Физиология ССС (Заика) — Стр 3
Объемная скорость — это количество крови протекающее через поперечное сечение сосуда в ед. времени (1 мин). В норме отток крови от сердца равен ее притоку к нему, это означает, что объемная скорость является величиной постоянной.
Линейная скорость — это скорость движения крови вдоль сосуда. Она различна в отдельных участках сосудистого русла и зависит от общей суммы площади просветов конкретного отдела сосудов.
Ваорте поперечное сечение равно 8 см2 (Д = 3 см), скорость движения крови составляет 50–70 см/с. В капиллярах общее сечение 8000 см2, скорость движения крови 0,05 см/с.
Вартериях скорость кровотока 20–40 см/с, артериолах — 0,5–10 см/с,
вполой вене — 20 см/с.
Всвязи с выбросом крови в сосуды отдельными порциями, кровоток в артериях имеет пульсирующий характер.
Непрерывность тока по всей системе сосудов связана с упругими свойствами аорты и артерий. Основная кинетическая энергия, обеспечивающая движение крови, сообщается ей сердцем во время систолы. Часть этой энергии идет на проталкивание крови, другая — превращается в потенциальную энергию растягиваемой стенки аорты и артерий во время систолы. Во время диастолы эта энергия переходит в кинетическую энергию движения крови.
Часть этой энергии идет на проталкивание крови, другая — превращается в потенциальную энергию растягиваемой стенки аорты и артерий во время систолы. Во время диастолы эта энергия переходит в кинетическую энергию движения крови.
2.Морфологическая и функциональная классификация сосудов
Амортизирующие сосуды — аорта, легочная артерия, др. крупные со-
суды. Содержат эластические элементы. Здесь сглаживаются подъемы АД при систоле.
Резистивные — артерии и артериолы. Гладкомышечные стенки способны значительно изменять диаметр сосуда, они регулируют кровоснабжение органов.
Сосуды сфинктеры — последние участки прекапиллярных артериол. Изменяядиаметрартериолопределяютчислофункционирующихкапилляров.
Обменные сосуды — капилляры. Строение стенок капилляров способствует обмену веществ.
Емкостные сосуды — венулы, вены. Стенки их тоньше артериальных, легко растяжимы, содержат клапаны. Вмещают много крови (особенно в венах печени, брюшной полости, подсосочкового сплетения кожи).
Шунтирующие (анастамозы) — связывают артерии с венами минуя капилляры. Участвуют в регуляции периферического кровотока, температуры частей тела. Это сосуды уха, носа, стопы и др.
3. Движение крови по сосудам высокого давления (артерии)
Все сосуды выстланы изнутри слоем эндотелия, образующего гладкую поверхность. Это препятствует свертыванию крови в норме. Кроме этого,
Регуляция работы сердца
3
Регуляция работы сердца.
Регуляция работы сердца – это изменение
его деятельности в соответствии с
потребностями организма. Результатом
изменения работы сердца является МОК.
МОК = ЧСС • СВ. Регулирующие механизмы
могут обеспечить изменение МОК через
каждую из этих величин.
МОК при различных условиях у
здорового человека.
Показатели | СВ | ЧСС | МОК |
Состояние покоя | 60 – 80 мл. | 60 – 85 в мин. | 4,5 – 5л. |
Максимум активности | 130 мл. | 240 . | До 30л. |

Изменение МОК наблюдается при ортостатике,
гиподинамии, физической работе,
эмоциональном напряжении, в экстремальных
ситуациях.
Классификация механизмов, регулирующих
деятельность сердца.
Различают клеточный, интраорганный и
экстракардиальный уровень регуляции.
Регулирующие влияния распространяются
на все физиологические свойства:
возбудимость, проводимость, сократимость
и автоматию.
1) Изменение автоматии отражается в
изменение частоты – хронотропный
эффект.
2) Изменение сократимости в силе сокращения
– инотропный эффект.
3) Изменение возбудимости – батмотропный.
4) Изменение проводимости – дромотропный.
Клеточные механизмы регуляции.
Речь идет о клетках водителях ритма.
Клеточный уровень регуляции обеспечивает
хронотропный эффект – изменение
ЧСС.
Причины, вызывающие изменение
хронотропного эффекта.
1) Смена водителя ритма.
2) Изменение крутизны медленной
диастолической деполяризации.
3) Изменение ПП.
4) Изменение величины КУМП.
Механизм. В основе лежит изменение
скорости диастолической деполяризации.
Механизм клеточного уровня регуляции
сокращения и расслабления (инотропный
эффект).
Регулируемыми показателями являются
сила и скорость сокращения; степень и
скорость расслабления.
Сила и скорость сокращения зависят:
1) от количества актина и миозина;
2) скорости образования акто – миозинового
комплекса;
3) количество Са2+, поступающего
внутрь волокна во время генерации ПД.
Степень и скорость расслабления зависят
от активности Са2+ насоса в
кардиоците.
Интраорганные механизмы. На
интраорганном уровне регулируется сила
сокращений в зависимости → от венозного
возврата (закон Старлинга).
↓ → от ЧСС (закон Боудича).
от сопротивления кровотоку
(закон Анрепа – Хилла).
1) В ответ на увеличение венозного
возврата.
Это гетерометрическая регуляция силы
сокращения – закон Старлинга или закон
сердца.
Увеличение длины миокарда при возрастании
венозного возврата вызывает увеличение
силы сокращения ↑ венозный возврат →
↑ длины миокарда → ↑ силы сокращения
→ ↑ СВ.
Механизм Растяжение мышцы способствует
образованию большого количества акто
– миозиновых мостиков и увеличивает
силу сокращения.
2) В ответ на увеличение сопротивления
кровотоку.
В этом случае венозный возврат не
изменяется, но меняется сопротивление
кровотоку – например, повышается АД в
большом круге кровообращения.
Это гомеометрическая регуляция силы
сокращения (закон Анрепа – Хилла).
Проявление. Повышение АД сопровождается
увеличением силы сокращения.
Следствием этого закона является
развитие гипертрофии миокарда при
стойком повышении АД.
3) В ответ на увеличение частоты СС
(закон Боудича).
Повышение частоты СС до 170 ударов в
минуту сопровождается увеличением силы
сокращения. Это связано с тем, что в
кардиомиоците накапливается Са, который
и увеличивает силу сокращения.
Экстракардиальный уровень.
Подчиняет себе клеточный и органный
(нервный, гуморальный).
Нервные влияния через симпатическую
и парасимпатическую нервную систему.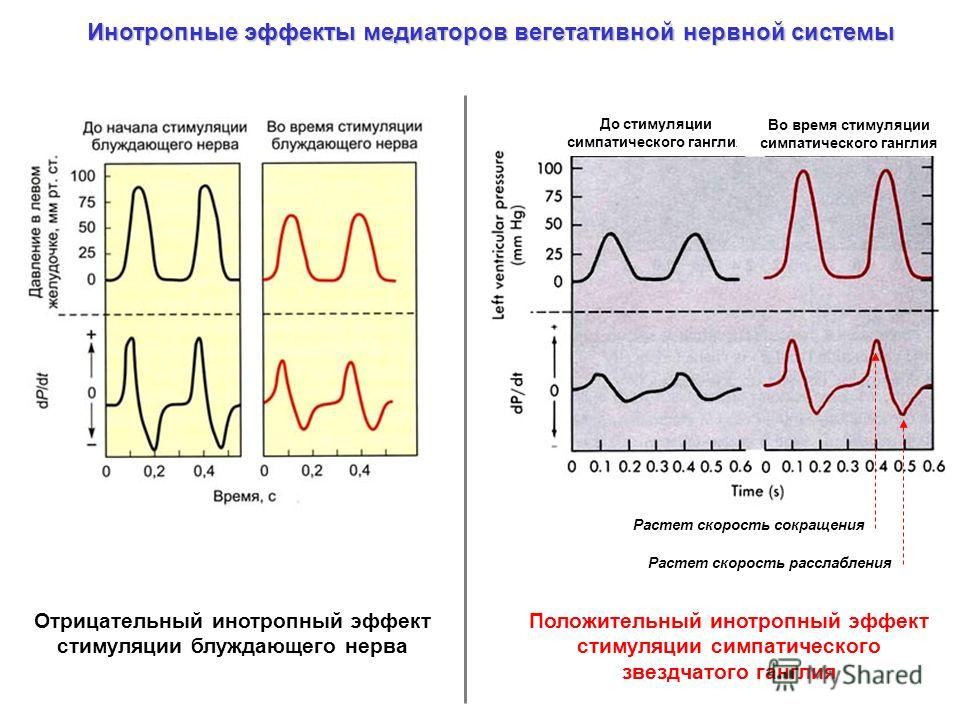
Блуждающий нерв – ядро в продолговатом
мозге. Преганглионарное волокно
прерывается в интрамуральном ганглии
сердца. Постганглионарное волокно
выделяет АХ, рецептор к нему на сердце
– М – ХР.
Раздражение блуждающего нерва вызывает
отрицательные ино – хроно – дромо и
батмотропные эффекты, т. е. тормозящий
эффект.
Механизм – снижение скорости
медленной диастолической деполяризации
в синоатриальном узле, гиперполяризация.
Эффект ускользания.
Ядра блуждающего нерва обладают тонусом.
У новорожденных этот тонус не выражен.
Колебания тонуса блуждающего нерва
проявляются в виде тахикардии и
брадикардии, дыхательно – сердечной
аритмии.
Поддержание тонуса блуждающего нерва
осуществляется афферентными импульсами
с дуги аорты и каротидных синусов. При
его перерезке ЧСС увеличивается.
Симпатическая иннервация осуществляется
из 5 верхних грудных сегментов. Иннервирует
Иннервирует
желудочки. Преганглионарные волокна
прерываются в звездчатом ганглии,
постганглионарные выделяют норадреналин,
β — адренорецепторы.
Влияние – возбуждающее, положительные
ино – хроно – дромо эффекты. Влияние
осуществляется при поступлении к
симпатическим центрам сигналов из
надсегментарного отдела и с периферии.
Симпатическая система оказывает
адаптационно – трофическое влияние,
т. е. обеспечивает приспособление ССС
к возросшим нагрузкам (физическим,
психическим, эмоциональным).
Рефлекторные влияния на сердце.
Различают вагальные и симпатические
рефлексы.
Вагальные рефлексы связаны с
повышением тонуса ядра блуждающего
нерва и усилением его тормозящего
влияния на сердце при раздражении
различных рефлексогенных зон.
Локализация зон.
1) В сердечно – сосудистой системе.
Пример: ↑ АД → барорецепторы дуги аорты,
легочной артерии, сосудов внутренних
органов, эндо – мио – и перикарда →
повышение тонуса Х пары → замедление
сокращений сердца.
2) За пределами ССС.
а) С рецепторов желудочков и кишечника
– рефлекс Гольца. При ударе по животу
урежение сердцебиений до остановки
сердца (чревный нерв → блуждающий нерв
→ ↑ частота сердцебиений).
б) При надавливании на глазное яблоко
– рефлекс Ашнера (глазо-сердечный).
в) Повышение тонуса блуждающего нерва
наблюдается при выдохе, проявляется в
виде дыхательно – сердечной аритмии.
Симпатические рефлексы связаны
со снижением тормозного влияния
блуждающего нерва и усиления тонуса
симпатических центров.
1) С рефлексогенных зон ССС. Например:
повышение давления в устье полых вен
при большом венозном возврате возбуждает
рецепторы устья полых вен и правого
сердца → повышение симпатических
влияний на сердце → увеличение ЧСС
(разгрузочный рефлекс Бейнбриджа).
2) ↓ АД → барорецепторы сосудистой
системы → ↓ тонуса блуждающего
нерва→↑ЧСС.
3) С хеморецепторов ССС.
С других рецепторов.
1) С болевых.
2) С тепловых терморецепторов.
3) При эмоциональных состояниях.
Условнорефлекторная регуляция работы
сердца.
Пример: предстартовое учащение
сердцебиений (на 22 35 удара).
Роль различных отделов ЦНС:
1) кора;
2) гипоталямус;
3) лимбикоретикулярный комплекс;
3) продолговатый мозг;
4) спинной мозг.
Гуморальная регуляция работы сердца.
Осуществляется веществами, переносимыми
кровью. Различают 1) непосредственное
и 2) опосредованное действие.
Непосредственное действие.
1) Гормоны: катехоламины увеличивают
частоту сокращений, активируют β –
адренорецепторы → АИ → АТФ → ц АМФ →
фосфорилаза → расщепление гликогена
→ ↑ сокращение.
↓
Са2+ —
увеличивает сопряжение возбуждения и
сокращения.
Катехоламины, кроме того увеличивают
проницаемость для Са2+ — клеточных
мембран.
Хронотропное действие уменьшает
время медленной деполяризации.
2) Глюкогон действует непосредственно
через симпатоадреналовую систему.
3) Глюкокортикоиды – увеличивают
силу сердечных сокращений.
4) Тироксин – увеличивает частоту.
Электролиты.
Са2+ увеличивает силу сокращений.
Передозировка – остановка в систолу.
К+ — на возбудимость, передозировка
остановка в диастолу.
Опосредованное влияние осуществляется
через нервные центры.
Н+ — повышает влияние симпатической
системы. АХ — ↑ тонус блуждающего нерва.
АХ — ↑ тонус блуждающего нерва.
3
★ Хронотропный эффект — cтатьи по кардиологии .. Информация
Пользователи также искали:
батмотропный эффект,
отрицательный хронотропный эффект препараты,
отрицательный хронотропный эффект сердечных гликозидов,
препараты с отрицательным хронотропным эффектом,
эффектом,
Хронотропный,
хронотропный,
эффект,
препараты,
отрицательный,
Хронотропный эффект,
хронотропным,
отрицательным,
гликозидов,
сердечных,
батмотропный,
батмотропный эффект,
препараты с отрицательным хронотропным эффектом,
отрицательный хронотропный эффект сердечных гликозидов,
отрицательный хронотропный эффект препараты,
хронотропный эффект,
cтатьи по кардиологии. хронотропный эффект,
хронотропный эффект,
Анаприлин инструкция по применению: показания, противопоказания, побочное действие – описание Anaprilin таб. 10 мг: 50 шт. (18927)
При одновременном применении с гипогликемическими средствами возникает риск развития гипогликемии за счет усиления действия гипогликемических средств.
При одновременном применении с ингибиторами МАО существует вероятность развития нежелательных проявлений лекарственного взаимодействия.
Описаны случаи развития выраженной брадикардии при применении пропранолола по поводу аритмии, вызванной препаратами дигиталиса.
При одновременном применении со средствами для ингаляционного наркоза повышается риск угнетения функции миокарда и развития артериальной гипотензии.
При одновременном применении с амиодароном возможны артериальная гипотензия, брадикардия, фибрилляция желудочков, асистолия.
При одновременном применении с верапамилом возможны артериальная гипотензия, брадикардия, диспноэ. Повышается Cmax в плазме крови, увеличивается AUC, уменьшается клиренс пропранолола вследствие угнетения его метаболизма в печени под влиянием верапамила.
Повышается Cmax в плазме крови, увеличивается AUC, уменьшается клиренс пропранолола вследствие угнетения его метаболизма в печени под влиянием верапамила.
Пропранолол не влияет на фармакокинетику верапамила.
Описан случай развития тяжелой артериальной гипотензии и остановки сердца при одновременном применении с галоперидолом.
При одновременном применении с гидралазином повышается Cmax в плазме крови и AUC пропранолола. Полагают, что гидралазин может уменьшать печеночный кровоток или ингибировать активность печеночных ферментов, что приводит к замедлению метаболизма пропранолола.
При одновременном применении пропранолол может ингибировать эффекты глибенкламида, глибурида, хлорпропамида, толбутамида, т.к. неселективные бета2-адреноблокаторы способны блокировать β2-адренорецепторы поджелудочной железы, связанные с секрецией инсулина.
Обусловленный действием производных сульфонилмочевины выброс инсулина из поджелудочной железы ингибируется бета-адреноблокаторами, что в некоторой степени препятствует развитию гипогликемического эффекта.
При одновременном применении с дилтиаземом повышается концентрация пропранолола в плазме крови вследствие ингибирования его метаболизма под влиянием дилтиазема. Наблюдается аддитивное угнетающее влияние на деятельность сердца в связи с замедлением проведения импульса через AV-узел, вызываемого дилтиаземом. Возникает риск развития выраженной брадикардии, значительно уменьшается ударный и минутный объем.
При одновременном применении описаны случаи повышения концентрации варфарина и фениндиона в плазме крови.
При одновременном применении с доксорубицином в экспериментальных исследованиях показано усиление кардиотоксичности.
При одновременном применении пропранолол препятствует развитию бронходилатирующего эффекта изопреналина, сальбутамола, тербуталина.
При одновременном применении описаны случаи повышения концентрации имипрамина в плазме крови.
При одновременном применении с индометацином, напроксеном, пироксикамом, ацетилсалициловой кислотой возможно уменьшение антигипертензивного действия пропранолола.
При одновременном применении с кетансерином возможно развитие аддитивного гипотензивного эффекта.
При одновременном применении с клонидином усиливается антигипертензивное действие.
У пациентов, получающих пропранолол, в случае резкой отмены клонидина возможно развитие тяжелой артериальной гипертензии. Полагают, что это связано с повышением содержания катехоламинов в циркулирующей крови и усилением их вазоконстрикторного действия.
При одновременном применении с кофеином возможно уменьшение эффективности пропранолола.
При одновременном применении возможно усиление эффектов лидокаина и бупивакаина (в т.ч. токсических), по-видимому, вследствие замедления метаболизма местных анестетиков в печени.
При одновременном применении с лития карбонатом описан случай развития брадикардии.
При одновременном применении описан случай усиления побочного действия мапротилина, что обусловлено, по-видимому, замедлением его метаболизма в печени и кумуляцией в организме.
При одновременном применении с мефлохином увеличивается интервал QT, описан случай остановки сердца; с морфином — усиливается угнетающее действие на ЦНС, вызываемое морфином; с натрия амидотризоатом — описаны случаи тяжелой артериальной гипотензии.
При одновременном применении с низолдипином возможны повышение Cmax и AUC пропранолола и низолдипина в плазме крови, что приводит к тяжелой артериальной гипотензии. Имеется сообщение об усилении бета-блокирующего действия.
Описаны случаи повышения Cmax и AUC пропранолола, артериальной гипотензии и уменьшения ЧСС при одновременном применении с никардипином.
При одновременном применении с нифедипином у пациентов с ИБС возможно развитие выраженной артериальной гипотензии, повышение риска развития сердечной недостаточности и инфаркта миокарда, что может быть обусловлено усилением отрицательного инотропного действия нифедипина.
У пациентов, получающих пропранолол, имеется риск развития выраженной артериальной гипотензии после приема первой дозы празозина.
При одновременном применении с прениламином увеличивается интервал QT.
При одновременном применении с пропафеноном повышается концентрация пропранолола в плазме крови и развивается токсическое действие. Полагают, что пропафенон ингибирует метаболизм пропранолола в печени, уменьшая его клиренс и повышая сывороточные концентрации.
При одновременном применении резерпина, других антигипертензивных средств повышается риск развития артериальной гипотензии и брадикардии.
При одновременном применении повышается Cmax и AUC ризатриптана; с рифампицином — уменьшается концентрация пропранолола в плазме крови; с суксаметония хлоридом, тубокурарина хлоридом — возможно изменение действия миорелаксантов.
При одновременном применении уменьшается клиренс теофиллина вследствие замедления его метаболизма в печени. Возникает риск развития бронхоспазма у пациентов с бронхиальной астмой или с ХОБЛ. Бета-адреноблокаторы могут блокировать инотропный эффект теофиллина.
При одновременном применении с фениндионом описаны случаи некоторого повышения кровоточивости без изменений показателей свертываемости крови.
При одновременном применении с флекаинидом возможно аддитивное кардиодепрессивное действие.
Флуоксетин ингибирует изофермент CYP2D6, что приводит к угнетению метаболизма пропранолола и его кумуляции и может усиливать кардиодепрессивное действие (в т.ч. брадикардию). Флуоксетин и, главным образом, его метаболиты характеризуются длительным T1/2, поэтому вероятность лекарственного взаимодействия сохраняется даже через несколько дней после отмены флуоксетина.
Хинидин ингибирует изофермент CYP2D6, что приводит к угнетению метаболизма пропранолола, при этом уменьшается его клиренс. Возможны усиление бета-адреноблокирующего действия, ортостатическая гипотензия.
При одновременном применении в плазме крови повышаются концентрации пропранолола, хлорпромазина, тиоридазина. Возможно резкое уменьшение АД.
Циметидин ингибирует активность микросомальных ферментов печени (в т.ч. изофермента CYP2D6), это приводит к угнетению метаболизма пропранолола и его кумуляции: наблюдается усиление отрицательного инотропного действия и развитие кардиодепрессивного действия.
При одновременном применении усиливается гипертензивное действие эпинефрина, возникает риск развития тяжелых угрожающих жизни гипертензивных реакций и брадикардии. Уменьшается бронхорасширяющее действие симпатомиметиков (эпинефрина, эфедрина).
При одновременном применении описаны случаи уменьшения эффективности эрготамина.
