этом листе, это дало бы значительно больше визуальной информации.
Планарийный червь — плоский червь, который дает нам представление о следующей стадии эволюции глаза. Глаз планарии представляет собой чашу по обе стороны от головы червя. Эти глаза представляют собой скопления фоторецепторов, которые располагаются в так называемом наглазнике — результат постепенного вдавливания. Эти крошечные черви используют информацию, полученную из наглазников, чтобы уйти из ярко освещенной среды, где они наиболее уязвимы, и спрятаться в более темных, менее уязвимых местах.Купирование дает животному больше информации о направлении, чем глазное пятно, потому что теперь оно может видеть тени. Возможно, развитие этого типа зрения является более сложной задачей с эволюционной точки зрения, поскольку только шесть типов животных имеют эту систему. Тем не менее, это свидетельство того, насколько полезен этот вид глаза, поскольку на эти шесть типов приходится более 95% всех живых существ на Земле.
Если бы это медленное углубление продолжалось, постепенно углубляясь по мере развития животного, какой была бы следующая стадия? Представьте, что по мере того, как углубление становится глубже, внешние края ткани начинают смыкаться, образуя почти сферическое образование.Чем глубже становится наглазник, тем более сложную визуальную информацию он может собирать, повышая физическую форму животного на каждом шагу. Этот прогресс может продолжаться до тех пор, пока наглазник не превратится в полностью замкнутый круг, и в этом случае свет не сможет проникнуть внутрь и животное останется слепым, или останется крошечный промежуток в самом начале круга. Этот зазор называется точечным отверстием.
Глаз наутилуса именно такой, глаз-торец. Глаз моллюска является простым отверстием, через которое водой может течь свободно, и она не имеет роговицы или объектива, а это означает, что отсутствует какая-либо огнеупорная способность.Тем не менее, глаз-обскура — это самое высокое разрешение, которое вы можете получить без роговицы или хрусталика, что значительно улучшает наглазник. Однако, как это часто бывает в эволюции, такое разрешение требует компромисса. Этот глаз не пропускает много света через маленькое отверстие, и поэтому у наутилуса очень тусклое зрение. Такие глаза-точечки существуют сегодня у немногих видов, кроме наутилусов, поскольку большинство животных развили средства зрения с большей разрешающей способностью и чувствительностью.
The Perfect Eye
Вы, наверное, уже догадались, как улучшить прорезь для глаз.Следующим этапом будет создание какого-то тонкого слоя покрытия над этим отверстием. На самом деле подойдет все, даже кожа, при условии, что она относительно полупрозрачна и пропускает свет. Это основа для формирования так называемой роговицы, самого внешнего слоя глаза. Но почему природа пошла на то, чтобы закрыть точечное отверстие и начать развитие роговицы; другими словами, какую пригодность это дает? Прикрытие этой дыры делает три вещи. Во-первых, он защищает глаза от инфекции, поскольку представляет собой физический барьер для бактерий.Во-вторых, он обладает способностью преломлять свет, давая животному возможность собирать больше визуальной информации и потенциально преобразовывать ее в более четкое изображение. В-третьих, закрытие этого отверстия позволяет организму сформировать в глазу особый юмор. Поскольку вода больше не может просто течь в глаз и выходить из него, постепенно может образоваться специализированная жидкость, защищающая сетчатку от вредного излучения, и в конечном итоге за роговицей может образоваться линза для более точной фокусировки полученной из света информации.
Когда Дарвин писал О происхождении видов , он не знал двух концепций.Во-первых, Дарвин ничего не знал о генетике. Грегор Мендель, отец генетики, открыл законы наследственности и вскоре после этого Дарвин выдвинул свою теорию, но Дарвин никогда не знал о своей работе. Во-вторых, Дарвин понятия не имел о ДНК. Только после того, как генетика стала полем деятельности, ученые различных дисциплин поспешили открыть субстраты наследственности — основные строительные блоки жизни, которые, как мы теперь знаем, являются ДНК. И все же его догадка была довольно близка к реальности. В этом объясняющая сила теории.Вам не всегда нужно знать детали, чтобы иметь возможность дать эволюционное объяснение чему-либо; вам нужно только понять саму теорию.
Мы начали с простого светочувствительного пигмента (несовершенный глаз) и закончили глазом с полной камерой, защищенным от любых внешних патогенов. Теперь природа может начать экспериментировать с различными компонентами глаза, постепенно совершенствуясь, чтобы улучшить физическую форму животного в зависимости от его потребностей в окружающей среде. Наш глаз — такой глаз, и он отлично помогает нам правильно функционировать в окружающей среде.