Имеются сообщения об изменении гемодинамических эффектов пропранолола при одновременном применении с этанолом.
РЕГУЛЯЦИЯ СЕРДЕЧНОЙ ДЕЯТЕЛЬНОСТИ. КРОВООБРАЩЕНИЕ В МИОКАРДЕ — Мегаобучалка
15–1. Хронотропный эффект в деятельности сердца – это изменение:
1 – проводимости миокарда
2 – силы сокращений
3 – возбудимости миокарда
4 – частоты сердечных сокращений
5 – тонуса миокарда
15–2. Инотропный эффект в деятельности сердца – это изменение:
1 – проводимости миокарда
2 – силы сокращений
3 – возбудимости миокарда
4 – частоты сердечных сокращений
5 – тонуса миокарда
15–3. Батмотропный эффект в деятельности сердца – это изменение:
Батмотропный эффект в деятельности сердца – это изменение:
1 – проводимости миокарда
2 – силы сокращений
3 – возбудимости миокарда
4 – частоты сердечных сокращений
5 – тонуса миокарда
15–4. Дромотропный эффект в деятельности сердца – это изменение:
1 – проводимости миокарда
2 – силы сокращений
3 – возбудимости миокарда
4 – частоты сердечных сокращений
5 – тонуса миокарда
15–6. Физиологический смысл закона сердца (Старлинга):
1 – увеличение силы сокращения сердца при увеличении объема притекающей к нему крови
2 – увеличение силы сокращения сердца при увеличении давления в аорте и легочной артерии
3 – увеличение силы сокращения сердца при увеличении частоты сердечных сокращений
4 – увеличение силы сокращения сердца при снижении артериального давления
5 – увеличение силы сокращения сердца при снижении частоты сердечных сокращений
15–7. Эффект Анрепа заключается в:
Эффект Анрепа заключается в:
1 – изменении силы сокращений сердца при изменении исходной длины мышечных волокон в диастоле
2 – уменьшении частоты сердечных сокращений при надавливании на глазные яблоки
3 – увеличении силы сокращения сердца при повышении давления в артериальной системе
4 – увеличении частоты сердечных сокращений при надавливании на глазные яблоки
5 – увеличении силы сокращений сердца при ударе по передней брюшной стенке
15–9. Пересаженное сердце у реципиента выведено из-под контроля:
1 – периферических рефлексов метасимпатической нервной системы сердца
2 – эндокринной системы реципиента
3 – непосредственного эфферентного влияния ЦНС
4 – опосредованного влияния ЦНС (через эндокринную систему)
5 – нервного влияния с проприоцепторов скелетных мышц
15–10. Центр парасимпатической иннервации сердца находится в:
1 – верхних шейных сегментах спинного мозга
2 – верхних грудных сегментах спинного мозга
3 – продолговатом мозге
4 – таламусе
5 – боковых рогах торако-люмбального отдела спинного мозга
15–11.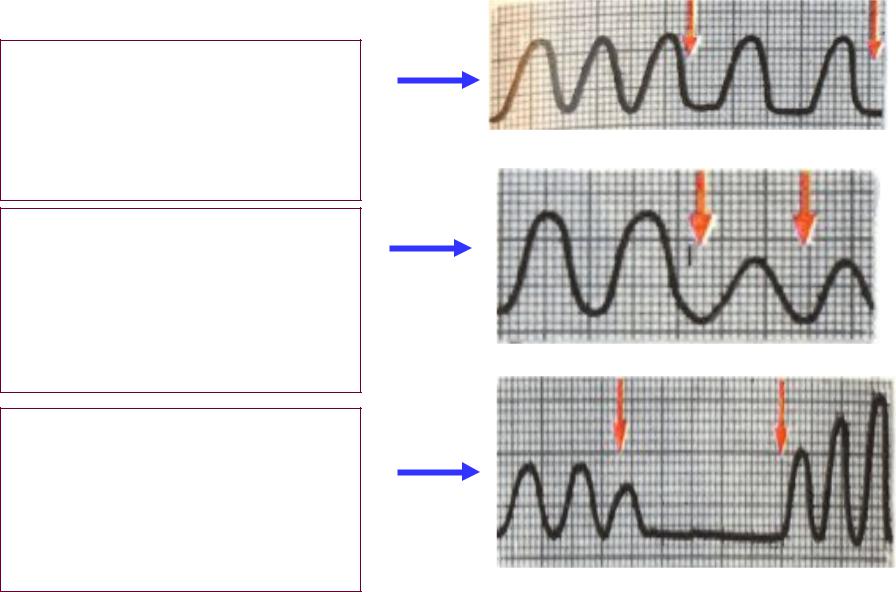 В окончаниях блуждающего нерва, иннервирующего сердце, как правило, выделяется:
В окончаниях блуждающего нерва, иннервирующего сердце, как правило, выделяется:
1 – адреналин
2 – серотонин
3 – ацетилхолин
4 – ГАМК
5 – глицин
15–12. Блуждающий нерв оказывает на сердце:
1 – отрицательные хроно–, ино–, батмо– и дромотропный эффекты
2 – отрицательные хроно–, ино–, батмотропный и положительный дромотропный эффекты
3 – отрицательные хроно–, инотропный и положительные батмо– и дромотропный эффекты
4 – положительные хроно–, ино–, батмо– и дромотропный эффекты
5 – не оказывает никакого влияния
15–13. Блуждающий нерв действует на сердце преимущественно через:
1 – альфа–адренорецепторы
2 – бета–адренорецепторы
3 –пуриновые рецепторы
4 – М–холинорецепторы
5 – серотонинорецепторы
15–14. Механизм отрицательного хронотропного действия вагуса на сердце связан:
1 – с уменьшением скорости медленной диастолической деполяризации
2 – с увеличением скорости медленной диастолической деполяризации
3 – с увеличением кальциевого тока
4 – со снижением калиевого тока
15–15. Центр симпатической иннервации сердца находится в:
Центр симпатической иннервации сердца находится в:
1 – верхних шейных сегментах спинного мозга
2 – продолговатом мозге
3 – верхних грудных сегментах спинного мозга (Th2 – 5)
4 – боковых рогах торако–люмбального отдела спинного мозга
5 – таламусе
15–16. Окончания волокон симпатического нерва, иннервирующего сердце, выделяют:
1 – ацетилхолин
2 – дофамин
3 – норадреналин
4 – ГАМК
5 – глицин
15–17. Симпатические нервы вызывают в сердце эффекты:
1 – отрицательные хроно–, ино–, батмо– и дромотропный эффекты
2 – отрицательные хроно–, ино–, батмотропный и положительный дромотропный эффекты
3 – отрицательные хроно–, инотропный и положительные батмо– и дромотропный эффекты
4 – положительные хроно–, ино–, батмо– и дромотропный эффекты
5 – не вызывают никаких эффектов в сердце
15–18. Механизм положительного хронотропного влияния симпатической иннервации на сердце связан:
1 – с увеличением скорости медленной диастолической деполяризации
2 – с уменьшением скорости медленной диастолической деполяризации
3 – с увеличением калиевого тока
4 – со снижением кальциевого тока
15–19. Рефлекс Данини–Ашнера заключается в:
Рефлекс Данини–Ашнера заключается в:
1 – изменении силы сокращения сердца при изменении исходной длины мышечных волокон
2 – изменении силы сокращения сердца при изменении давления в артериальной системе
3 – уменьшении частоты сердечных сокращений при надавливании на глазные яблоки
4 – увеличении частоты сердечных сокращений при надавливании на глазные яблоки
5 – увеличении силы сердечных сокращений при надавливании на глазные яблоки
15–20. Адреналин оказывает на сердце:
1 – положительное хроно–, ино–, батмо– и дромотропное действие
2 – отрицательное хроно–, ино–, батмо– и дромотропное действие
3 – положительное хроно–, инотропное действие, отрицательное батмо- и дромотропное действие
4 – отрицательное хроно–, инотропное действие
15–21. Тироксин, действуя на сердце:
1 – увеличивает ритм, силу сокращений и возбудимость сердца
2 – снижает ритм, силу сокращений и возбудимость сердца
3 – уменьшает ритм и силу сокращений, увеличивает возбудимость сердца
4 – не изменяет функции сердца
15–23. Кровоснабжение миокарда левого желудочка осуществляется:
Кровоснабжение миокарда левого желудочка осуществляется:
1 – преимущественно во время систолы
2 – практически одинаково во время систолы и диастолы
3 – преимущественно во время диастолы
4 – в период изометрического напряжения
РЕГУЛЯЦИЯ РАБОТЫ СЕРДЦА — Студопедия
Выберите один правильный ответ.
565. СИНХРОННОЕ СОКРАЩЕНИЕ КАРДИОМИОЦИТОВ ОБЕСПЕЧИВАЕТСЯ
1) внутрисердечным периферическим рефлексом
2) внутриклеточной регуляцией
3) межклеточным взаимодействием
566. УСИЛЕНИЕ СОКРАЩЕНИЯ ЛЕВОГО ЖЕЛУДОЧКА ПРИ РАСТЯЖЕНИИ СТЕНОК ПРАВОГО ОБЕСПЕЧИВАЕТСЯ
1) внутриклеточной регуляцией
2) межклеточным взаимодействием
3) внутрисердечным периферическим рефлексом
567. УСИЛЕНИЕ СОКРАЩЕНИЯ МИОКАРДА ПРИ УВЕЛИЧЕНИИ ИСХОДНОЙ ДЛИННЫ МЫШЕЧНЫХ ВОЛОКОН ОБЕСПЕЧИВАЕТСЯ
1) внутрисердечным периферическим рефлексом
2) межклеточным взаимодействием
3)внутриклеточной регуляцией
568. ПРИ РАЗДРАЖЕНИИ БЛУЖДАЮЩЕГО НЕРВА СОДЕРЖАНИЕ В СЕРДЕЧНОЙ МЫШЦЕ ИОНОВ КАЛИЯ
ПРИ РАЗДРАЖЕНИИ БЛУЖДАЮЩЕГО НЕРВА СОДЕРЖАНИЕ В СЕРДЕЧНОЙ МЫШЦЕ ИОНОВ КАЛИЯ
1) уменьшается
2) не изменяется
3) в начальную фазу увеличивается, затем уменьшается
4) увеличивается
569. БАТМОТРОПНЫЙ ЭФФЕКТ В ДЕЯТЕЛЬНОСТИ СЕРДЦА — ЭТО ИЗМЕНЕНИЕ
1) ЧСС
2) проводимости миокарда
3) силы сокращений
4) возбудимости миокарда
570. ИНОТРОПНЫЙ ЭФФЕКТ В ДЕЯТЕЛЬНОСТИ СЕРДЦА — ЭТО ИЗМЕНЕНИЕ
1) проводимости миокарда
2) ЧСС
3) возбудимости миокарда
4) силы сокращений
571. ДРОМОТРОПНЫЙ ЭФФЕКТ В ДЕЯТЕЛЬНОСТИ СЕРДЦА — ЭТО ИЗМЕНЕНИЕ
1) силы сокращений
2) возбудимости миокарда
3) ЧСС
4) проводимости миокарда
572. ХРОНОТРОПНЫЙ ЭФФЕКТ В ДЕЯТЕЛЬНОСТИ СЕРДЦА — ЭТО ИЗМЕНЕНИЕ
1) проводимости миокарда
2) силы сокращений
3) возбудимости миокарда
4) ЧСС
573. СИМПАТИЧЕСКИЕ НЕРВЫ ОКАЗЫВАЮТ НА СЕРДЕЧНУЮ МЫШЦЫ ЭФФЕКТЫ
СИМПАТИЧЕСКИЕ НЕРВЫ ОКАЗЫВАЮТ НА СЕРДЕЧНУЮ МЫШЦЫ ЭФФЕКТЫ
1) положительный инотропный, отрицательный хронотропный
2) отрицательный инотропный, положительный хронотропный
3) отрицательный инотропный, отрицательный хронотропный
4) положительный инотропный, положительный хронотропный
574. В ОКОНЧАНИЯХ СИМПАТИЧЕСКОГО НЕРВА, ИННЕРВИРУЮЩЕГО СЕРДЦЕ, ВЫДЕЛЯЕТСЯ МЕДИАТОР
1) ацетилхолин
2) серотонин
3) норадреналин
575. В ОКОНЧАНИЯХ БЛУЖДАЮЩЕГО НЕРВА ВЫДЕЛЯЕТСЯ МЕДИАТОР
1) норадреналин
2) серотонин
3) ацетилхолин
576. ПРИ АПЛИКАЦИИ АЦЕТИЛХОЛИНА НА СЕРДЕЧНУЮ МЫШЦУ ПРОИЗОЙДЕТ
1) деполяризация миоцитов
2) блокада натриевых каналов
3) активация натриевых каналов
4) гиперполяризация миоцитов
577. ДЫХАТЕЛЬНАЯ АРИТМИЯ ПРОЯВЛЯЕТСЯ
1) в увеличении ЧСС к концу выдоха
2) в учащении дыхания при аритмии
3) в уменьшении ЧСС к концу выдоха
578. ЦЕНТР СИМПАТИЧЕСКОЙ ИНЕРВАЦИИ СЕРДЦА НАХОДИТСЯ
ЦЕНТР СИМПАТИЧЕСКОЙ ИНЕРВАЦИИ СЕРДЦА НАХОДИТСЯ
1) в верхних шейных сегментах спинного мозга
2) в продолговатом мозге
3) в верхних грудных сегментах спинного мозга
579. ЦЕНТР ПАРАСИМПАТИЧЕСКОЙ ИНЕРВАЦИИ СЕРДЦА НАХОДИТСЯ
1) в верхних шейных сегментах спинного мозга
2) в верхних грудных сегментах спинного мозга
3) в продолговатом мозге
580. ГОМЕОМЕТРИЧЕСКИЙ МЕХАНИЗМ РЕГУЛЯЦИИ РАБОТЫ СЕРДЦА ЗАКЛЮЧАЕТСЯ В ИЗМЕНЕНИИ
1) частоты сердечных сокращений при изменении давления в артериальной системе
2) силы сокращений сердца при изменении исходной длины мышечных волокон
3) силы сокращений сердца при изменении давления в артериальной системе
581. ГЕТЕРОМЕТРИЧЕСКИЙ МЕХАНИЗМ РЕГУЛЯЦИИ РАБОТЫ СЕРДЦА ЗАКЛЮЧАЕТСЯ В ИЗМЕНЕНИИ
1) силы сокращений сердца при изменении давления в артериальной системе
2) частоты сердечных сокращений при изменении исходной длины мышечных волокон
3) силы сокращений сердца при изменении исходной длины мышечных волокон
582. РЕФЛЕКС ГОЛЬЦА — ЭТО
РЕФЛЕКС ГОЛЬЦА — ЭТО
1) изменение силы сокращений сердца при изменении исходной длины мышечных волокон
2) изменение силы сокращений сердца при изменении давления в артериальной системе
3) рефлекторная остановка сердца при ударе в эпигастральную область
583. РЕФЛЕКС АШНЕРА ЗАКЛЮЧАЕТСЯ
1) в изменении силы сокращений сердца при изменении исходной длины мышечных волокон
2) в изменении силы сокращений сердца при изменении давления в артериальной системе
3) в уменьшении ЧСС при надавливании на глазные яблоки
584. ЭФФЕКТ АНРЕПА ЗАКЛЮЧАЕТСЯ
1) в изменении силы сокращений сердца при изменении исходной длины мышечных волокон
2) в уменьшении ЧСС при надавливании на глазные яблоки
3) в изменении силы сокращения сердца при изменении давления в артериальной системе
585. ЧАСТОТА СЕРДЕЧНЫХ СОКРАЩЕНИЙ МОЖЕТ ИЗМЕНЯТСЯ УСЛОВНОРЕФЛЕКТОРНО
1) нет
2) да
586. РОЛЬ ГИПОТАЛАМУСА В РЕГУЛЯЦИИ РАБОТЫ СЕРДЦА ЗАКЛЮЧАЕТСЯ
РОЛЬ ГИПОТАЛАМУСА В РЕГУЛЯЦИИ РАБОТЫ СЕРДЦА ЗАКЛЮЧАЕТСЯ
1) в условнорефлекторном изменении ЧСС
2) в изменении ЧСС при задерке дыхания
3) в обеспечении работы сердца, адекватной ситуации и поведению
Установите соответствие.
587.
РЕГУЛЯТОРНЫЕ ВЛИЯНИЯ НА ДЕЯТЕЛЬНОСТЬ СЕРДЦА…. ПРОЯВЛЯЮТСЯ
А.2 Инотропное 1. В увеличении ЧСС.
Б.1 Хронотропное 2. В увеличении силы сокращения миокарда.
В.4 Батмотропное 3. В увеличении проводимости.
Г.3 Дромотропное 4. В увеличении возбудимости.
588.
ЭФФЕКТ…. ЯВЛЯЕТСЯ ПРИМЕРОМ РЕГУЛЯЦИИ РАБОТЫ СЕРДЦА
А.2 Франка — Старлинга 1. Нейрогенной интракардиальной.
Б.3 Анрепа 2. Миогенной гетерометрической.
В.1 Рефлекса Бэйнбриджа 3. Миогенной гомеометрической.
Г.1 Тахикардии в условиях 4. Гуморальной.
гипоксемии
589.
МЕДИАТОРЫ ПОСТГАНГЛИОНАРНЫХ ОКОНЧАНИЙ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ ВЗАИМОДЕЙСТВУЮТ С РЕЦЕПТОРАМИ
КАРДИОМИОЦИТОВ
А. 1 Ацетилхолин 1. Мускариновыми холинорецепторами.
1 Ацетилхолин 1. Мускариновыми холинорецепторами.
Б.4 Норадреналин 2. Никотиновыми холинорецепторами.
3. Альфа-адренорецепторами.
4. Бета-адренорецепторами.
590.
МЕДИАТОР… ИЗМЕНЯЕТ ПРОНИЦАЕМОСТЬ МЕМБРАНЫ КАРДИОМИОЦИТА ДЛЯ ИОНОВ
А.1 Ацетилхолин 1. Увеличивает для калия.
Б.3 Норадреналин 2. Снижает для калия.
3. Снижает для калия, увеличивает для Nа и Са.
4. Снижает для Nа и Са.
591.
ХРОНОТРОПНЫЙ ЭФФЕКТ В ДЕЯТЕЛЬНОСТИ СЕРДЦА…НАБЛЮДАЕТСЯ
А.135 Положительный 1. При гипоксемии.
Б.24 Отрицательный 2. При гипероксии.
3. При увеличении кровенаполнения в полых венах.
4. При раздражении механорецепторов органов брюшной полости.
5. При снижении давления в аорте.
592.
ВОЗБУЖДЕНИЕ ВЕГЕТАТИВНЫХ НЕРВОВ…. ОКАЗЫВАЕТ ВЛИЯНИЕ НА СЕРДЕЧНУЮ ДЕЯТЕЛЬНОСТЬ
А.135 Симпатических 1. Положительное инотропное.
Б.246 Парасимпатических 2. Отрицательное инотропное.
3. Положительное хронотропное.
4. Отрицательное хронотропное.
5. Положительное дромотропное.
6. Отрицательное дромотропное.
Определите верны или неверны утверждения и связь между ними.
593. Постганглионарные парасимпатические волокна оказывают отрицательный инотропный эффект на сердечную мышцу, потому что их окончания выделяют ацетилхолин.
1) ВВН
2) ВНН
3) НВН
4) ННН
5) ВВВ
594. Постганглионарные симпатические волокна оказывают положительный инотропный эффект на сердечную мышцу, потому что их окончания выделяют норадреналин.
1) ВВН
2) ВНН
3) НВН
4) ННН
5) ВВВ
595. Частота сердечных сокращений может изменяться условнорефлекторно, потому что в регуляции сердечной деятельности участвует гипоталамус.
1) ВНН
2) ВВВ
3) НВН
4) ННН
5) ВВН
596. ЧСС у спортсмена на старте увеличивается, потому что регуляция сердечной деятельности осуществляется и условно-рефлекторным путем.
1) ВВН
2) ВНН
3) НВН
4) ННН
5) ВВВ
597. Частота сердечных сокращений возрастает при эмоциональном напряжении, потому что в регуляции деятельности сердца участвует гипоталамус.
1) ВВН
2) ВНН
3) НВН
4) ННН
5) ВВВ
ПЕРИФЕРИЧЕСКОЕ КРОВООБРАЩЕНИЕ
Выберите один правильный ответ.
598. К ЕМКОСТНЫМ СОСУДАМ ОТНОСЯТСЯ
1) аорта
2) крупные артерии
3) капилляры
4) вены
599. ОСНОВНЫМ ЗВЕНОМ В СИСТЕМЕ МИКРОЦИРКУЛЯЦИИ ЯВЛЯЕТСЯ
1) вены и венулы
2) артериолы
3) крупные артерии
4) капилляры
600. РЕЗИСТИВНЫМИ СОСУДАМИ НАЗЫВАЮТ
1) аорту
2) вены и венулы
3) мелкие артерии и артериолы
601. СОСУДАМИ КОМПРЕССИОННОЙ КАМЕРЫ (КОТЛА) НАЗЫВАЮТ
1) артерии и вены
2) капилляры
3) аорту
4) крупные эластические и мышечные сосуды
602. ЛИНЕЙНАЯ СКОРОСТЬ КРОВОТОКА В АОРТЕ РАВНА
ЛИНЕЙНАЯ СКОРОСТЬ КРОВОТОКА В АОРТЕ РАВНА
1) 0.5 см/c
2) 25 cм/c
3) 50 см/c
603. ЛИНЕЙНАЯ СКОРОСТЬ КРОВОТОКА В КАПИЛЛЯРАХ РАВНА
1) 50 мм/c
2) 25 мм/c
3) 0.5 мм/c
604. ВРЕМЯ ПОЛНОГО ОБОРОТА КРОВИ ПО СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЕ РАВНО
1) 1.5-2 мин
2) 40-45 c
3) 20-23 c
605. КРОВЯНОЕ ДАВЛЕНИЕ В КАПИЛЛЯРАХ БОЛЬШОГО КРУГА РАВНО
1) 80-70 мм рт. ст.
2) 5-3 мм рт. ст.
3) 40-10 мм рт. ст.
606. ЛИНЕЙНАЯ СКОРОСТЬ КРОВОТОКА МЕНЯЕТСЯ ПО ХОДУ СОСУДИСТОГО РУСЛА
1) нет
2) да
607. ОБЪЕМНАЯ СКОРОСТЬ КРОВОТОКА МЕНЯЕТСЯ ПО ХОДУ СОСУДИСТОГО РУСЛА
1) да
2) нет
608. СОСУДОДВИГАТЕЛЬНЫЙ ЦЕНТР РАСПОЛОЖЕН
1) в спинном мозге
2) в варолиевом мосту
3) в продолговатом мозге
609. ПРОСВЕТ СОСУДОВ УВЕЛИЧИВАЕТСЯ ПОД ДЕЙСТВИЕМ
1) вазопрессина
2) серотонина
3) ацетилхолина
610. ОКОНЧАТЫЕ КАПИЛЛЯРЫ РАСПОЛАГАЮТСЯ
ОКОНЧАТЫЕ КАПИЛЛЯРЫ РАСПОЛАГАЮТСЯ
1) в печени, костном мозге
2) в мышцах, легких,жировой и соединительной тканях
3) в почках, железах внутренней секреции
611. СПЛОШНЫЕ КАПИЛЛЯРЫ РАСПОЛАГАЮТСЯ
1) в печени, костном мозге
2) в почках, железах внутренней секреции
3) в мышцах, легких, жировой и соединительной тканях
612. НЕСПЛОШНЫЕ КАПИЛЛЯРЫ РАСПОЛАГАЮТСЯ
1) в мышцах, легких, жировой и соеденительной тканях
2) в почках, железах внутренней секреции
3) в печени, костном мозге
613. РАЗДРАЖЕНИЕ БАРОРЕЦЕПТОРОВ АОРТЫ И СОННОЙ АРТЕРИИ ВЫЗЫВАЕТ РЕФЛЕКСЫ
1) депрессорные
2) прессорные
614. РАЗДРАЖЕНИЕ МЕХАНОРЕЦЕПТОРОВ БИФУРКАЦИИ ЛЕГОЧНОЙ АРТЕРИИ ВЫЗЫВАЕТ РЕФЛЕКСЫ
1) прессорные
2) депрессорные
615. КОРОНАРНЫЙ КРОВОТОК МАКСИМАЛЕН
1) в систолу предсердий
2) в систолу желудочков
3) в общую паузу
616. МИКРОПИНОЦИТОЗ ЯВЛЯЕТСЯ ОДНИМ ИЗ ВИДОВ ТРАНСКАПИЛЛЯРНОГО ОБМЕНА
МИКРОПИНОЦИТОЗ ЯВЛЯЕТСЯ ОДНИМ ИЗ ВИДОВ ТРАНСКАПИЛЛЯРНОГО ОБМЕНА
1) нет
2) да
617. МИКРОПИНОЦИТОЗ ЯВЛЯЕТСЯ ОДНИМ ИЗ ВИДОВ ПАССИВНОГО ТРАНСКАПИЛЛЯРНОГО ПРЕНОСА
1) да
2) нет
Дополните утверждение.
618. БАЗАЛЬНЫЙ ТОНУС СОСУДОВ — ЭТО ТОНУС, ОБУСЛОВЛЕННЫЙ……
1) влиянием парасимпатического отдела ВНС
2) влиянием симпатического отдела ВНС
3) автоматией гладких мышечных клеток сосудистой стенки
619. АДРЕНАЛИН …….ПРОСВЕТ ПЕРИФЕРИЧЕСКИХ СОСУДОВ
1) не изменяет
2) увеличивает
3) уменьшает
620. АДРЕНАЛИН ……..ПРОСВЕТ СОСУДОВ МОЗГА
И КОРОНАРНЫХ СОСУДОВ
1) не изменяет
2) уменьшает
3) увеличивает
621. АЦЕТИЛХОЛИН ……..ПРОСВЕТ СОСУДОВ
1) не изменяет
2) уменьшает
3) увеличивает
622. СЕРОТОНИН . …….ПРОСВЕТ СОСУДОВ
…….ПРОСВЕТ СОСУДОВ
1) увеличивает
2) не изменяет
3) уменьшает
623. ГИСТАМИН ……..ПРОСВЕТ СОСУДОВ
1) не изменяет
2) уменьшает
3) увеличивает
624. ФИЛЬТРАЦИЯ-АБСОРБЦИЯ ЯВЛЯЕТСЯ ………ВИДОМ ТРАНСПОРТА
1) активным
2) пассивным
Установите соответствие.
625.
ИЗМЕНЕНИЕ
ПРОСВЕТА АРТЕРИОЛ… ВЫЗЫВАЕТ ВЕЩЕСТВО
А.25 Сужение 1. Адреналин (при связывании
Б.134 Расширение c бета-адренорецепторами).
2. Норадреналин (при связыании
c альфа-адренорецепторами).
3. Гистамин.
4. Углекислый газ.
5. Ионы Са.
626.
ФЕНОМЕН…. ЗАКЛЮЧАЕТСЯ
А.3 Реактивной гиперемии 1. В увеличении кровотока в органе
Б.1 Рабочей гиперемии при усилении его деятельности.
В.4 Ауторегуляции 2. Увеличении кровотока в органе при снижении его деятельности.
кровотока 3. Увеличении кровотока в органе после временного его прекращения.
Увеличении кровотока в органе после временного его прекращения.
4. Стабилизации кровотока в органах при изменении АД в результате изменения просвета артериол.
5. Увеличении кровотока в венах нижних конечностей в вертикальном положении.
627.
ТИПЫ КАПИЛЛЯРОВ…. ПРОНИЦАЕМЫ ДЛЯ
А.2 Сплошные 1. Макромолекул и форменных элементов крови.
Б.3 Окончатые 2. Воды и растворенных солей.
В.1 Несплошные 3. Воды, растворенных солей, макромолекул.
(синусоидные)
628.
УРОВНИ РЕГУЛЯЦИИ КРОВООБРАЩЕНИЯ…..ВЫПОЛНЯЮТ ФУНКЦИИ
А.2 Спинальный 1. Координации кровообращения с другими
Б.3 Бульбарный висцеральными и соматическими функциями.
В.1 Гипоталамический 2. Передачи нисходящих эфферентных влияний на эффекторные структуры сердечно-сосудистой системы
Г.4 Кортикальный 3. Прямой тонической регуляции кроовобращения, осуществление большинства сердечно-сосудистых рефлексов.
4. Обеспечения сопряженных кардиальных рефлексов при поведенческих реакциях.
629.
КРОВЕНОСНЫЕ СОСУДЫ…. ДЕЛЯТСЯ НА ГРУППЫ
А.2 Вены 1. «Компрессионной камеры».
Б.1 Аорта и крупные артерии 2. Емкостными сосудами.
В.3 Артерии мелкого и среднего 3. Резистивными сосудами.
калибра, артериолы 4. Обменными сосудами.
Г.4 Капилляры
630.
ЦЕНТР……РАСПОЛАГАЕТСЯ….
А.2 Высший центр 1. В ядрах среднего мозга.
симпатической регуляции 2. В боковых рогах шейных сегментов
сердечной деятельности спинного мозга.
Б.1 Высший центр парасим- 3. В оливах продолговатого мозга.
патической регуляции 4. В ядрах моста
сердечной деятельности
В.3 Сосудодвигательный центр
631.
ЧЕРЕЗ КАПИЛЛЯРНУЮ СТЕНКУ ПОСРЕДСТВОМ…. ТРАНСПОРТИРУЕТСЯ ВЕЩЕСТВО
А.3 Пассивной диффузии 1. Крупные белковые молекулы.
Б.2 Активного транспорта 2. Аминокислоты.
В.4 Микропиноцитоза 3. Ионы электролитов.
Г.1 Макропиноцитоза 4. Пептиды.
МЕТОДЫ ИССЛЕДОВАНИЯ ССС
Выберите один правильный ответ.
632. ЭЛЕКТРОДЫ ДЛЯ РЕГИСТРАЦИИ ЭКГ В I СТАНДАРТНОМ ОТВЕДЕНИИ РАСПОЛАГАЮТСЯ ТАК
1) правая рука — левая нога
2) левая рука — левая нога
3) правая рука — левая рука
633. ЭЛЕКТРОДЫ ДЛЯ РЕГИСТРАЦИИ ЭКГ В II СТАНДАРТНОМ ОТВЕДЕНИИ РАСПОЛАГАЮТСЯ ТАК
1) правая рука — левая рука
2) левая рука — левая нога
3) правая рука — левая нога
634. ЭЛЕКТРОДЫ ДЛЯ РЕГИСТРАЦИИ ЭКГ В III СТАНДАРТНОМ ОТВЕДЕНИ РАСПОЛАГАЮТСЯ ТАК
1) правая рука — левая рука
2) правая рука — левая нога
3) левая рука — левая нога
635. ОДНОПОЛЮСНЫМИ ЯВЛЯЮТСЯ
1) стандартные отведения
2) грудные отведения по Вильсону
636. ЗУБЕЦ Р НА ЭЛЕКТРОКАРДИОГРАММЕ ОТРАЖАЕТ
1) возбуждение в желудочках
2) реполяризацию в желудочках
3) возбуждение предсердий
637. КОМПЛЕКС QRS НА ЭЛЕКТРОКАРДИОГРАММЕ ОТРАЖАЕТ
1) возбуждение предсердий
2) реполяризацию желудочков
3) возбуждение желудочков
638. ЗУБЕЦ Т НА ЭЛЕКТРОКАРДИОГРАММЕ ОТРАЖАЕТ
ЗУБЕЦ Т НА ЭЛЕКТРОКАРДИОГРАММЕ ОТРАЖАЕТ
1) возбуждение желудочков
2) возбуждение предсердий
3) реполяризацию желудочков
639. ИНТЕРВАЛ Т-Р НА ЭЛЕКТРОКАРДИОГРАММЕ СООТВЕТСТВУЕТ
1) диастоле желудочков
2) систоле предсердий
3) общей паузе сердца
640. ПО ЭЛЕКТРОКАРДИОГРАММЕ МОЖНО СУДИТЬ
1) о силе сокращений сердца
2) о сердечном выбросе
3) о характере возникновения и распространения возбуждения
по миокарду
641. СУТЬ МЕТОДА ВЕКТОРЭЛЕКТРОКАРДИОГРАФИИ ЗАКЛЮЧАЕТСЯ
1) в регистрации суммарной активности кардиомиоцитов
2) в регистрации вектора ЭДС и электрической оси сердца
642. I ТОН СЕРДЦА ВОЗНИКАЕТ
1) в фазу быстрого пассивного наполнения желудочков
2) при захлопывании полулунных клапанов
3) при захлопывании створчатых клапанов
643. II ТОН СЕРДЦА ВОЗНИКАЕТ
1) в фазу быстрого пассивного наполнения желудочков
2) при захлопывании створчатых клапанов
3) при захлопывании полулунных клапанов
644. III ТОН СЕРДЦА РЕГИСТРИРУЕТСЯ НА ФОНОКАРДИОГРАММЕ
III ТОН СЕРДЦА РЕГИСТРИРУЕТСЯ НА ФОНОКАРДИОГРАММЕ
1) при захлопывании полулунных клапанов
2) при захлопывании створчатых клапанов
3) в фазу быстрого пассивного наполнения желудочков
645. IV ТОН СЕРДЦА РЕГИСТРИРУЕТСЯ НА ФОНОКАРДИОГРАММЕ
1) в фазу быстрого пассивного наполнения желудочков
2) при захлопывании створчатых клапанов
3) при сокращении предсердий и дополнительном поступлении крови в желудочки
646. МИТРАЛЬНЫЙ КЛАПАН ЛУЧШЕ ПРОСЛУШИВАЕТСЯ
1) справа от грудины у основания мечевидного отростка
2) во втором межреберье справа от грудины
3) в пятом межреберье слева на 1.5 см кнутри от среднеключичной линии
647. ТРЕХСТВОРЧАТЫЙ КЛАПАН ЛУЧШЕ ПРОСЛУШИВАЕТСЯ
1) во втором межреберье справа от грудины
2) в пятом межреберье слева на 1.5 см кнутри от среднеключичной линии
3) справа от грудины у основания мечевидного отростка
648. КЛАПАН ЛЕГОЧНОГО СТВОЛА ЛУЧШЕ ПРОСЛУШИВАЕТСЯ
КЛАПАН ЛЕГОЧНОГО СТВОЛА ЛУЧШЕ ПРОСЛУШИВАЕТСЯ
1) справа от грудины у основания мечевидного отростка
2) во втором межреберье справа от грудины
3) во втором межреберье слева от грудины
649. АОРТАЛЬНЫЙ КЛАПАН ЛУЧШЕ ПРОСЛУШИВАЕТСЯ
1) справа от грудины у основания мечевидного отростка
2) во втором межреберье слева от грудины
3) во втором межреберье справа от грудины
650. СУТЬ МЕТОДА ПЛЕТИЗМОГРАФИИ СОСТОИТ
1) в измерении давления крови в разные фазы кардиоцикла
2) в измерении сопротивления ткани электрическому току
3) в измерении объема части тела в зависимости от его наполнения кровью
651. СОКРАТИТЕЛЬНУЮ ФУНКЦИЮ МИОКАРДА ПОЗВОЛЯЕТ ИССЛЕДОВАТЬ МЕТОД
1) фонокардиография
2) сфигмография
3) плетизмография
4) баллистокардиография
652. ВОЛНЫ ПЕРВОГО ПОРЯДКА НА КРИВОЙ КРОВЯНОГО ДАВЛЕНИЯ, ЗАРЕГИСТРИРОВАННОЙ ПО СПОСОБУ ЛЮДВИГА, СВЯЗАНЫ
1) с фазами дыхания
2) с тонусом вазомоторного центра
3) с работой сердца
653. ВОЛНЫ ВТОРОГО ПОРЯДКА НА КРИВОЙ КРОВЯНОГО ДАВЛЕНИЯ,
ВОЛНЫ ВТОРОГО ПОРЯДКА НА КРИВОЙ КРОВЯНОГО ДАВЛЕНИЯ,
ЗАРЕГИСТРИРОВАННОЙ ПО СПОСОБУ ЛЮДВИГА, СВЯЗАНЫ
1) с работой сердца
2) с тонусом вазомоторного центра
3) с фазами дыхания
Установите соответствие.
654.
К ОТВЕДЕНИЯМ ЭКГ…. ОТНОСЯТСЯ
А.345 Монополярным 1. Стандартные отведения.
Б.12 Биполярным 2. Отведения по небу.
3. Усиленные от конечностей.
4. Грудные по Вильсону.
5. Интракардиальные.
655.
ФОНОКАРДИОГРАФИЧЕСКИЕ ТОНЫ СЕРДЦА…. ВОЗНИКАЮТ В ФАЗЫ СЕРДЕЧНОЙ ДЕЯТЕЛЬНОСТИ
A.1 I-й 1. Напряжения.
Б.3 II-й 2. Изгнания.
В.4 III-й 3. Конец протодиастолического периода.
Г.6 IV-й 4. Быстрого наполнения.
5. Медленного наполнения.
6. Предсистолический период.
656.
ФОНОКАРДИОГРАФИЧЕСКИЕ ОБУСЛОВЛЕНЫ ТОНЫ СЕРДЦА….
A.4 I-й 1. Сокращением предсердий.
Б.5 II-й 2. Закрытием створчатых клапанов.
В.3 III-й 3. Колебаниями стенок желудочков при быстром наполнении их кровью.
Г.1 IV-й 4. Захлопыванием створчатых клапанов и сокращением желудочков.
5. Закрытием полулунных клапанов.
657.
МЕТОД… ПРЕДСТАВЛЯЕТ СОБОЙ ГРАФИЧЕСКУЮ ЗАПИСЬ
А.3 Флебография 1. Колебаний потенциалов на поверхности тела,
Б.4 Реоплетизмография возникающих в результате
В.1 Электрокардиография сердечной деятельности.
Г.5 Сфигмография 2. Нет такого метода.
Д.6 Реокардиография 3. Колебаний стенок венозных сосудов.
4. Колебаний электрического сопротивления тканей при изменении кровенаполнения сосудов.
5. Пульсовых колебаний артериальных стенок.
6. Колебаний электрического сопротивления тканей сердца при его сокращениях.
658.
ФРАГМЕНТ ЭКГ…. ОТРАЖАЕТ ПРОЦЕСС
А.2 Зубец «Р» 1. Деполяризацию желудочков.
Б.5 Интервал «P-Q» 2. Деполяризацию предсердий.
В.1 Комплекс «QRS» 3. Реполяризацию желудочков.
Г.3 Зубец «Т» 4. Реполяризацию предсердий.
5. Деполяризацию атриовентрикулярного соединения.
659.
В ОТВЕДЕНИЯХ ЭКГ….. ЭЛЕКТРОДЫ РАСПОЛАГАЮТСЯ ТАК
А.1 I стандартное 1. Правая рука — левая рука.
Б.3 II стандартное 2. Левая рука — левая нога.
В.2 III стандартное 3. Правая рука — левая нога.
4. Правая рука — правая нога.
660.
ТЕРМИН… ОБОЗНАЧАЕТ УСИЛЕННОЕ ОТВЕДЕНИЕ ЭКГ
А.4 avR 1. От правой ноги.
Б.3 avL 2. От левой ноги.
В.2 avF 3. От левой руки.
4. От правой руки.
Раздел 6
Здоровая щитовидная железа — ключ к гармонии и красоте : ПОЗИТИВМЕД
Щитовидная железа — важнейший эндокринный орган, накапливающий йод и синтезирующий йодсодержащие гормоны: тиронин (трийодтиронин или Т3), тироксин (тетрайодтиронин или Т4). От состояния щитовидной железы зависят не только репродуктивная функция, работа органов пищеварения, иммунная и нервная системы, но и обмен веществ во всём организме человека.
Расположение щитовидной железы
Железа расположена в передней области шеи, над щитовидных хрящом, за что и получила свое название. Орган состоит из двух долей, соединённых перешейком. Форма щитовидной железы напоминает бабочку.
Тиреотропный гормон (ТТГ)
Работает и развивается щитовидная железа под влиянием ТТГ- тиреотропного гормона. Этот гормон вырабатывается в главной железе организма — гипофизе. Гипофиз выделяет тропные гормоны, которые определяют работу всех других желез. Таким образом, чем выше показатель ТТГ, тем сильнее стимулируется щитовидная железа.
Функция щитовидной железы
К функциям щитовидной железы относятся:
- накопление йода;
- выделение гормонов (Т3, Т4 и кальцитонин).
Гормон тироксин (Т4) и гормон трийодтиронин (Т3)
Это безусловно главный гормон щитовидной железы. Т4 образуется из аминокислоты тирозин, содержит 4 атома йода. Гормон активируется путём отщепления одного атома йода и превращения в Т3 гормон — трийодтиронин.
Эффекты тиреоидных гормонов
Гормоны воздействует практически на все клетки организма.
Сердце:
- хронотропный эффект — увеличение числа и аффинности бета-адренорецепторов, а воздействие на бета рецепторы повышает число сердечных сокращений и потребление кислорода клетками;
- инотропный эффект — усиление действия катехоламинов — то есть адреналина, а также повышение давления и частоты сердечных сокращений.
Жировая ткань: стимуляция липолиза, то есть расщепления жиров.
Мышцы: разрушение белков.
Кости:рост и развитие костей.
Нервная система: нормализация развития головного мозга. Эта функция особенно важна при беременности, так как при нескомпенсированном гипотиреозе у матери нервная система плода развивается неправильно, что может привести к различным патологиям развития ребенка.
Кишечник: увеличение всасывания углеводов.
Основная функция гормонов — ускорение катаболизма, что впоследствии влияет на эффективность и скорость обмена веществ.
Гормон кальцитонин
Щитовидная железа имеет также С-клетки, которые выделяют гормон пептидной природы — кальцитонин. Этот гормон отвечает за кальциево-фосфорный обмен и костеобразование в нашем организме. Показатель кальцитонина имеет значение, как онкомаркёр медуллярного рака щитовидной железы — это очень опасный, быстро растущий и метастатический рак. При этом раке кальцитонин повышается. При снижении кальцитонина ничего страшного не происходит, так люди после удаления щитовидной железы по определенным причинам живут спокойно, уровень кальция в крови поддерживается паратгормоном — гормоном паращитовидной железы.
Симптомы поражения щитовидной железы
Тиреотоксикоз — когда щитовидная железа выделяет слишком много гормонов. Соответственно обмен веществ разгоняется. Основные симптомы тиреотоксикоза:
- усиленное потоотделение, приливы
- снижение мышечной массы
- одышка
- увеличение груди и гинекоастия у мужчин
- немотивированная потеря веса
- тахикардия
- нарушение менструального цикла
- претибиальная мексидема (отёк)
- нервозность, раздражительность
- аритмия
- повышенный аппетит
- тремор
- мышечная слабость
- плохое состояние ногтей
Гипотиреоз — снижение функции щитовидной железы, когда она вырабатывает гормонов меньше, чем нужно, либо не вырабатывает их вообще. Симптомы гипотиреоза зависят от выраженности и очень неспецифичны. При лёгком гипотиреозе человек может не жаловаться ни на что вообще. При более выраженном отмечается:
- выпадение волос, особенно в области бровей
- задержка жидкости, отёки
- чувство зябкости и холода
- снижение потооделения
- сухость кожи
При тяжелом гипотиреозе:
- появление желтушности кожных покровов
- слабость, вялость, сонливость
- ухудшение памяти
- снижение когнитивных функций
- редкий сердечный ритм (брадикардия)
- снижение рефлексов
- запоры
- повышение холестерина
- повышение, иногда наоборот снижение давления
- увеличение языка
- понижение голоса, осиплость
- храп во сне
- нарушение менструального цикла
- прибавка веса.
Узлы щитовидной железы
Частой патологией щитовидной железы являются узлы. Их признакамимогут быть:
- ощущение сдавления шеи
- затруднение глотания
- чувство уплотнения в передней области шеи
- внешнее увеличение щитовидной железы
Не все узлы являются заболеванием, но при таких признаках необходимо проведение УЗИ щитовидной железы. Маленькие узлы могут никак себя не проявлять, потому рекомендовано делать УЗИ всем людям 1 раз в несколько лет. При обнаружении крупных узлов, некоторых признаках на УЗИ выполняется тонкоигольная аспирационная биопсия узла под контролем УЗИ. Необходимо также определение кальцитонина, как онкомаркера, с целью исключения онкопроцесса.
Таким образом, общее состояние щитовидной железы можно проверить по двум показателям: ТТГ и УЗИ. Если эти показатели показывают отклонения от норм, то необходимо проверить уровни гормонов Т3 и Т4, а также кальцитонина.
Гармония начинается там, где есть быстрый и эффективный обмен веществ. За качество обмена веществ в организме отвечает щитовидная железа, поэтому так важно следить за состоянием этого маленького, но такого важного органа.
Автор статьи — врач-эндокринолог медицинского центра «Позитивмед» Александр Владимирович Лынник.
Еще больше о направлении эндокринологии в нашем центре вы можете узнать здесь. Не стоит забывать, что наш организм очень хрупок — порой серьезные патологии возникают без явных симптомов. Специалисты нашего центра рекомендуют всем пациентам раз в несколько лет проходить обследование для определения общего состояния эндокринной системы.
Вы можете записаться на прием на нашем сайте, а также по телефону: 8 (812) 67-97-202
Ионная основа хронотропного действия ацетилхолина на синоатриальный узел кролика | Сердечно-сосудистые исследования
Цель: Цель состояла в том, чтобы изучить ионную основу хронотропных эффектов ванны с применением ацетилхолина и стимуляции блуждающего нерва на синусно-предсердном узле кролика. Методы: . Хронотропный эффект ацетилхолина, нанесенного на ванну, измеряли на отдельных клетках и небольших многоклеточных препаратах из синоатриального узла кролика, а хронотропный эффект постганглионарной стимуляции блуждающего нерва измеряли в интактном синоатриальном узле.Роль тока, активированного гиперполяризацией, i f , тока калия, активированного ацетилхолином, i K, ACh , и кальциевого тока L-типа, i Ca , была исследована путем блокировки токов с помощью 1-2 мМ Cs + или 10 −6 M UL-FS49, 0,2–1,0 мМ Ba 2+ и 6 × 10 −6 M нифедипин, соответственно. Результаты: В контрольных условиях небольшие многоклеточные препараты были примерно на два порядка менее чувствительны к ацетилхолину, нанесенному в ванну, чем отдельные клетки.Однако после блокирования ацетилхолинэстеразы эзерином в небольших многоклеточных препаратах чувствительность двух типов препаратов была примерно одинаковой. Блок i f либо не оказывал заметного эффекта, либо усиливал хронотропный эффект ацетилхолина, нанесенного в ванну, на отдельные клетки или небольшие многоклеточные препараты, тогда как частичный блок i K, ACh значительно его снижал. Точно так же блок i f не подавлял начальное замедление потенциалов спонтанного действия за счет стимуляции блуждающего нерва, тогда как частичный блок i K, ACh уменьшал его.Гиперполяризация заблокированного синоатриального узла в ответ на стимуляцию блуждающего нерва также была существенно снижена блоком i K, ACh . Частичный блок i Ca вызывал значительное снижение амплитуды потенциала действия и максимального диастолического потенциала, но небольшое снижение скорости спонтанных потенциалов действия и, следовательно, не имитировал эффект ацетилхолина. Выводы: Хронотропные эффекты ацетилхолина и стимуляции блуждающего нерва в ванне в основном не являются результатом подавления i f или i Ca , тогда как активация i K, ACh может играть важную роль.
ХРОНОТРОПНОЕ ДЕЙСТВИЕ АТФ И РОДСТВЕННЫХ СОЕДИНЕНИЙ, ИЗУЧЕННОЕ ПРИ ПРЯМОЙ ПЕРФУЗИИ SINUS NODE
Abstract
АТФ оказывает немедленное сильное, но относительно кратковременное отрицательное хронотропное действие на прямую перфузию синусового узла собаки. Атропин не меняет этот эффект.
Аденозин, A5MP и ADP действуют аналогично АТФ. Аденин и креатинфосфат не обладают значительным хронотропным действием. Следовательно, в АТФ фосфатные связи не важны для его хронотропного действия, но D-рибозная цепь важна.
Инозин, не обладающий хронотропным действием, отличается от аденозина только наличием гидроксильной группы вместо аминогруппы в 6-углеродном положении. Следовательно, аминогруппа и цепь D-рибозы необходимы для отрицательного хронотропного эффекта аденозина (и АТФ).
Гуанозин отличается от аденозина главным образом расположением аминогруппы у 2 атомов углерода вместо 6 атомов углерода, и его хронотропный эффект (ускорение) противоположен таковому у аденозина.Следовательно, положение аминогруппы имеет важное влияние на хронотропное действие этих пуриновых нуклеозидов.
A2MP, A3MP и циклический АМФ имеют одинаковое отрицательное хронотропное действие, которое отличается от действия АТФ, будучи более медленным в начале и менее выраженным в максимальной степени, но более продолжительным. Следовательно, положение фосфатных связей в D-рибозной цепи адениновых нуклеотидов изменяет характер их отрицательного хронотропного действия.
Два пиримидиновых нуклеозида (цитидин и уридин) и их нуклеотиды не обладают значительным хронотропным действием.Нуклеотид инозина активен, напоминая АТФ, а инозин — нет. Нуклеотид гуанозина неактивен, в то время как гуанозин ускоряет работу синусового узла, действие, на которое не влияет блокада бета -рецепторов. Значение различных действий инозина и гуанозина и их нуклеотидов неясно.
DNP вызывает относительно постепенное развитие брадикардии при прямой перфузии синусового узла и не зависит от атропина. Отрицательный хронотропный эффект ДНФ не обратим с помощью АТФ, но обратим с помощью ЦП (хотя и не всегда).Инверсия эффекта DNP с CP указывает на то, что синусовый узел собаки может использовать CP в качестве источника энергии.
На основании настоящих и предыдущих исследований этой лаборатории и в свете других недавних наблюдений предполагается, что отрицательное хронотропное действие дипиридамола связано с его эффектом сохранения аденозина.
Обсуждается возможное значение отрицательного хронотропного действия аденозина и АТФ по отношению к патофизиологии угнетения функции синусового узла в некоторых случаях острого инфаркта миокарда человека.
Pitt Кардиология | Лекарства, применяемые в кардиологии
Терминология
Для начала давайте рассмотрим терминологию, используемую для лекарств, влияющих на адренергическую передачу.
Симпатомиметики : препараты, которые действуют как нейротрансмиттеры, выделяемые симпатической нервной системой (например, норадреналин и адреналин).
В этот класс препаратов входят:
- Симпатомиметики прямого действия -> Лекарства, которые напрямую стимулируют α- и β-рецепторы.
- Симпатомиметики непрямого действия -> Лекарства, которые вызывают высвобождение норадреналина из нервных окончаний или блокируют захват нейромедиатора.
- Симпатомиметики смешанного действия -> Лекарства, которые вызывают высвобождение норадреналина из нервных окончаний И , стимулируют адренергические рецепторы.
Симпатолитики : препараты, которые препятствуют действиям симпатической нервной системы, такие как антагонисты α- и β-рецепторов.
Хронотропы : препараты, изменяющие частоту сердечных сокращений.
В этот класс препаратов входят:
- Отрицательные хронотропы -> Уменьшить частоту сердечных сокращений.
- Положительные хронотропы -> Увеличение частоты сердечных сокращений.
Инотропы : лекарственные средства или агенты, влияющие на сокращение сердечной мышцы
В этот класс препаратов входят:
- Отрицательные инотропы -> Ослабляют сердечные сокращения.
- Положительные инотропы -> Усиливают сердечные сокращения.
Адренергические препараты, используемые в кардиологии
Как обсуждалось в модуле адренергической передачи , препараты, которые изменяют адренергическую передачу, вызывают широкий спектр физиологических изменений. Ниже обсуждается действие адренергических препаратов, наиболее часто используемых в кардиологии. Описанные эффекты ограничиваются воздействием на сердечно-сосудистую систему.
Безусловно, в кардиологии используются различные препараты в дополнение к тем, которые действуют на симпатическую нервную систему, в том числе диуретиков , блокаторов кальциевых каналов и т. Д. Эти препараты будут обсуждаться позже в курсе и в последующих курсах, таких как как почечная блокада. Основная цель этого модуля — укрепить фундаментальные научные принципы, обсуждавшиеся в предыдущих лекциях, путем обсуждения действия определенных адренергических препаратов.
Симпатомиметики
Адреналин
В малых дозах адреналин в основном служит положительным инотропом и хронтропом.Однако дозы адреналина, обычно вводимые фармакологически, достаточны для стимуляции как α-, так и β-рецепторов. Следовательно, системные инъекции адреналина вызывают увеличение частоты сердечных сокращений и сократительной способности (за счет связывания с сердечными рецепторами β-1), и сужения сосудов (за счет связывания с рецепторами α-1 в гладких мышцах сосудов).
Однако увеличение общего периферического сопротивления, вызванного введением адреналина, весьма незначительно. Это связано с тем, что большие дозы адреналина активируют рецепторы α-1 и β-2 в мышечных артериолах, а физиологические эффекты компенсируются (связывание с
рецепторы α-1 способствуют сужению сосудов, связываясь с
β-2 рецепторы способствует
расширение сосудов).
Системный адреналин часто вводят до сердечно-легочной реанимации (при наличии брадикардии; остановка сердца) или пациентам с анафилактическим шоком . Его также можно использовать у пациентов с фибрилляцией желудочков.
Норэпинефрин
Основное различие в действии фармакологических доз норадреналина и адреналина состоит в том, что адреналин связывается с β-2 рецепторами, а норадреналин — нет.Оба этих препарата обладают хронотропным и инотропным действием, но норадреналин вызывает гораздо большее увеличение общего периферического сопротивления. Поскольку постнагрузка резко возрастает после введения норэпинефрина, увеличение сердечного выброса может быть ограничено, несмотря на стимулирующее действие препарата на сердце.
Поскольку норэпинефрин вызывает резкое повышение артериального давления, его введение активирует барорецепторный рефлекс. Это уменьшает действие препарата на сердце, поэтому хронотропные эффекты ослабляются, и препарат не вызывает тахикардию.
Введение норэпинефрина не вызывает каких-либо физиологических эффектов, связанных с агонистами β-2, таких как расширение гладких мышц дыхательных путей. Системный норадреналин используется для лечения глубокой гипотензии.
Дофамин
Хотя дофамин обычно считается нейротрансмиттером центральной нервной системы, дофамин также продуцируется и секретируется некоторыми почечными клетками . Фармакология системного введения дофамина сложна.При низком уровне доза препарат в основном связывается с рецепторами D1 в почках, что приводит к расширению почечных сосудов и снижению общего периферического сопротивления. Увеличение почечного кровотока приведет к увеличению диуреза, уменьшению задержки жидкости и уменьшению отеков.
| В более высоких дозах дофамин связывается с рецепторами β-1, вызывая сердечные эффекты. Интересно, что умеренные дозы дофамина оказывают необычное действие на сердце: избирательное увеличение силы сокращения миокарда происходит без значительного влияния на частоту сердечных сокращений.Высокие дозы дофамина вызывают как инотропный, так и хронотропный эффект. Кроме того, высокие дозы дофамина активируют рецепторы α-1, вызывая увеличение общего периферического сопротивления. |
Дофамин использовался для лечения пациентов с сердечной недостаточностью, поскольку он стимулирует сердечную функцию, вызывая при этом почечные эффекты, которые помогают очищать накопление жидкости в результате несоответствия накачки левого и правого желудочков.
Изопротеренол
Изопротеренол классифицируется как неселективный β-адренергический агонист, поскольку он хорошо связывается как с рецепторами β-1, так и с β-2, но имеет более высокую чувствительность к рецепторам β-1.Таким образом, изопротеренол обладает как хронотропным, так и инотропным действием, а также снижает постнагрузку, вызывая расширение сосудов мышечных артериол (посредством воздействия на β-2 рецепторы в гладких мышцах сосудов) . Таким образом, изопротеренол является идеальным лекарством для лечения пациентов с плохой сократимостью миокарда и низкой частотой сердечных сокращений, но с высоким периферическим сопротивлением. Клинически он используется чаще всего из-за его хронотропных эффектов.
Добутамин
Добутамин был разработан как структурный аналог изопротеренола.В фармакологических дозах он является агонистом β-1, но в очень высоких дозах он также связывается с рецепторами β-2. Таким образом, добутамин увеличивает частоту сердечных сокращений и сократительную способность, вызывая меньшие изменения периферического сопротивления, чем изопротеренол.
Добутамин — полезный препарат для лечения пациентов с низкой сократимостью сердца из-за органических заболеваний или хирургических вмешательств. Добутамин также широко используется в больницах в качестве фармакологического стресс-теста для выявления ишемической болезни сердца.Однако его использование ограничено склонностью к тахикардии.
Клонидин
Клонидин является агонистом рецепторов α-2, который проникает через гематоэнцефалический барьер и особенно эффективен при связывании с подтипом рецепторов α-2 в стволе мозга. Как отмечено в модуле адренергической передачи , рецепторы α-2 расположены на пресинаптических окончаниях норадренергических нейронов, включая те из нейронов, которые выступают в ростральную вентролатеральную мозговую оболочку (RVLM).Связывание клонидина с этими пресинаптическими рецепторами приводит к снижению высвобождения норэпинефрина в синапсе и, следовательно, к меньшему возбуждению постсинаптического нейрона. Следовательно, клонидин приводит к снижению активности нейронов RVLM, снижению активности симпатической нервной системы и снижению артериального давления. Клонидин используется для снижения артериального давления у пациентов, устойчивых к другим видам лечения гипертонии. Поскольку клонидин действует на центральную нервную систему, его также использовали не по прямому назначению для лечения пациентов с рядом неврологических и психиатрических проблем.Клонидин оказывает седативное действие из-за его действия на нервную систему, что является основным побочным эффектом препарата.
Классификация клонидина как симпатомиметического препарата может быть подвергнута сомнению, поскольку он не полностью попадает в эту категорию. Хотя препарат является агонистом адренорецепторов, его прием приводит к снижению активности симпатической нервной системы. Некоторые классифицируют клонидин как симпатолитическое средство. Терминология не применима к этому конкретному препарату.
фенилэфрин
Фенилэфрин является агонистом рецептора α-1. Фенилэфрин обычно используется в качестве вазопрессора для повышения артериального давления у нестабильных пациентов с гипотонией. Поскольку препарат вызывает внезапное повышение артериального давления, он может активировать барорецепторный рефлекс, тем самым вызывая рефлекторное снижение частоты сердечных сокращений и сократительной способности.
Симпатолитики
Празозин / Доксазоцин / Теразозин / Тамсулозин
Эти препараты являются антагонистами рецептора α-1.Все они снижают артериальное давление, вызывая расширение периферических сосудов и снижая общее периферическое сопротивление. Однако, как обсуждается в модуле адренергической передачи , начальное падение артериального давления, вызванное антагонистом рецептора α-1, вызывает опосредованное барорецепторным рефлексом увеличение частоты сердечных сокращений и сократимости, тем самым увеличивая потребность миокарда в кислороде. Следовательно, эти препараты являются препаратами второго ряда для лечения гипертонии, но обычно используются при других заболеваниях, таких как доброкачественная гиперплазия предстательной железы .
Феноксибензамин / фентоламин
Эти препараты являются антагонистами рецепторов α-1 и α-2. Они обладают такими же сосудорасширяющими эффектами, что и селективные антагонисты рецепторов α-1, но также блокируют пресинаптические рецепторы α-2 на периферии, в том числе на симпатических эфферентных волокнах в сердце. Как следствие, высвобождение норэпинефрина из симпатических нервных окончаний увеличивается, поскольку нормальное подавление пресинаптической обратной связи, опосредованное рецептором α-2, блокируется.Как отмечалось выше, вазодилататоры вызывают опосредованное барорецепторным рефлексом увеличение частоты сердечных сокращений и сократимости. Рефлекторно-опосредованное увеличение частоты сердечных сокращений и сократимости больше после введения комбинированного антагониста α-1 / α-2, чем селективного антагониста α-1, поскольку больше норадреналина высвобождается из симпатических постганглионарных окончаний в сердце.
Прочие лекарства
Антагонисты β-рецепторов
(β-блокаторы) обычно используются в кардиологии для лечения гипертонии, поскольку они снижают сердечный выброс за счет отрицательных хронотропных и инотропных эффектов.Эти препараты будут подробно рассмотрены позже в ходе курса.
Обзор
В таблице ниже кратко описаны действия препаратов, обсуждаемых в этом модуле.
| Наркотики | Агонист или Антагонист | Затронутые рецепторы | Влияние на общее периферическое сопротивление | Хронотропный эффект | Инотропный эффект | Прочая информация |
| Адреналин | Агонист | β1 = β2> α | Уменьшение с низкой дозой; Умеренное увеличение при высокой дозе | Положительно | Положительно | Меньшее повышение ОСО по сравнению с норадреналином |
| Норэпинефрин | Агонист | β1> α >> β2 | Увеличение | Положительно | Положительно | Значительно увеличивает постнагрузку; хронотропные эффекты ослабляются барорецепторным рефлексом; меньший риск тахикардии, чем у других бета-агонистов |
| Дофамин — низкая доза | Агонист | Дофамин> β> α | Уменьшение | Маленький | Маленький | В основном действует на почки, увеличивая почечный кровоток |
| Дофамин — умеренная доза | Уменьшение | Маленький | Положительно | |||
| Дофамин — высокая доза | Увеличение | Положительно | Положительно | |||
| Изопротеренол | Агонист | β1> β2 Нет α | Уменьшение | Положительно | Положительно | |
| Добутамин | Агонист | β1 >> β2> α | Немного, но уменьшается при высокой дозе | Положительно | Положительно | |
| фенилэфрин | Агонист | Селективная α1 | Увеличение | Уменьшение | Уменьшение | Воздействие на сердце обусловлено барорецепторным рефлексом |
| Клонидин | Агонист | Селективный α2 (действует в стволе мозга) | Уменьшение | Уменьшение | Уменьшение | Действует на центральную нервную систему; снижает активность нейронов RVLM |
| Празосин | Антагонист | Селективный α1 | Уменьшение | Увеличение | Увеличение | Хронотропные и инотропные эффекты осуществляются через механизмы рефлекса барорецепторов |
| Феноксибензамин | Антагонист | Селективная α | Уменьшение | Увеличение | Увеличение | Обладает более сильными хронотропными и инотропными эффектами, чем Празозин |
Основы сердечной фармакологии | Изучите педиатрию
Щелкните, чтобы открыть pdf: Основы сердечной фармакологии
Какие основные классы сердечных препаратов?
Существует 7 классов лекарств, сгруппированных по их физиологическому действию:
- Инотропные препараты
- Гликозиды наперстянки
- Симпатомиметические амины
- Ингибиторы фосфодиэстеразы
- Сосудорасширяющие препараты
- Ингибиторы ангиотензинпревращающего фермента
- Антагонисты рецепторов ангиотензина II типа 1
- Сосудорасширяющие средства прямого действия
- Блокаторы кальциевых каналов
- Органические нитраты
- Антиадренергические препараты
- Центральные ингибиторы адренорецепторов
- Антагонисты окончания симпатических нервов
- Антагонисты периферических альфа-адренорецепторов
- Антагонисты бета-адренорецепторов
- Противоаритмические препараты
- Антиаритмические средства I класса
- Антиаритмические средства класса IA
- Антиаритмические средства класса IB
- Антиаритмические средства класса IC
- Антиаритмические средства II класса
- Антиаритмические средства III класса
- Антиаритмические средства IV класса
- Аденозин
- Антиаритмические средства I класса
- Диуретики
- Петлевые диуретики
- Тиазидные диуретики
- Калийсберегающие диуретики
- Антитромботические препараты
- Ингибиторы тромбоцитов
- Антикоагулянтные препараты
- Липидорегулирующие препараты
- Ингибиторы редуктазы HMG CoA
- Агенты, связывающие желчные кислоты
- Ниацин
- Фибраты
Как они работают?
Инотропные препараты
Инотропные препараты действуют за счет увеличения силы сокращения желудочков при нарушении систолической функции миокарда.Хотя каждый препарат в этой группе действует через свой механизм, обычно все они работают для улучшения сердечного сокращения за счет увеличения внутриклеточной концентрации кальция. Когда внутриклеточный кальций увеличивается, взаимодействия сердечного актина / миозина усиливаются, что приводит к увеличению ударного объема и сердечного выброса для каждого наполнения желудочков.
- Гликозиды наперстянки (дигоксин)
Дигоксин, пожалуй, один из наиболее часто используемых сердечных гликозидов, который широко изучался.Дигоксин действует путем ингибирования активности Na-K АТФазы сарколеммы, что приводит к увеличению внутриклеточной концентрации Na. Это приводит к увеличению градиента натрия в транссарколемме и увеличивает обмен Na / Ca. Конечным результатом является повышение внутриклеточной концентрации Са, что означает, что сократительным белкам доступно больше Са. Ответ миокарда приводит к увеличению сократимости и увеличению сердечного выброса. Точно так же повышенная внутриклеточная концентрация натрия задерживает реполяризацию и, следовательно, снижает как синусовую частоту, так и AV-проводимость.
Адренергические агонисты имитируют действие симпатической нервной системы. Стимуляция бета1-адренорецепторов в сердце дает положительный инотропный (увеличивает сократительную способность), хронотропный (увеличивает частоту сердечных сокращений), дромотропный (увеличивает скорость проводимости через АВ-узел) и лузитропный (увеличивает расслабление миокарда во время диастолы). Эффекты агонистов бета1 опосредуются стимуляцией G-связанного белка, который стимулирует аденилатциклазу и генерирует цАМФ.Стимуляция G-связанного белка в конечном итоге приводит к активации цАМФ-зависимых протеинкиназ и фосфорилированию ключевых регуляторных белков в сердечных миоцитах.
Следует соблюдать осторожность при назначении этих препаратов в педиатрических условиях. Необходимо учитывать несколько факторов, в том числе: условия нагрузки, объемный статус и отзывчивость периферической сосудистой сети. Это может быть особенно сложно у тяжелобольных младенцев и детей. Кроме того, точное титрование этих препаратов дополнительно осложняется тем фактом, что существуют возрастные изменения в сцеплении рецептор-эффектор, активности киназ, доступности субстрата, активности фосфатазы и гидролизе цАМФ фосфодиэстеразами.Эти факторы способствуют возрастной вариабельности реакции на адренергические агонисты.
Примеры симпатомиметических аминов:
| Название лекарства | Банкноты |
| Дофамин | Допамин — это предшественник эндогенного катехоламина адреналина, который способен прямо или косвенно повышать кровяное давление. Дофамин вводят в виде непрерывной инфузии. Дофамин имеет специфические эффекты дофаминергического (DA) рецептора при низких концентрациях, эффекты рецептора Beta1 (положительная инотропия и хронотропия) при умеренных концентрациях и эффекты рецептора Alpha1 (сужение сосудов) при высоких концентрациях.Допамин также вызывает почечную вазодилатацию (controvercial). |
| Добутамин | Добутамин влияет в основном на бета-адренорецепторы. У детей Добутамин увеличивает сократительную способность и частоту сердечных сокращений, тем самым улучшая сердечный выброс. В результате может снизиться системное сопротивление сосудов (вторичное расширение сосудов). Добутамин не расширяет сосуды почек. Добутамин часто используется вместе с допамином, учитывая балансирующее воздействие на периферическое сосудистое сопротивление. |
| Изопротеренол | Изопротеренол представляет собой синтетический катехоламин, который является селективным в агонизме бета-рецепторов. Введение приведет к стимуляции как рецепторов Beta1, обнаруженных в основном в миокарде, так и периферических рецепторов Beta2. Введение изопротеренола увеличивает сократимость сердца и частоту сердечных сокращений за счет стимуляции рецептора Beta1, одновременно снижая системное сопротивление сосудов в результате стимуляции Beta2. Изопротеренол снижает сопротивление легочных сосудов и может помочь пациентам с легочной гипертензией.Побочные эффекты включают синусовую тахикардию и аритмию, поэтому пациенты должны находиться под тщательным наблюдением. |
- Ингибиторы фосфодиэстеразы
— это препараты, контролирующие деградацию внутриклеточного цАМФ. С повышением внутриклеточного уровня цАМФ увеличивается концентрация кальция в саркоплазме, что приводит к увеличению сократимости сердечной мышцы (положительная инотропия) с небольшим влиянием на хронотропию. В сосудистой сети повышенные уровни цАМФ связаны с расслаблением гладких мышц и расширением сосудов.
Примеры ингибиторов фосфодиэстеразы
| Название лекарства | Банкноты |
| Милриноне | Милринон — мощный и селективный ингибитор фосфодиэстеразы. Обычно милринон лучше переносится педиатрами, чем амринон, но может вызывать значительную гипотензию из-за сосудорасширяющего действия на периферическую сосудистую сеть. Также он может вызвать тромбоцитопению при длительном применении. Добутамин часто предпочтительнее милринона у новорожденных из-за его лучшего профиля побочных эффектов. |
| Амринон | Относительно слабый и неселективный ингибитор фосфодиэстеразы. Длительная пероральная терапия связана со значительными токсическими эффектами. В педиатрии используется нечасто. |
Сравнение различных типов инотропных препаратов
| Класс | Тип | Примеры | Эффект преобладающего агониста | Сократимость | После нагрузки | Пульс |
| Инотропный | Гликозиды наперстянки | Дигоксин | н / д Повышает внутриклеточный Ca. | ↑↑ | ↓ | |
| Симпатомиметические амины | Дофамин, | Бета1 = DA1> Альфа | ↑↑ | ↑↑ | ↑↑ | |
| Добутамин, | Бета1 | ↑↑ | (↓) | ↑ | ||
| изопротеренол, | Beta1 = Beta2 | ↑ | ↓↓ | ↑↑↑ | ||
| Адреналин, | Бета1 = Бета2 = Альфа | ↑↑ | ↑↑ | ↑↑ | ||
| норэпинефрин | Beta1 = Alpha | ↑↑ | ||||
| Ингибиторы фосфодиэстеразы | Амринон, Милринон | нет данных Повышает цАМФ, ↑ внутриклеточный Ca и вазодилат | ↑ | ↓↓ | ( |
Сосудорасширяющие препараты
Начальная вазоконстрикция — это механизм, используемый организмом для компенсации падения сердечного выброса.Это вазоконстрикция в основном опосредуется норадреналином и ангиотензином II. Венозное сужение может в конечном итоге привести к застою в легких из-за повышения гидростатического давления в легочных капиллярах. Венозное сужение также может привести к увеличению сопротивления, против которого должен сокращаться левый желудочек, и тем самым препятствует сердечному выбросу. Таким образом, назначаются сосудорасширяющие препараты, чтобы опосредовать усиление сужения и уменьшить нежелательные побочные эффекты компенсаторных механизмов.Эти препараты могут снижать повышенное кровяное давление и, следовательно, являются гипотензивными средствами. Каждый из сосудорасширяющих препаратов действует на определенные сосудистые участки, а некоторые (ингибиторы АПФ, альфа-блокаторы и нитропруссид натрия) способны воздействовать на обе стороны кровообращения.
Антиадренергические препараты
Обычно стимуляция симпатического нерва приводит к высвобождению норадреналина и стимуляции альфа- или бета-рецепторов. Как правило, каждый препарат воздействует на симпатическую нервную систему
Противоаритмические препараты
a) Хотя существует ряд различных систем классификации, эти препараты обычно делятся на четыре группы (классы с I по IV) в зависимости от их механизма действия, а также есть препараты, которые не попадают в эту классификационную схему и имеют просто названия. (например, аденозин)
- Основной причиной использования этих препаратов является устранение механизмов возникновения тахиаритмий.
б) Антиаритмические препараты I класса
- Блокировка быстрого натриевого канала, ответственного за деполяризацию фазы 0 потенциала действия
- Далее делятся на три подтипа в зависимости от степени блокады натриевых каналов и влияния препарата на рефрактерный период клетки.
c) Антиаритмические средства II класса
- Блокаторы бета-адренорецепторов
г) Антиаритмические средства III класса
- Эти препараты значительно продлевают потенциал действия, не влияя на рост деполяризации фазы 0
- Механизм действия заключается в блокировании реполяризующего калиевого тока во время фазы 2 и фазы 3 потенциала действия
e) Антиаритмические средства IV класса
- Блокировать медленный кальциевый канал L-типа
е) Аденозин
- Эндогенный нуклеозид с очень коротким периодом полураспада
- Самый эффективный препарат для устранения повторного ПСВТ
- Связывается со специфическими аденозиновыми рецепторами и активирует калиевые каналы.Когда калиевые каналы активированы, входящий внутрь калиевый ток гиперполяризует клеточную мембрану и тем самым подавляет спонтанную деполяризацию SA-узла, а также замедляет проводимость через AV-узел.
Диуретики
Чаще всего используется для лечения сердечной недостаточности и гипертонии и считается основой противозастойной терапии. При сердечной недостаточности наблюдается повышенная реабсорбция натрия и воды почками, что приводит к периферическим отекам и застою в легких.Диуретики используются для облегчения симптомов застойных явлений, связанных с избытком жидкости во внеклеточном пространстве. Клинический ответ на мочегонное средство зависит от того, насколько эффективно соль и вода поступают в почечные канальцы.
а) Петлевые диуретики
- Этот тип мочегонного средства действует на петлю Генле. В петле Генле петлевые диуретики ингибируют совместный транспорт хлорида натрия и калия в толстой восходящей конечности. Это приводит к снижению реабсорбированного хлорида, натрия и калия, а также к чистому увеличению выведения свободной воды.
б) Тиазидные диуретики
- Они работают, подавляя транспорт натрия и хлоридов в дистальных извитых канальцах нефрона.
c) Калийсберегающие диуретики
- Калийсберегающие диуретики обладают противозастойным действием, то есть увеличивают потерю воды без ущерба для калия.
г) Осмотические диуретики
- Осмотические диуретики предназначены для применения в острых случаях. Они обычно используются у пациентов с тяжелой недостаточностью кровообращения с ограниченным почечным выбросом.
Противотромботические препараты
Модуляция функции тромбоцитов и пути коагуляции помогает контролировать патогенез, связанный с такими сердечно-сосудистыми заболеваниями, как нестабильная стенокардия и острый инфаркт миокарда, а также такими заболеваниями, как венозный тромбоз, тромбы, связанные с фибрилляцией предсердий, дилатационная кардиомиопатия или механические протезы сердечных клапанов
Липидорегулирующие препараты
- Помогите модулировать патогенез атеросклероза
- Лекарства, улучшающие липидный исход, являются кардиозащитными и ингибируют прогрессирование атеросклероза
Рекомендации для педиатрических пациентов
Пероральный прием лекарств — наиболее распространенный метод, применяемый у детей.Новорожденные, по сравнению с младенцами и детьми старшего возраста, имеют меньшую секрецию кислоты в желудке в течение первых нескольких недель после рождения. Время прохождения через ЖКТ более быстрое в течение первых 5 лет детства, и это может повлиять на всасывание лекарств, вводимых в препарате с замедленным высвобождением
Распределение лекарств
Лекарства распределяются из плазмы в разные органы на основе их сродства к тканям с высоким содержанием воды или липидов. Например, у недоношенных детей вода составляет 85% МТ, тогда как у годовалого ребенка она составляет 60%.Когда процент внеклеточной воды высок, как это было бы у новорожденного пациента, лекарство необходимо вводить в большей дозе n в миллиграммах на килограмм для достижения той же концентрации в сыворотке
Количество воды в организме меняется с развитием
| Возраст | Внеклеточная вода (%) | Всего воды в организме (%) |
| Недоношенные дети | 60 | 85 |
| Доношенный | 55 | 75 |
| 5-месячный ребенок | 50 | 60 |
| 1-летний ребенок | 40 | 60 |
| Подростки | 40 | 60 |
Метаболизм лекарств
- Преимущественно в печени
2 основных направления:
- Фаза 1: метаболизирует вещества и состоит из оксидазной системы со смешанными функциями, системы cyp450 и путей N-деметилирования.
- Недоношенные и доношенные новорожденные имеют от 50 до 75% этих ферментов по сравнению с уровнями у младенцев.
- Это означает, что препараты, метаболизируемые этими системами, могут иметь более высокую концентрацию в сыворотке при той же относительной дозе у новорожденных по сравнению с младенцами.
- Фаза 2: реакции конъюгирования эндогенных соединений или определенных лекарств и их метаболитов в водорастворимые соединения путем глюкуронизации или ацетилирования.
- Их обычно достаточно для метаболизма большинства лекарств и эндогенных соединений через 1-2 недели после рождения у доношенных детей
- Дозы некоторых лекарств должны быть соответственно скорректированы в первые несколько недель неонатальной жизни, иначе новорожденный может быть предрасположен к лекарственной токсичности
Выведение наркотиков
Основной путь выведения лекарств — через почки.Выведение лекарства также может происходить через желчные, желудочно-кишечные и дыхательные пути в меньшей степени. У новорожденных наблюдается снижение почечной элиминации, что необходимо учитывать при дозировании Rx.
Развитие почек у детей
| Возраст | GFR (мл / мин / 1,73 м2) | Макс тубулярная секреция (мг / мин / 1,73) |
| Доношенный новорожденный | 33 | 16 |
| 1 месяц | 50 | 30 |
| 2 месяца | 70 | 50 |
| 6 месяцев | 110 | 60 |
| 3 года | 130 | 75 |
Обычные терапевтические и токсические концентрации лекарственных средств в сыворотке
| Сердечно-сосудистый агент | Терапевтический уровень | Уровень токсичности |
| Дигоксин | 0.От 5 до 2 нг / мл | > 3 нг / мл |
| Лидокаин | от 2 до 5 мг / мл | > 6 мг / мл |
| Хинидин | от 1 до 5 мг / мл | > 7 мг / мл |
| прокаинамид | от 4 до 10 мг / мл | > 10 мг / мл |
Сводная таблица
| Класс | Как они работают? | Тип | Примеры |
| Инотропный |
| Гликозиды наперстянки | Дигоксин |
| Симпатомиметические амины | допамин, добутамин, изопротеренол, адреналин, норэпинефрин | ||
| Ингибиторы фосфодиэстеразы | Амринон, Милринон | ||
| Сосудорасширяющий |
|
|
|
| Антиадренергические |
|
| Сальбутамол (блокатор бета-2) Празосин (блокатор Альфа-1) |
| Антиаритмический |
| Антиаритмические средства I класса — Блокировать быстрый натриевый канал, ответственный за деполяризацию потенциала действия фазы 0 — Далее делятся на три подтипа в зависимости от степени блокады натриевых каналов и влияния препарата на рефрактерный период клетки
| 1A — прокаинамид 1B — Лидокаин 1С — Пропафенон |
Антиаритмические средства II класса
| Пропранолол | ||
Антиаритмические средства III класса
| Амиодарон | ||
Антиаритмические средства IV класса
| Верапамил | ||
Аденозин
| Аденозин | ||
| Диуретики | Чаще всего используется для лечения сердечной недостаточности и гипертонии, а также для облегчения симптомов застойных явлений, связанных с избытком жидкости во внеклеточном пространстве |
| фуросемид Гидрохлоротиазид спиронолактон |
| Противотромботические препараты | Модуляция функции тромбоцитов и пути коагуляции помогает контролировать патогенез, связанный с такими сердечно-сосудистыми заболеваниями, как нестабильная стенокардия и острый инфаркт миокарда, а также такими заболеваниями, как венозный тромбоз, тромбы, связанные с фибрилляцией предсердий, дилатационная кардиомиопатия или механические протезы сердечных клапанов | Ингибиторы тромбоцитов Антикоагулянтные препараты | Активатор тканевого плазминогена Варфарин Гепарин |
| Липидорегулирующие препараты | улучшенные липидные исходы являются кардиозащитными и подавляют прогрессирование атеросклероза | Ингибиторы редуктазы HMG CoA Агенты, связывающие желчные кислоты |
Возможное участие независимого от циклического аденозинмонофосфата механизма в положительном хронотропном эффекте норэпинефрина в изолированном правом предсердии морской свинки | Анестезиология
ТЕРАПЕВТИЧЕСКИЕ подходы с ингибиторами фосфодиэстеразы III, такими как аморинон и милринон, к пациентам с синдромом низкого сердечного выброса были разработаны для периоперационного контроля кровообращения.1,2 Эти агенты вызывают увеличение сократимости сердца и одновременное расширение сосудов, ингибируя деградацию циклического аденозинмонофосфата (АМФ) в кардиомиоцитах и гладкомышечных клетках сосудов. 3,4 Интересно, что введение одного ингибитора фосфодиэстеразы III или комбинированное лечение катехоламинами, такими как добутамин, эффективно увеличивает сократимость сердца с минимальным увеличением частоты сердечных сокращений в клинических условиях. 1,5–8 Поскольку считается, что повышение уровня циклического АМФ участвует в опосредованном β-адренорецептором положительном хронотропном эффекте, а также в положительном инотропном эффекте, 9 кажется странным, что ингибиторы фосфодиэстеразы III модулируют опосредованный β-адренорецептором инотропный эффект. и хронотропные эффекты по-разному.
Недавно сообщалось о независимых от циклических АМФ эффектах агонистов β-адренорецепторов в нескольких тканях, таких как гладкие мышцы трахеи, гладкие мышцы коронарных артерий и сердечные миоциты. 10–12 В сообщениях предполагается, что прямое взаимодействие стимулирующего гуанозин-5′-трифосфат-связывающего белка (Gs) с ионными каналами имеет отношение к циклическому AMP-независимому механизму β-адренергической стимуляции. Агонисты β-адренорецепторов оказывают положительный хронотропный эффект, изменяя несколько ионных токов, таких как потенциал-зависимый ток Ca 2+ L-типа (I Ca (L) ) и входящий ток, активируемый гиперполяризацией (I f ), которые вносят вклад в конфигурацию потенциала действия пейсмекерной клетки синоатриального узла.13 Таким образом, возможно, что агонисты β-адренорецепторов модулируют эти ионные каналы мембранно-ограниченным образом без соответствующего изменения вторичного мессенджера циклического АМФ.
Целью настоящего исследования было выяснить, вовлечено ли увеличение внутриклеточного циклического АМФ в положительный хронотропный эффект, опосредованный β-адренорецепторами. С этой целью мы сравнили фармакологически положительный хронотропный эффект норадреналина, эндогенного симпатического нейромедиатора, с действием форсколина, прямого активатора аденилатциклазы, с использованием ингибиторов фосфодиэстеразы, потенциал-зависимого ингибитора каналов Ca 2+ верапамила и I f ингибитор каналов хлорида цезия (CsCl) в спонтанно поражающемся правом предсердии морской свинки.Если положительный хронотропный эффект норадреналина опосредован активацией аденилатциклазы, он будет усилен ингибиторами фосфодиэстеразы. Кроме того, верапамил и CsCl должны аналогичным образом модулировать хронотропные реакции на норэпинефрин и форсколин.
Все эксперименты были выполнены в соответствии с Руководящими принципами исследований с участием животных и людей Медицинской школы Университета Йокогамы (Иокогама, Япония).
Использовали морских свинок-самцов массой 250–350 г. После выполнения торакотомии по средней линии во время анестезии диэтиловым эфиром сердце быстро иссекали и помещали в чашку для препарирования, заполненную насыщенным кислородом раствором Кребса-Хенселейта следующего состава: 119,0 мм NaCl, 2,5 мм CaCl 2 , 4,8 мм KCl 1,2 мм MgSO 4 , 1,2 мм KH 2 PO 4 , 24,9 мм NaHCO 3 и 10.0 мм глюкозы.
Спонтанно бьющееся правое предсердие осторожно рассекали и устанавливали вертикально в 10-миллилитровую стеклянную камеру с двойными стенками, заполненную раствором Кребса-Хенселейта, который непрерывно газировали 95% O 2 и 5% CO. и поддерживали при 36 ° C. Нижний конец правого предсердия был закреплен на крючке, а верхний конец был соединен шелковой нитью с изометрическим датчиком силы-смещения (модель UL-10GR, Minebea, Нагано, Япония) через предусилитель (модель AS2103, Sanei -Сокки, Токио, Япония).Диастолическое давление довели до 0,5 г. Частоту самопроизвольных биений подсчитывали по диаграмме, фиксирующей его развитое напряжение. Перед началом эксперимента препарат уравновешивали не менее 60 мин.
Кривые «концентрация-ответ» для положительного хронотропного эффекта норадреналина и форсколина были определены кумулятивным способом путем увеличения их концентраций с шагом 0,5 молярной логарифмической единицы.Разница между максимальной скоростью биения, производимой агентами, и базовой скоростью биения была принята за 100%, и были нанесены на график процентные изменения скорости биения, производимой различными концентрациями агентов. При изучении влияния антагониста α 1 -адренорецепторов, антагониста β-адренорецепторов, ингибиторов фосфодиэстеразы или ингибиторов ионных каналов на хронотропные эффекты агентов кривые «концентрация-ответ» строили через 30 мин после применения каждого теста. сложный.Поскольку ингибиторы фосфодиэстеразы и ингибиторы ионных каналов изменяли частоту спонтанных биений, скорость биений в присутствии каждого соединения принималась за базальную частоту биений. Концентрации норэпинефрина или форсколина, вызывающие полумаксимальный ответ (ЕС 50 ) в отсутствие и в присутствии различных тестируемых соединений, получали из графиков log-probit индивидуального ответа как функции концентрации. Следующее уравнение было использовано для получения значения pK B :
, где [A] представляет собой EC 50 агониста в присутствии антагониста, [A] 0 представляет собой EC 50 агониста в отсутствие антагониста, а [B] представляет собой концентрацию антагониста.
Чтобы сделать эффекты испытуемых соединений на хронотропные эффекты норэпинефрина и форсколина понятными, кривые «концентрация – ответ», изображенные на основе абсолютных значений частоты биений, были вставлены в соответствующие рисунки.
Левое предсердие было рассечено и установлено вертикально в стеклянной камере, как описано ранее, за исключением того, что в ванне поддерживалась температура 30 ° C.Натяжение покоя, приложенное к препарату, доводили до 1 г. Атриум подвергался электрическому стимулированию с помощью прямоугольных импульсов с частотой 0,5 Гц, длительностью 3 мс и напряжением в 1,2 раза превышающим пороговое значение, подаваемым парой спиральных платиновых электродов, подключенных к электрическому стимулятору (SEN-7203, Nihon-Kohden, Tokyo, Япония) через изолирующий блок (SS-201J, Nihon-Kohden). Перед началом эксперимента препарат уравновешивали не менее 60 мин.
Кривая «концентрация-ответ» для инотропного эффекта норадреналина была определена кумулятивным способом путем увеличения его концентрации с шагом 0.5 log молярная единица. Разница между максимальной сократительной силой, создаваемой норэпинефрином, и базальной сократительной силой была принята за 100%. При изучении влияния амринона на инотропный эффект норадреналина кривые «концентрация – ответ» строили через 30 мин после применения амринона. Поскольку амринон увеличивал сократительную силу, это значение в присутствии амринона принимали за базальную сократительную силу.
Были использованы следующие соединения: норэпинефрин, атенолол, празозин, амринон, (±) верапамил, форсколин, 3-изобутил-1-метилксантин (IBMX) и CsCl (Sigma Chemical Co., Сент-Луис, Миссури). Атенолол, форсколин и амринон растворяли в диметилсульфоксиде и дополнительно разбавляли дистиллированной водой. Все остальные химические вещества растворяли в дистиллированной воде.
Влияние CsCl, ингибитора I f , и верапамила, ингибитора I Ca (L) , на положительные хронотропные эффекты форсколина и норадреналина показано на рисунках 4 и 5.Сам CsCl в концентрации 2 мМ снижал частоту спонтанных биений на 56 ± 11 ударов / мин (n = 12), тогда как верапамил в концентрации 0,2 мкм приводил к снижению частоты спонтанных биений на 27 ± 19 ударов / мин (n = 9). . В присутствии CsCl форсколин и норэпинефрин увеличивали частоту сердечных сокращений на 87 ± 11 ударов / мин (n = 6) и 82 ± 9 ударов / мин (n = 6) соответственно. Однако влияние CsCl на кривые нормализованная концентрация-ответ норадреналина и форсколина было противоречивым. CsCl имел тенденцию усиливать положительный хронотропный эффект форсколина при более низких концентрациях и подавлять эффект агента при более высоких концентрациях, что приводило к пересечению двух кривых концентрация-ответ при концентрации форсколина около 0.5 мкм. Однако в целом CsCl не оказывал значительного влияния на нормированную кривую «концентрация – ответ» для форсколина (рис. 4A). С другой стороны, он значительно подавлял нормализованную кривую «концентрация-ответ» для норадреналина (рис. 4B). Как и в случае с CsCl, хотя кривая концентрация-ответ для форсколина в присутствии верапамила пересекала контрольную кривую ответа в аналогичной точке, верапамил не влиял на положительный хронотропный эффект форсколина в целом (рис.5А). Напротив, верапамил значительно подавлял положительный хронотропный эффект норадреналина (рис. 5B). В присутствии верапамила форсколин и норэпинефрин увеличивали частоту сердечных сокращений на 149 ± 10 ударов / мин (n = 4) и 124 ± 16 ударов / мин (n = 5) соответственно.
Рис. 4. Кривые «концентрация-ответ» для форсколина ( A ) и норадреналина ( B ) в отсутствие (белый кружок; n = 8 для форсколина и n = 6 для норадреналина) и в присутствии 2 мМ CsCl ( замкнутый круг; n = 6 для форсколина и n = 6 для норадреналина) в правом предсердии спонтанно бьющейся морской свинки.Баллы являются средними ± стандартное отклонение. Увеличение частоты сердечных сокращений (ударов в минуту [уд / мин]), производимое форсколином или норэпинефрином, было принято за 100%. Когда препарат обрабатывали CsCl, частота биений в присутствии CsCl принималась за базальную частоту биений. * Значительно отличается от соответствующего контроля ( P <0,05) по результатам дисперсионного анализа с последующим тестом Бонферрони t . ( Врезка ) Кривые «концентрация – реакция», изображенные на основе изменений абсолютных значений частоты биений.C = контроль; T = лечение.
Рис. 4. Кривые концентрация-ответ для форсколина ( A ) и норадреналина ( B ) в отсутствие (белый кружок; n = 8 для форсколина и n = 6 для норадреналина) и в присутствии 2 мМ CsCl. (закрашенный кружок; n = 6 для форсколина и n = 6 для норадреналина) в правом предсердии спонтанно бьющейся морской свинки. Баллы являются средними ± стандартное отклонение. Увеличение частоты сердечных сокращений (ударов в минуту [уд / мин]), производимое форсколином или норэпинефрином, было принято за 100%.Когда препарат обрабатывали CsCl, частота биений в присутствии CsCl принималась за базальную частоту биений. * Значительно отличается от соответствующего контроля ( P <0,05) по результатам дисперсионного анализа с последующим тестом Бонферрони t . ( Врезка ) Кривые «концентрация – реакция», изображенные на основе изменений абсолютных значений частоты биений. C = контроль; T = лечение.
Рис.5. Кривые «концентрация – ответ» для форсколина ( A ) и норадреналина ( B ) в отсутствие (белый кружок; n = 8 для форсколина и n = 6 для норадреналина) и в присутствии 0,2 мкм верапамила (закрашенный кружок; n = 4 для форсколина и n = 5 для норадреналина) в правом предсердии морской свинки со спонтанным биением. Баллы являются средними ± стандартное отклонение. Увеличение частоты сердечных сокращений (ударов в минуту [уд / мин]), производимое форсколином или норэпинефрином, было принято за 100%. Когда препарат обрабатывали верапамилом, частота сердечных сокращений в присутствии верапамила принималась за базальную частоту сердечных сокращений.* Значительно отличается от соответствующего контроля ( P <0,05) по результатам дисперсионного анализа с последующим тестом Бонферрони t . ( Врезка ) Кривые «концентрация – реакция», изображенные на основе изменений абсолютных значений частоты биений. C = контроль; T = лечение.
Рис. 5. Кривые «концентрация-ответ» для форсколина ( A ) и норадреналина ( B ) в отсутствие (белый кружок; n = 8 для форсколина и n = 6 для норадреналина) и при наличии 0.2 мкм верапамила (закрашенный кружок; n = 4 для форсколина и n = 5 для норадреналина) в правом предсердии спонтанно бьющихся морских свинок. Баллы являются средними ± стандартное отклонение. Увеличение частоты сердечных сокращений (ударов в минуту [уд / мин]), производимое форсколином или норэпинефрином, было принято за 100%. Когда препарат обрабатывали верапамилом, частота сердечных сокращений в присутствии верапамила принималась за базальную частоту сердечных сокращений. * Значительно отличается от соответствующего контроля ( P <0,05) по результатам дисперсионного анализа с последующим тестом Бонферрони t .( Врезка ) Кривые «концентрация – реакция», изображенные на основе изменений абсолютных значений частоты биений. C = контроль; T = лечение.
В текущем исследовании мы продемонстрировали, что положительный хронотропный эффект, опосредованный β-адренорецепторами, не усиливался ингибиторами фосфодиэстеразы амриноном и IBMX, тогда как положительный хронотропный эффект, продуцируемый форсколином, усиливался обоими агентами.Ингибитор I f CsCl и ингибитор I Ca (L) верапамил замедляли скорость увеличения частоты биений, продуцируемых норадреналином. С другой стороны, ни CsCl, ни верапамил существенно не влияли на нормированную кривую «концентрация – ответ» для положительного хронотропного эффекта форсколина. Эти результаты предполагают, что основной механизм положительного хронотропного эффекта β-адренергической активации отличается от механизма активатора аденилатциклазы форсколина.
Активация симпатической нервной системы вызывает как положительные инотропные, так и хронотропные эффекты через β-адренорецепторов в сердечных миоцитах и клетках синоатриального узла. Считается, что система аденилатциклаза – циклический АМФ играет ключевую роль в опосредованном β-адренорецептором положительном хронотропном эффекте, а также в положительном инотропном эффекте. 9,14–16 Подтверждая это мнение, форсколин оказывает положительный хронотропный эффект, как показано в текущем исследовании.Внутриклеточное применение циклического АМФ также дает положительный хронотропный эффект. 17,18
Поскольку циклический АМФ дегенерируется фосфодиэстеразой, ингибиторы фосфодиэстеразы теоретически могут не только имитировать, но и потенцировать циклические АМФ-опосредованные ответы. Фактически, было продемонстрировано, что положительный инотропный эффект изопреналина на желудочковые клетки человека и кролика, а также антимиграционный эффект форсколина на гладкомышечные клетки аорты крысы усиливаются ингибиторами фосфодиэстеразы.19–21 Если положительный хронотропный эффект β-адренергической стимуляции также будет опосредован циклическим АМФ, эффект должен быть усилен ингибированием фосфодиэстеразы. Однако ингибитор фосфодиэстеразы III амринон не усиливал положительный хронотропный эффект норадреналина, хотя амринон сам по себе не только увеличивал частоту сердечных сокращений, но и усиливал положительный хронотропный эффект форсколина. Поскольку существует несколько типов фосфодиэстеразы, 22циклический АМФ, усиленный β-адренергической стимуляцией в клетках синоатриального узла морской свинки, может не дегенерировать под действием фосфодиэстеразы III.Чтобы выяснить эту возможность, мы также исследовали влияние неспецифического ингибитора фосфодиэстеразы IBMX на положительные хронотропные эффекты норэпинефрина и форсколина. В соответствии с результатом, полученным с амриноном, IBMX усиливал действие форсколина, но не действие норадреналина. Кроме того, амринон может усиливать положительный инотропный эффект норадреналина в левом предсердии морской свинки. Шахид и Роджер 20 также сообщили, что и амринон, и IBMX усиливали положительный инотропный эффект изопреналина в папиллярных мышцах кролика, но не положительный хронотропный эффект агента в правом предсердии кролика.
Есть как минимум три возможных способа интерпретации текущих результатов. Во-первых, положительный хронотропный эффект норадреналина не зависит от β-адренорецепторов. Во-вторых, циклический АМФ, увеличивающийся за счет β-адренергической активации, компартментализируется в пейсмейкерной клетке, как было предложено Jurevicius и Fischmeister, 23 и используемые ингибиторы фосфодиэстеразы труднодоступны в этом отсеке. В-третьих, генерация циклического АМФ причинно не связана с положительным хронотропным эффектом норадреналина.Как показано на рисунке 1, селективный антагонист β 1 -адренорецепторов атенолол сдвинул вправо кривую «концентрация – ответ» для положительного хронотропного эффекта норадреналина. Значение pK B , равное 6,8 для атенолола в концентрации 1 мкМ, указывает на участие β 1 -адренорецепторов в положительном хронотропном эффекте норадреналина в текущем эксперименте. 24 Недавние сообщения продемонстрировали участие α 1 -адренорецептора в регулировании частоты сердечных сокращений.25,26 Однако празозин не изменил положительный хронотропный эффект норадреналина в текущем исследовании, что указывает на то, что α 1 -адренорецептор играет второстепенную роль в контроле частоты сердечных сокращений в текущих экспериментальных условиях. Амринон не усиливал положительный хронотропный эффект норадреналина даже в присутствии празозина. Следовательно, α 1 -адренорецепторы, активируемые норадреналином, также не связаны с неспособностью амринона усиливать положительный хронотропный эффект норадреналина.
Частота синусового узла определяется скоростью спонтанного срабатывания потенциала действия кардиостимулятора в синоатриальном узле. Считается, что агонисты β-адренорецепторов, а также увеличение внутриклеточного циклического АМФ увеличивают частоту спонтанной активации посредством активации каналов I f и каналов I Ca (L) . 13,17,27 Таким образом, мы исследовали влияние ингибитора каналов I f CsCl 28 и ингибитора каналов Ca 2+ верапамила на положительные хронотропные эффекты норадреналина и форсколина.CsCl и верапамил сдвинули кривую нормализованная концентрация-ответ для норадреналина вправо. С другой стороны, ни CsCl, ни верапамил существенно не повлияли на нормированную кривую «концентрация – ответ» для форсколина. Даже если циклический АМФ, который был увеличен норадреналином, был бы разделен на части, поскольку циклический АМФ имел отношение к положительному хронотропному эффекту норадреналина, и CsCl, и верапамил должны модулировать положительные хронотропные эффекты норэпинефрина и форсколина аналогичным образом.Однако с текущими результатами дело обстоит иначе. Взятые вместе, текущие результаты предполагают, что циклический АМФ причинно не связан с основным механизмом положительного хронотропного эффекта норадреналина в сердце морской свинки. Однако следует отметить, что нельзя сделать решающий вывод, пока не будут получены прямые доказательства, такие как отсутствие увеличения циклического АМФ в пейсмекерной клетке после лечения норадреналином или продуцируемое норадреналином увеличение активности ионных каналов в ограниченных мембранами. модель.
Yatani et al. 29,30 сообщил, что связанный с β-адренорецептором гуанозин-5′-трифосфат-связывающий белок Gs может активировать оба канала I Ca (L) и I f напрямую, хотя циклическая AMP-зависимая активация обоих каналов уже была продемонстрирована . 16,31 Они также показали динамику продуцируемого изопротеренолом увеличения I Ca в изолированных кардиомиоцитах морских свинок.32 Изопротеренол увеличивал его двухфазным образом, с быстрой постоянной времени 150 мс и медленной постоянной времени 36 с. Форсколин и IBMX медленно увеличивали I Ca с постоянными времени 56 и 150 с соответственно. Таким образом, они пришли к выводу, что быстрый ответ миоцита на изопротеренол был вызван прямой модуляцией Gs на каналах I Ca (L) , а медленный ответ был опосредован вторичным мессенджером циклическим АМФ. 32 Изопротеренол также быстро активировал I f с постоянной времени 800 мс в бесклеточной конфигурации пластыря наизнанку.30 Эти прямые модуляции активируемых β-адренорецепторами Gs на каналах I Ca (L) и I f считаются удобными для симпатической нервной системы для регулирования частоты сердечных сокращений. 33,34 Текущее исследование может продемонстрировать эти прямые взаимодействия β-адренорецепторов-ионных каналов Gs, которые были продемонстрированы с помощью экспериментов с патч-зажимом, в функциональных экспериментах.
Однако следует отметить, что существуют некоторые ограничения для экстраполяции текущих данных на клинические условия.Мы использовали ткани предсердий и подсчитали количество мышечных сокращений в минуту вместо подсчета количества потенциала действия в клетках кардиостимулятора. Кроме того, мы использовали изолированные ткани. Хотя изоляция ткани или клетки упрощает сложную задачу, следует проявлять большую осторожность при экстраполяции данных на все тело, которое сложно регулируется нервной системой и гормональным контролем. Мы также должны учитывать различие видов. Базовая частота сердечных сокращений морской свинки намного выше, чем у человека, что свидетельствует о существовании тонкой разницы во внутриклеточной регуляции активации потенциала действия в клетках кардиостимулятора между людьми и морской свинкой.
Использование ингибиторов фосфодиэстеразы III амринона и милринона является важным терапевтическим подходом, особенно в сочетании с катехоламинами, у пациентов с синдромом низкого сердечного выброса. 2 Комбинация ингибиторов фосфодиэстеразы III с катехоламинами синергетически увеличивает сократимость сердца, тогда как увеличение частоты сердечных сокращений минимально. 1,8 Эти особенности, по-видимому, соответствуют идеальному инотропному лечению при периоперационном низком сердечном выбросе 1, поскольку увеличение частоты сердечных сокращений ставит под угрозу баланс потребности миокарда в кислороде.35 Настоящее исследование может объяснить, по крайней мере частично, причину дифференциального воздействия ингибиторов фосфодиэстеразы III на сократимость и частоту сердечных сокращений, наблюдаемых в клинических условиях. Поскольку сердце иннервируется симпатической нервной системой, сердечные β-адренорецепторы в различной степени активируются эндогенным норадреналином. Следовательно, ингибиторы фосфодиэстеразы III могут синергетически усиливать положительный инотропный эффект эндогенных и экзогенных катехоламинов, минимально влияя на эти хронотропные эффекты.
В заключение следует отметить, что амринон и IBMX усиливают положительный хронотропный эффект форсколина. Однако препараты не усиливали хронотропный эффект норадреналина. И CsCl, и верапамил сдвинули вправо нормализованную кривую «концентрация-ответ» для положительного хронотропного эффекта норадреналина, не влияя на кривую «концентрация-ответ» для форсколина. Эти результаты убедительно свидетельствуют о том, что циклический АМФ не имеет отношения к положительному хронотропному эффекту активации β-адренорецепторов в сердце морской свинки.
Авторы благодарят Noritsugu Tohse, M.D., Ph.D. (Профессору и заведующему кафедрой физиологии Медицинского колледжа Саппоро, Саппоро, Япония) за конструктивную критику.
Оказывает ли глюкагон положительный инотропный эффект на сердце человека? | Сердечно-сосудистая диабетология
Вскоре после того, как экспериментальные исследования показали его положительные инотропные и хронотропные эффекты, пациентам был назначен глюкагон.В пионерском исследовании его сердечно-сосудистые эффекты у людей были изучены во время диагностической катетеризации сердца. У 11 пациентов внутривенное введение 3-5 мг значительно увеличило сердечный индекс, среднее артериальное давление, частоту сердечных сокращений и максимальную скорость развития давления в левом желудочке, но конечное диастолическое давление в левом желудочке и системное сосудистое сопротивление существенно не изменились. В другой группе из шести пациентов, получавших болюс 1 мг глюкагона, сердечный индекс и частота систолической фракции выброса увеличились, тогда как системное сосудистое сопротивление снизилось.На основании этих результатов авторы предлагают глюкагон в качестве потенциально полезного препарата для лечения острой сердечной недостаточности [13]. Однако противоречивые результаты были получены при назначении глюкагона для лечения состояний с низким сердечным выбросом. Например, 24 из 50 пациентов, страдающих сердечной недостаточностью, кардиогенным шоком или и тем и другим, показали некоторое клиническое улучшение после добавления глюкагона к комбинированной терапии, которая включала дигитализацию, фуросемид и спиронолактон. Однако остальные 26 пациентов не ответили на лечение, и 16 из них умерли [14].В другом исследовании сообщалось, что непрерывная инфузия глюкагона в средней дозе 4 мг / ч в течение нескольких дней приводила к отчетливому улучшению клинического состояния 12 из 16 пациентов с сердечной недостаточностью или кардиогенным шоком. Это улучшение, отмеченное повышением артериального давления и диуреза, а также уменьшением одышки, легочных хрипов, потоотделения и периферических отеков, могло быть связано с сопутствующим лечением, поскольку 9 из этих пациентов ранее не получали диуретики. препараты получали диуретики в процессе терапии глюкагоном [15].Некоторые данные указывают на то, что положительное влияние на работу сердца, о котором сообщалось в некоторых случаях после введения глюкагона, больше связано с влиянием на периферическое сосудистое сопротивление, чем с прямым положительным инотропным эффектом глюкагона. Например, внутривенное введение глюкагона вызвало повышение сердечного выброса и частоты сердечных сокращений у трех пациентов с тяжелым стенозом аорты. Значительно повысилось конечное диастолическое давление в левом желудочке и заметно увеличился объем левого желудочка.Тем не менее, средняя скорость сокращения окружности как показатель сократимости не изменилась. По мере роста систолического давления в левом желудочке наблюдалось значительное увеличение силы стенки, возможно, из-за эффекта Франка-Старлинга в расширенном левом желудочке. Таким образом, авторы пришли к выводу, что наблюдаемые сердечные изменения были в основном вторичными по отношению к увеличению венозного возврата [16]. В дальнейшем исследовании, направленном на изучение гемодинамических эффектов глюкагона центральных и периферических сосудов у пациентов с органическими заболеваниями сердца, введение 2 или 5 мг глюкагона внутривенно приводило к значительному увеличению сердечного выброса и улучшению сердечной деятельности, но снижению периферического и легочного сосудистого сопротивления. также наблюдалось снижение легочного артериального давления [17].Таким образом, эти результаты свидетельствуют о том, что сосудорасширяющее действие глюкагона [18] снижает системное сосудистое сопротивление [13, 17], снижает сердечную нагрузку и улучшает функцию почек, таким образом улучшая сердечную деятельность и клиническое состояние пациентов. Метаболические эффекты глюкагона могут способствовать улучшению сердечной деятельности. Действительно, гипергликемия, возникающая в результате глюконеогенеза и гликогенолиза в печени [16], может улучшить метаболизм миокарда и, следовательно, сердечную функцию, учитывая «метаболическое ремоделирование», происходящее при сердечной недостаточности, которое способствует окислению сердечной глюкозы, а не окислению жирных кислот, являющемуся основным источником. энергии для миокарда в нормальных условиях [19].Вероятность того, что гемодинамические эффекты глюкагона, о которых сообщают у людей, связаны с увеличением сократительной способности сердца в результате активации рецептора глюкагона, очень маловероятна, поскольку глюкагон лишен какого-либо прямого инотропного действия на сердце человека. Это было продемонстрировано «in vitro» на сосочковых мышцах желудочков, полученных из эксплантированных сердец человека [20], а также на трабекулах миокарда предсердий, полученных от пациентов, перенесших операцию на сердце для восстановления клапана (неопубликованные результаты).Это мнение дополнительно подтверждается недавним открытием, что транскрипты мРНК рецепторов глюкагона отсутствуют в левом желудочке (наиболее важная камера сердца для поддержания адекватных гемодинамических параметров) и что только следы были обнаружены в левом или правом предсердии и правом желудочке в 2 случаях. из 15 исследованных человеческих сердец [21]. В соответствии с этим открытием, влияние глюкагона на сократительную способность миокарда у людей было незначительным [22] или нулевым [23].Действительно, хотя, как указано выше, в некоторых случаях сообщалось о положительных результатах, считается, что глюкагон лишен положительных клинических эффектов у пациентов с застойной сердечной недостаточностью, и его введение не рекомендуется в текущих руководствах по лечению сердечной недостаточности [24,25,26 , 27]. При передозировке бета-блокаторами или блокаторами кальциевых каналов клиническое улучшение было связано с введением глюкагона в нескольких отчетах о случаях, но его клиническая эффективность не оценивалась ни в одном контролируемом клиническом исследовании; некоторый положительный эффект, о котором сообщалось, мог быть связан с другими сопутствующими видами терапии, полученными этими пациентами [28].Действительно, глюкагон не всегда улучшает выживаемость у этих пациентов, и сообщалось о невозможности ответа на глюкагон, особенно у субъектов с токсичностью пропранолола [29]. Благоприятные эффекты глюкагона при передозировке антагонистов кальция или β-рецепторов, по-видимому, связаны с обращением брадикардии, а не с улучшением подавленной сократимости миокарда [29]. Действительно, глюкагон оказывает положительное хронотропное действие у людей [9, 13, 17], что может быть связано с более высокой экспрессией рецепторов глюкагона в синоатриальном узле, чем в рабочем миокарде, подобно правому предсердию крысы [3].Кроме того, индуцированное глюкагоном усиление активации симпатических нервов может способствовать хронотропному эффекту глюкагона, поскольку экспериментальные данные показывают, что глюкагон стимулирует симпатическую активность, действуя на уровне гипоталамуса [30] и повышая уровень циркулирующих катехоламинов [31]. Однако необходимы дальнейшие исследования, чтобы установить фактический механизм, ответственный за положительный эффект глюкагона у пациентов с симптоматической брадикардией.
Границы | Моделирование хронотропного действия изопреналина на синоатриальный узел кролика
Введение
β-Адренергическая стимуляция увеличивает частоту сердечных сокращений за счет ускорения спонтанной активности водителя ритма сердца, синоатриального узла (SAN; Abramochkin et al., 2009). Считается, что это происходит за счет модуляции ионных токов, опосредованной β-адренорецепторами, которые вносят вклад в активность кардиостимулятора; однако точные ионные механизмы, лежащие в основе эффекта β-адренергической стимуляции, еще полностью не выяснены. Эксперименты показали, что изопреналин (ISO), агонист β-адренорецепторов, увеличивает ток кальция L-типа ( I CaL ; Noma et al., 1980; Walsh et al., 1988; Zaza et al., 1996 ; Виноградова и др., 2002; Ке и др., 2007; Алиг и др., 2009), калиевый ток запаздывающего выпрямителя (как I Kr , так и I Ks ; Walsh et al., 1988; Duchatelle-Gourdon et al., 1989; Giles et al., 1989; Yazawa and Kameyama , 1990; Freeman, Kass, 1993; Lei et al., 2000, 2002; Ke et al., 2007; Виноградова и др., 2008) и сдвигает зависимую от напряжения активацию тока, активированного гиперполяризацией ( I f ) в сторону положительных потенциалов, что приводит к увеличению I f (Brown et al., 1979; Cai et al., 1995; ДиФранческо, 1995; Заза и др., 1996; Accili et al., 1997a, b; Bucchi et al., 2003; Барбути и др., 2007; Алиг и др., 2009; Baruscotti et al., 2010; Liao et al., 2010). Поскольку I f считается основным водителем ритма в клетках водителя ритма млекопитающих, предполагается, что увеличение I f является основным механизмом, лежащим в основе положительного хронотропного эффекта β-адренергической стимуляции (DiFrancesco , 1995, 2010; Заза и др., 1996; ДиФранческо и Борер, 2007; Liao et al., 2010). Однако было обнаружено, что ISO увеличивает скорость стрельбы кроличьего SAN, когда I f было заблокировано Cs + (Cai et al., 1995), предполагая, что I f улучшение не может быть преимущественно отвечает за положительное хронотропное действие ISO. Предыдущие исследования показали, что ISO увеличивает амплитуду систолического подъема внутриклеточной концентрации Ca 2+ ([Ca 2+ ] i ) в сердечных клетках (Ju and Allen, 1999; Huser et al., 2000; Шеннон и др., 2004; Мальцев, Лакатта, 2009; Wu et al., 2009). Это повышает вероятность того, что изменения в [Ca 2+ ] и с помощью ISO могут способствовать увеличению скорости стрельбы за счет механизма «Ca 2+ clock» (Виноградова и др., 2002, 2008; Мальцев и Лакатта, 2009). Ряд исследований показал, что вмешательства, изменяющие [Ca 2+ ] и , изменяют скорость активации пейсмекерных клеток (Hagiwara, 1989; Ju and Allen, 1999; Huser et al., 2000; Шеннон и др., 2004; Виноградова и др., 2008; Мальцев, Лакатта, 2009; Wu et al., 2009). Как в клетках сердца млекопитающих (Hagiwara, 1989; Huser et al., 2000; Vinogradova et al., 2002, 2008; Maltsev, Lakatta, 2009; Wu et al., 2009), так и в клетках кардиостимулятора амфибий, в которых I f отсутствует (Ju and Allen, 1999), было обнаружено, что скорость воспламенения зависит от амплитуды переходного процесса [Ca 2+ ] i : агенты, модифицирующие высвобождение SR Ca 2+ , соответственно влияют на скорострельность.β-Адренергическая стимуляция увеличивает амплитуду переходного процесса [Ca 2+ ] i . Утверждалось, что большая часть увеличения скорости возбуждения, вызванной β-стимуляцией, по-видимому, происходит за счет механизма «часов Ca 2+ » (Виноградова и др., 2002, 2008; Мальцев и Лакатта, 2009). Также сообщалось, что ISO увеличивает амплитуду входящего устойчивого тока ( I st ; Guo et al., 1995, 1997; Shinagawa and Noma, 2000; Toyoda et al., 2005) и амплитуду I Kr и I Ks (Lei et al., 2000, 2002). Также было обнаружено смещение кривой активации I Kr в сторону более отрицательных мембранных потенциалов и увеличение скорости его дезактивации (Yazawa and Kameyama, 1990; Lei et al., 2000, 2002; Ke et al., 2007). Поскольку I st активируется в диапазоне мембранного потенциала от -70 до -50 мВ (Guo et al., 1995, 1997; Shinagawa and Noma, 2000), увеличение I st может играть роль. в ускорении темпа активности кардиостимулятора.Увеличение скорости дезактивации I Kr снижает выходной ток, что приводит к относительно большему чистому входящему току во время диастолической деполяризации, что также может способствовать увеличению скорости спонтанной активности. В этом исследовании мы модифицировали существующие модели электрической активности центральных и периферических клеток SAN кролика (Boyett et al., 2001; Kurata et al., 2008), чтобы смоделировать положительный хронотропный эффект ISO и изучить лежащую в основе ионную основу. для действий ISO с использованием вычислительного подхода.Мы включили уравнения для зависимых от концентрации воздействий ISO на макроскопическую проводимость I CaL , I Kr и I Ks , I st и для описания изменений кинетики. из I f и I Kr , а также в [Ca 2+ ] i . Эти уравнения построены на основе экспериментальных данных. Используя модели, смоделированный эффект ISO показывает зависимость от концентрации, которая количественно согласуется с экспериментальными записями.Чтобы исследовать лежащую в основе ионную основу воздействия ISO, мы исследовали его влияние в диапазоне концентраций на токи, ответственные за активность кардиостимулятора SAN, и мы исследовали индивидуальную роль воздействия ISO на токи I CaL , I f , I Kr , I st и [Ca 2+ ] i . Мы обнаружили, что хронотропный эффект ISO отражает комплексное воздействие на ионные токи I CaL , I f , I Kr , I Ks , I и st [Ca 2+ ] i , среди которых повышение скорости дезактивации I Kr представляется особенно важным, в то время как влияние ISO на I f и [Ca 2 + ] i тоже играет роль.
Разработка модели
На основе экспериментальных данных о кинетике ионных каналов и региональных различий в плотностях ионного тока были разработаны хорошо известные модели потенциалов действия центра и периферии SAN-клеток кролика (Zhang et al., 2000; Boyett). et al., 2001; Kurata et al., 2008). Zhang et al. (2000) модели были модернизированы за счет включения внутреннего устойчивого тока ( I st ) и уравнений регуляции внутриклеточного Ca 2+ (Boyett et al., 2001). В этом исследовании мы модифицировали модели Boyett et al. (2001) и Kurata et al. (2008) путем включения уравнений, описывающих действия ISO на I CaL , I f , I Kr , I Ks , I st и [Ca 2+ ] и .
Увеличение
I CaL
Экспериментально установлено, что
Изопреналин увеличивает I CaL без значительного изменения его кинетики (Zaza et al., 1996). Увеличение I CaL зависит от дозы. Уравнение, описывающее дозозависимое увеличение I CaL , было построено на основе экспериментальных данных Zaza et al. (1996) получены из клеток SAN кролика, которые имеют вид:
fCa = fCa, макс. [ISO] K0,5, Ca + [ISO] (1)
Где f Ca — процент увеличения I CaL . Подгоняя уравнение. 1 к данным Zaza et al. (1996; кружки) и Виноградова и др.(2002; треугольники), как показано на рисунке 1A, мы получили наиболее подходящие значения для f Ca, max (максимальное процентное увеличение I CaL ) и K 0,5, Ca (ISO концентрация, необходимая для полумаксимального увеличения I CaL ) как 0,54 (т.е. 54%) и 7 нМ, соответственно. Полученное значение K 0,5, Ca (7 нМ) соответствует экспериментальным данным EC 50 ISO на I CaL (Zaza et al., 1996). Сплошная линия на рисунке 1A показывает взаимосвязь между увеличением I CaL и концентрацией ISO, предсказанной уравнением. 1.
Рис. 1. Эффекты ISO , зависящие от концентрации. (A) Дозозависимое фракционное увеличение на г CaL . Открытые кружки: экспериментальные данные Zaza et al. (1996) из клеток SAN кролика. Сплошная линия: рассчитано по модели. (B) Дозозависимый фракционный сдвиг кривой активации I f .Открытые кружки: данные Zaza et al. (1996) из клеток SAN кролика. Сплошная линия: рассчитано по модели. (C) Дозозависимое фракционное увеличение на г Kr . Закрашенные кружки: данные Yazawa and Kameyama (1990) по клеткам SAN морских свинок. Открытый треугольник: данные Lei et al. (2000) из клеток SAN кролика. Сплошная линия: рассчитано по модели. (D) Дозозависимый сдвиг кривой активации I Kr . Открытый треугольник: данные Lei et al. (2000) из клеток SAN кролика.Сплошная линия: рассчитано по модели. (E) Дозозависимое дробное уменьшение постоянной времени дезактивации I Kr . Открытые треугольники: данные Lei et al. (2000) из клеток SAN кролика. Сплошная линия: рассчитано по модели. (F) Дозозависимое дробное увеличение г st . Открытый квадрат: данные Guo et al. (1995) из клеток SAN кролика. Сплошная линия: рассчитано по модели.
Сдвиг кривой активации
I f
Изопреналин увеличивает I f , сдвигая его кривую активации, зависящую от напряжения, в сторону более положительных мембранных потенциалов (Zaza et al., 1996; Accili et al., 1997a, b). Уравнение для описания сдвига S f (в мВ) было построено на основе экспериментальных данных Zaza et al. (1996) получены из клеток SAN кролика, как показано ниже:
Sf = Sf, макс. [ISO] nfK0.5, fnf + [ISO] nf (2)
Подгоняя уравнение. 2 к данным Zaza et al. (1996; рис. 1B, кружки), мы получили наиболее подходящие значения для S f, max (максимальное смещение), K 0,5, f (концентрация ISO, необходимая для полумаксимального смещения активации. кривая) и n f (коэффициент Хилла) как 9.62 мВ, 13,5 нМ и 0,392 соответственно, что близко к полученным Zaza et al. (1996). Сплошная линия на рисунке 1B показывает взаимосвязь между сдвигом кривой активации I f и концентрацией ISO, предсказанной уравнением. 2.
Действия с
I Kr и I Ks
Изопреналин имеет комплексное действие на I K . Сообщалось, что β-адренергические агонисты увеличивают I K в клетках SAN кролика (Lei et al., 2000, 2002), небольшие многоклеточные препараты SAN кролика (Walsh et al., 1988), клетки SAN морских свинок (Freeman and Kass, 1993), миоциты желудочков морских свинок (Yazawa and Kameyama, 1990) и клетки предсердий лягушки (Giles et al., 1989). В клетках SAN кролика I K состоит из двух компонентов: быстро активируемого I Kr и медленно активируемого I Ks , из которых I Kr обычно является преобладающим компонентом ( Habuchi et al., 1995; Ито и Оно, 1995; Лей и Браун, 1996; Lei et al., 2000, 2002). Было обнаружено, что ISO увеличивает амплитуду тока I Kr и I Ks (Lei et al., 2000, 2002), увеличивает скорость дезактивации I Kr в клетках SAN кролика. (Lei et al., 2000; Ke et al., 2007), и сдвинуть кривую активации I Kr в сторону более отрицательных мембранных потенциалов в миоцитах желудочков морских свинок (Yazawa and Kameyama, 1990), клетках предсердий лягушки. (Джайлз и др., 1989), и клетки SAN кролика (Lei et al., 2000).
Увеличение
г крон и г Ks ( I Kr и I Ks )
Уравнение для зависящего от концентрации увеличения г Kr было построено на основе экспериментальных данных Lei et al. (2000, 2002), полученные из клеток SAN кролика, показанных на рисунке 1C (треугольники), и данные Yazawa и Kameyama (1990), полученные из миоцитов желудочков морских свинок, показанных на рисунке 1C (кружки).Уравнение принимает вид:
fK = fK, макс. [ISO] K0,5, gK + [ISO] (3)
Где f K — процентное увеличение I Kr . Подгоняя уравнение. 3 к экспериментальным данным Lei et al. (2000, 2002) и Yazawa and Kameyama (1990), мы получили наиболее подходящие значения для f K, max (максимальное процентное увеличение I K ) и K 0,5, Ca ( концентрация ISO, необходимая для увеличения наполовину максимального значения г Kr ) как 1.87 (т.е. 187%) и 19 нМ соответственно. Сплошная линия на рисунке 1C показывает взаимосвязь между увеличением г Kr и концентрацией ISO, предсказанной уравнением. 3.
Изопреналин также увеличивает I Ks : экспериментальные данные показали, что 10 нМ ISO увеличивают I Ks примерно на 20% (Lei et al., 2002), что сопоставимо с увеличением I Kr с аналогичной концентрацией по ISO.Как из-за этого сходства, так и из-за отсутствия полных экспериментальных данных «концентрация-ответ», в этом исследовании мы предположили аналогичную зависимую от концентрации модуляцию по ISO г Ks и г Kr , оба из которых были смоделированы по формуле. 3.
Смена активации
I Kr
Уравнение для описания сдвига зависимого от напряжения отношения активации I Kr ( S K ) было построено на основе экспериментальных данных Lei et al.(2000), полученные из клеток SAN кролика. Поскольку доступных экспериментальных данных в отношении зависимости эффекта от концентрации нет, мы предположили, что зависящий от концентрации сдвиг кривой активации I Kr такой же, как и дозозависимое увеличение г Kr , который принимает вид:
SK = SK, макс. [ISO] K0,5, Kacti + [ISO] (4)
Путем подбора уравнения. 4 к экспериментальным данным Lei et al. (2000), как показано на рисунке 1D (треугольник), мы получили значение наилучшего соответствия S K, max (максимальный сдвиг I K , в мВ) как -15 мВ и K 0.5, Kacti как 7,5 нМ.
Увеличение скорости деактивации
I Kr
Уравнение, описывающее увеличение скорости дезактивации I Kr , было построено на основе экспериментальных данных для I K от Lei et al. (2000), полученные из клеток SAN кролика, показанных на Фигуре 1E (треугольники). На рисунке данные представлены как уменьшение постоянной времени дезактивации (τ Кр ). Мы снова предположили, что дозозависимое изменение τ Kr такое же, как дозозависимое увеличение г Kr .Уравнение принимает вид:
dτKr = dτKr, макс [ISO] K0,5, τKr + [ISO] (5)
dτKr — частичное уменьшение τ Kr . Подгоняя уравнение к экспериментальным данным Lei et al. (2000), мы получили значение наилучшего соответствия dτKr, max как 3,0 и K 0,5 , τ Kr как 24 нМ.
Увеличение
I st
Сообщается, что
изопреналин увеличивает I st (Guo et al., 1995, 1997; Синагава и Нома, 2000; Toyoda et al., 2005). Уравнение увеличения I st было построено на основе экспериментальных данных, полученных на клетках SAN кролика (Guo et al., 1997). Из-за отсутствия достаточных экспериментальных данных мы предположили, что зависящее от концентрации увеличение I st такое же, как зависящее от концентрации увеличение г Kr . Уравнение принимает вид:
fst = fst, max [ISO] K0.5, ст + [ISO] (6)
Путем подбора уравнения. 6 к экспериментальным данным Guo et al. (1997), как показано на рисунке 1F (незакрашенный квадрат), мы получили значение наилучшего соответствия для f st, max как 1,0 и K 0,5, st как 33 нМ.
Ca
2+ Обработка
Изопреналин увеличивает амплитуду и минимальный диастолический уровень [Ca 2+ ] и у млекопитающих (Hagiwara, 1989; Huser et al., 2000; Vinogradova et al., 2002, 2008; Шеннон и др., 2004; Мальцев, Лакатта, 2009; Wu et al., 2009) и пейсмекерные клетки амфибий (Ju, Allen, 1999). Было обнаружено, что ISO изменяет поглощение и высвобождение SR Ca 2+ путем стимуляции кальмодулинкиназы II (CamKII; Shannon et al., 2005; Мальцев, Лакатта, 2009). При моделировании воздействия ISO мы использовали подход Харче и др. (2011), чтобы изменить уравнения обработки Ca 2+ для увеличения амплитуды и минимального диастолического уровня [Ca] i , как это наблюдалось в экспериментальных исследованиях (Ju and Allen, 1999; Виноградова и др., 2002) за счет увеличения максимального выделения SR Ca 2+ (на 20%) и уменьшения потоков высвобождения SR Ca 2+ (на 20%).
Уравнения 1–6 были включены в математические модели потенциала действия Boyett et al. (2001) и Kurata et al. (2008) для центральных и периферических клеток SAN кролика, чтобы моделировать хронотропный эффект ISO и исследовать ионную основу, лежащую в основе такого хронотропного эффекта.
Результаты
Хронотропный эффект изопреналина
Смоделированные эффекты ISO на потенциалы спонтанного действия показаны на рисунке 2.На рисунке слева показаны результаты, рассчитанные для центрального (A) и периферийного (C) Boyett et al. модели клеток, на правой панели показаны результаты, рассчитанные для центрального (B) и периферийного (D) Kurata et al. сотовые модели. Для каждого типа клеток и каждой модели показаны эффекты ISO с концентрацией 0,05 мкМ. Во всех клеточных моделях ISO влиял на форму потенциалов действия: он увеличивал скорость возбуждения, сокращал продолжительность потенциала действия, увеличивал амплитуду потенциалов действия и максимальный диастолический потенциал (максимальный диастолический потенциал становился более отрицательным).Эти смоделированные эффекты ISO на потенциалы действия согласуются с экспериментальными наблюдениями на клетках SAN кролика (Zaza et al., 1996; Lei et al., 2000).
Рисунок 2. I CaL , I f , I K (сумма I Kr и I Ks ), и I st во время потенциалов действия в управлении и с ISO . Данные в контроле и с 0,05 мкМ ISO показаны для: (A) Boyett et al .; (B) Kurata et al.модели центральных ячеек. (C) Boyett et al .; (D) Kurata et al. модели периферических клеток. Во всех четырех моделях ячеек ISO вызывал увеличение скорости самопроизвольного возгорания, что сопровождалось увеличением I CaL , I f , I K и I st .
Влияние ISO на ионные токи, лежащие в основе потенциалов действия (верхняя панель на каждом из рисунков 2; левые панели для исследования Boyett et al.центральная (рис. 2А) и периферическая (рис. 2С) модели клеток; и правые панели для Kurata et al. Затем были изучены центральная (рис. 2В) и периферийная (рис. 2D) клеточная модель [а именно на I CaL , I f , I K (сумма I Kr и I Ks ) и I st ]. Профили I CaL (вторая панель), I f (третья панель), I K (четвертая панель) и I st (пятая панель). ) показаны на рисунке 2 в контрольных условиях и при наличии 0.05 мкМ ISO. В смоделированном присутствии ISO входящие токи I CaL , I f и I st были увеличены, в то время как чистый I K увеличился в фазе реполяризации, но уменьшился в фаза диастолической деполяризации. Повышенное значение I K во время реполяризации потенциала действия увеличивало скорость реполяризации (тем самым сокращая продолжительность потенциала действия) и увеличивало максимальный диастолический потенциал.Увеличение входных токов I CaL , I f и I st вместе с уменьшением внешнего тока I K в фазе деполяризации увеличивало скорость деполяризации. и, следовательно, скорость спонтанной активности. Увеличение амплитуды потенциала действия можно отнести к увеличению I CaL .
Смоделированные эффекты ISO демонстрируют четкую зависимость от концентрации: чем выше смоделированные концентрации ISO, тем сильнее наблюдается хронотропный эффект.Это показано на рисунке 3, на котором потенциалы действия, вычисленные в контрольных условиях, накладываются на потенциалы, вычисленные с различными концентрациями ISO для центрального (A) и периферического (C) Boyett et al. модели клеток (левые панели), а также центральную (B) и периферическую (D) Kurata et al. (правые панели) модели ячеек. При моделировании с использованием метода Boyett et al. (2001) модели, 0,005 мкМ ISO увеличивали скорость на 7% для модели центральной клетки и на 6% для модели периферической клетки. 0,05 мкМ ISO увеличивал скорость на 20% для модели центральной клетки и на 15% для модели периферической клетки.0,5 мкМ ISO увеличивал скорость на 21% для модели центральной клетки и на 16% для модели периферической клетки. При моделировании с использованием Kurata et al. (2008) модели, 0,005 мкМ ISO увеличили скорость на 13% для модели центральной клетки, но не значительно изменили скорость для модели периферических клеток. 0,05 мкМ ISO увеличивает скорость на 25% для модели центральной клетки и на 7% для модели периферической клетки; 0,5 мкМ ISO увеличивал скорость на 29% для модели центральной клетки и на 16% для модели периферической клетки. Результаты, суммирующие смоделированные хронотропные эффекты ISO, показаны на рисунке 4, на котором процент уменьшения продолжительности цикла кардиостимуляции (BCL) нанесен на график зависимости от концентрации ISO.Данные, рассчитанные на основе моделей центральных ячеек Boyett et al. и Kurata et al. показаны на рисунках 4A, B соответственно. Данные, рассчитанные на моделях периферических клеток Boyett et al. и Kurata et al. показаны на рисунках 4C, D соответственно. Смоделированные данные (сплошная линия) сравнивали с экспериментальными данными, полученными Lei et al. Для изолированных клеток SAN кролика. (2000; незакрашенный кружок), Zaza et al., 1996; открытый треугольник, обращенный вправо), Choi et al. (1999; открытый ромб) и Barbuti et al. (2007; открытый перевернутый треугольник).Экспериментально 0,02 мкМ ISO увеличивал скорость на 13% (Lei et al., 2000), аналогично моделированной реакции всех четырех моделей клеток на ту же концентрацию ISO. Несоответствие моделирования и экспериментальных данных Zaza et al. (1996) или Choi et al. (1999) незначительна и может быть связана с ограничениями модели (см. Обсуждение). Обратите внимание, что данные моделирования из моделей центральных клеток лучше соответствовали экспериментальным данным, чем модели периферических клеток, поскольку в этих экспериментальных исследованиях рассматривались только первичные клетки SAN без различия между центральными и периферическими клетками.Роли I CaL , I f , I Kr , I st и [Ca 2+ ] i в полученной ISO положительной хронотропии были также считается, как показано на рисунке 5.
Рисунок 3. Зависимое от концентрации влияние ISO на потенциалы действия . (A) Boyett et al .; (B) Kurata et al. модели центральных ячеек. (C) Boyett et al.; (D) Kurata et al. модели периферических клеток. Потенциалы действия показаны в контрольных условиях и в присутствии 0,005, 0,05 и 0,5 мкМ ISO. Интенсивность самопроизвольной стрельбы увеличивалась с помощью ISO в зависимости от дозы.
Рис. 4. Зависимое от концентрации уменьшение продолжительности цикла кардиостимуляции (BCL) в процентах согласно ISO . (A, B) Boyett et al. и Kurata et al. модели центральных ячеек. (C, D) Boyett et al.и Kurata et al. модели периферических клеток. На рисунке представлены данные моделей и соответствующих экспериментальных данных «концентрация-реакция». Черная линия: данные стандартной конфигурации модели, в которой эффекты ISO учтены для всех I CaL , I f , I Kr , I Ks , I st и Ca 2+ переходный. Открытые кружки: экспериментальные данные Lei et al. (2000) из клеток SAN кролика.Открытый правый треугольник: данные Zaza et al. (1996) из клеток SAN кролика. Открытые ромбы: данные Choi et al. (1999) из клеток SAN кролика. Открытые перевернутые треугольники: данные Barbuti et al. (2007) из клеток SAN кролика.
Рис. 5. Зависимое от концентрации уменьшение продолжительности цикла кардиостимуляции (BCL) в процентах по ISO при различных условиях моделирования . ( A, B ) Boyett et al. и Kurata et al. модели центральных клеток ( C, D ) Boyett et al.и Kurata et al. модели периферических клеток. Черная сплошная линия: данные стандартной конфигурации модели, в которой эффекты ISO учтены для всех I CaL , I f , I Kr , I Ks , I st и Ca 2+ переходный. Черная пунктирная линия: данные модели без учета эффектов на I CaL . Серая сплошная линия: данные модели без учета эффектов на I f .Серая пунктирная линия: данные для модели, когда g Kr не подвержены влиянию ISO. Серая штрихпунктирная линия: данные модели, когда постоянная времени деактивации τ Kr не зависит от ISO. Черная пунктирная линия: данные модели без учета влияния ISO на I st .
Роль
I CaL
Роль I CaL в ISO-индуцированной положительной хронотропии была исследована путем сравнения эффекта ISO, когда его действие на I CaL было включено в модели, с эффектом, наблюдаемым, когда это действие было устранено.Зависимость изменения скорости от концентрации (когда I CaL не зависит от ISO, а I f , I Kr , I Ks , [Ca] i , и I st подвержены влиянию ISO) показано на рисунке 5 (черная пунктирная линия), а также показан стандартный ответ [полученный, когда были учтены все действия ISO (сплошная черная линия)]. В моделях центральных клеток, не влияя на I CaL , изменение скорости в зависимости от концентрации было аналогично стандартному ответу.Для моделей периферических клеток, когда I CaL не было изменено ISO, изменение скорости, зависящее от концентрации, всегда было больше стандартного значения во всем диапазоне рассматриваемых концентраций ISO. Это означает, что увеличение I CaL не внесло вклад в увеличение скорости ISO в наших условиях моделирования; действительно, во всяком случае, это снизило скорость.
Роль
I f
Изопреналин сдвигает зависимость кривой активации I f от напряжения в сторону более положительных мембранных потенциалов с максимальным сдвигом около +15 мВ в клетках SAN кролика (Zaza et al., 1996; Accili et al., 1997a, b; и примерно от +7 до +18 мВ в мышиных клетках SAN; Алиг и др., 2009; Baruscotti et al., 2010; Liao et al., 2010). Этот сдвиг приводит к увеличению перенапряжения I f , соответствующего диапазону потенциала кардиостимулятора. Приняв подход, аналогичный подходу, принятому выше для I CaL , роль I f в индуцированной ISO положительной хронотропии была исследована путем сравнения моделирования, включающего эффекты ISO на I f с теми в котором это действие не было включено.Смоделированные данные показаны на рисунке 5 (сплошная серая линия) и сравниваются со стандартными симуляциями (сплошная черная линия). И в моделях центральных, и в периферических клетках зависимые от концентрации изменения скорости были уменьшены по сравнению со стандартным значением, что позволяет предположить, что I f вносит вклад в индуцированную ISO положительную хронотропию.
Вычисленные потенциалы действия и I f в различных условиях представлены на рисунке 6, чтобы более четко проиллюстрировать роль I f в хронотропном эффекте ISO.На левой панели показаны результаты, рассчитанные на основе метода Boyett et al. модели, в то время как правые панели показывают результаты, рассчитанные на основе Kurata et al. модели. «Контрольные» следы были получены без моделирования эффектов ISO, следы с пометкой «ISO» были получены от включения всех воздействий 0,05 мкМ ISO, в то время как следы с пометкой «ISO, № I f shift» опускали действия на I f . В этих симуляциях ISO увеличил скорость кардиостимуляции за счет сокращения продолжительности цикла спонтанной активности как для центральной (рис. 6A, B), так и для периферической (рис. 6C, D) модели клеток.В случае «ISO, no I f shift» активность кардиостимуляторов обеих моделей замедляется, но менее чем на 3% по сравнению с условием «ISO». Положительный сдвиг в зависящей от напряжения активации для I f привел к значительному увеличению величины I f , как показано на нижних панелях на рисунках 6A, B (центральные модели) и рисунках 6C, D ( периферийные модели). Однако наши симуляции показывают, что увеличение I f не было исключительно результатом этого сдвига, поскольку, когда он отсутствовал, все еще наблюдалось увеличение I f , связанное с увеличением максимального диастолического давления. потенциал.
Рис. 6. Влияние индуцированного ISO увеличения I f на спонтанную активность . (A) Boyett et al. и (B) Kurata et al. модели центральных ячеек. (C) Boyett et al. и (D) Kurata et al. сотовые модели. На каждой панели показаны потенциалы действия (верхние кривые) и записи I f (нижние кривые) в контрольных условиях и при наличии 0.05 мкМ ISO. Показаны два моделирования с использованием ISO: в одном I f было затронуто, а в другом — нет. Это показывает, что роль I f в хронотропном эффекте ISO мала.
Роль
I Kr
Роль I Kr в хронотропном эффекте ISO состояла из трех компонентов: увеличение на г Kr , сдвиг кривой активации I Kr и увеличение кривой активации I Kr ставка отключения I Kr .Приняв подход к моделированию, можно было, в свою очередь, исследовать вклад каждого из этих элементов в действие ISO. Вычисленные зависимые от концентрации изменения спонтанной скорости центральной и периферийной моделей при исключении воздействия ISO на г Kr показаны на рисунке 5 (пунктирная серая линия) и в сравнении со стандартным моделированием ISO (сплошная черная линия ). В этих симуляциях Boyett et al. и Kurata et al. модели центральных ячеек ответили качественно аналогичным образом.Как в Boyett et al. и Kurata et al. центральные модели SAN (рис. 5A, B), устраняющие действие ISO на g Kr , ускоряли кардиостимуляцию, о чем свидетельствует большее уменьшение продолжительности цикла кардиостимуляции по сравнению со стандартным состоянием. Такой усиленный хронотропный эффект без модификации ISO г Kr также наблюдался в работе Boyett et al. модель периферических клеток (рис. 5C), хотя данные Kurata et al. Модель периферических клеток (рис. 5D) показала другой ответ.В Kurata et al. Модель периферических клеток, устранение воздействия ISO на г Kr привело к уменьшению продолжительности цикла кардиостимуляции по сравнению со стандартным значением.
При моделировании мы также вычислили зависящее от концентрации изменение спонтанной скорости, когда действие ISO на зависимость активации I Kr от напряжения было опущено, и сравнили результаты со стандартным значением (данные не показаны). В обоих исследованиях Boyett et al.и Kurata et al. В моделях центральных клеток вычисленные значения превышают стандартные значения, из чего следует, что сдвиг в зависимой от напряжения активации I Kr не способствует стимулированному ISO увеличению частоты стимуляции ритма (напротив, это фактически снизить скорость кардиостимуляции). Отсутствие этого действия ISO оказало незначительное влияние на реакцию моделей периферических клеток. Точно так же зависящее от концентрации изменение скорости стимуляции ритма при исключении воздействия ISO на τ Kr также показано на рисунке 5 (серая пунктирная линия).Как в центральных, так и в периферических моделях клеток устранение воздействия ISO на τ Kr оказало сильное влияние на хронотропный эффект ISO. В моделях центральных клеток, когда эффекты на τ Kr не учитывались, ISO снижал скорость стимуляции ритма (в большей степени в модели Boyett et al. По сравнению с моделью Kurata et al.). Подобные эффекты наблюдались для моделей периферических клеток, для которых ISO значительно замедлял скорость стимуляции ритма при высоких концентрациях. Результаты этого моделирования показывают, что влияние ISO на τ Kr играет важную роль в хронотропном эффекте ISO.
Базовая основа для влияния на спонтанную скорость изменения τ Kr со стороны ISO показана на рисунке 7. Здесь показаны потенциалы действия для Boyett et al. модели центральных (A) и периферических (C) клеток (левые панели) и Kurata et al. модели центральной ячейки (B) и периферийной ячейки (D) (правые панели). Следы, помеченные как «контроль», были получены в отсутствие ISO, следы, обозначенные «ISO», были получены при моделировании, в котором учитывались все эффекты 0,05 мкМ ISO.Следы с пометкой «τ Kr только затронуты» были получены в результате моделирования, в котором учитывалось только влияние 0,05 мкМ ISO на «τ Kr ». Также на рисунке в нижних панелях показаны соответствующие записи I Kr . В работе Boyett et al. при моделировании 0,05 мкМ ISO увеличивал частоту кардиостимуляции на 20 и 15% для моделей центральных и периферических клеток, соответственно. В обеих моделях ISO увеличивал максимальный диастолический потенциал (т.е. становился более отрицательным; см. Таблицу 1).Однако, когда был затронут только τ Kr , ISO увеличил скорость на 33 и 10% для моделей центральных и периферических клеток, соответственно, каждая с повышенным максимальным диастолическим потенциалом (см. Таблицу 1). По сравнению с «контрольным» состоянием, как в моделях центральной, так и в периферической клетке, ISO увеличилось I Kr во время начальной фазы реполяризации, но уменьшило I Kr в более поздней реполяризации и фазе деполяризации. Эти изменения в I Kr являются результатом изменения τ Kr .Когда было затронуто только τ Kr , наблюдалось увеличение I Kr в фазе ранней реполяризации, тогда как наблюдалось уменьшение I Kr в период поздней реполяризации и последующей фазы деполяризации. Снижение I Kr во время фазы деполяризации внесло значительный вклад в увеличение скорости кардиостимуляции. Подобные наблюдения были замечены с Kurata et al. модели (таблица 1). 0,05 мкМ ISO увеличивал скорость кардиостимуляции на 25% для модели центральной клетки и на 7% для модели периферической клетки.Когда затрагивался только τ Kr , ISO увеличивал скорость на 20% для модели центральной клетки и на 9% для модели периферической клетки.
Рис. 7. Эффекты увеличения постоянной времени дезактивации под действием ISO I Kr . Спонтанная активность в центральной части (A) Boyett et al. (B) Kurata et al. клеточные модели и периферийные устройства (C) Boyett et al. (D) Kurata et al.сотовые модели. На каждой панели показаны потенциалы действия и записи I Kr в контрольных условиях и в присутствии 0,05 мкМ ISO. Показаны два условия моделирования с использованием ISO — в одном было затронуто только τ Kr , а в другом были включены модулирующие эффекты на токи. Когда было затронуто только τ K , произошло существенное увеличение скорости стрельбы, но также было большое уменьшение максимального диастолического потенциала, что не наблюдается экспериментально.
Таблица 1 . Влияние ISO и вызванного ISO изменения скорости деактивации I Kr на характеристики потенциалов действия кардиостимулятора .
При моделировании мы также вычислили зависящее от концентрации изменение спонтанной скорости, когда действие ISO на увеличение г Ks было опущено. По сравнению с результатами, полученными в стандартных условиях, отсутствие воздействия ISO на g Ks оказало незначительное влияние на частоту стимуляции ритма (данные не показаны).
В предыдущем экспериментальном исследовании Lei et al. (2002) показали, что блокирование I Ks оказывает незначительное влияние на спонтанное количество клеток SAN кролика в контрольных условиях. Однако при применении 10 нМ ISO блокирование I Ks приводило к увеличению продолжительности цикла кардиостимуляции на ~ 10%. Это означает, что I Ks играет более важную роль в создании пейсмейкинга SAN при наличии ISO, чем при его отсутствии (Lei et al., 2002). Это экспериментальное наблюдение роли I Ks в потенциале стимуляции ритма SAN-клеток кролика было частично воспроизведено моделью. При моделировании блокирование I Ks в контрольных условиях приводило к незначительному изменению продолжительности цикла кардиостимуляции в обоих исследованиях Boyett et al. модели центральных и периферических клеток. При 10 нМ ISO блокирование I Ks приводило к увеличению продолжительности цикла кардиостимуляции на ~ 3 и 2% для модели центральной и периферической клетки, соответственно.Эти результаты моделирования качественно аналогичны экспериментальным наблюдениям Lei et al. (2002). Количественные различия между модельным моделированием и экспериментальными данными Lei et al. (2002) о роли I Ks в активности клеток SAN под контролем и в условиях ISO может быть связано с внутренними ограничениями модели, как подробно обсуждается в Zhang et al. (2000), или из-за ограничений в моделировании ISO (см. Обсуждение).
Роль
I st
Подобно другим исследованным токам, роль I st в положительном хронотропном эффекте ISO была исследована путем сравнения моделирования эффекта ISO, который включал его эффекты на I st , с теми, в которых это действие был опущен.Расчетные данные показаны на рисунке 5 (черная пунктирная линия), и их можно сравнить со стандартным откликом, включающим все эффекты ISO (сплошная черная линия). Путем исключения действия ISO на I st вычисленные дозозависимые изменения в Boyett et al. модели центральной клетки были лишь немного уменьшены по сравнению со стандартными значениями. В работе Boyett et al. В модели периферических клеток рассчитанные изменения скорости, зависящие от концентрации, также были близки к стандартным значениям.Эти результаты предполагают, что I st не играет важной роли в индуцированной ISO положительной хронотропии в условиях нашего моделирования. Поскольку в Kurata et al. Нет I st . клеточных моделей, роль увеличенного I st в положительном хронотропном эффекте ISO не анализировалась с использованием Kurata et al. сотовые модели.
Роль Ca
2+ Обработка и Na + -Ca 2+ Обмен
Экспериментальные исследования показали, что ISO увеличивает систолический и диастолический уровни внутриклеточного Ca 2+ ([Ca 2+ ] i ) (Ju and Allen, 1999; Vinogradova et al., 2002, 2008). Здесь мы рассмотрели, как индуцированные ISO изменения в I NaCa , последовавшие за вызванными ISO изменениями в [Ca 2+ ] i , способствуют положительному хронотропному действию ISO, если исключить влияние на другие токи. Результаты Boyett et al. модели показаны на рисунке 8. На рисунке временные кривые потенциалов действия (рисунки 8Ai, Bi), [Ca 2+ ] i (рисунки 8Aii, Bii) и I NaCa (рисунки 8Aiii) , Biii) накладывались в разных условиях.Следы, помеченные как «контроль», были получены в отсутствие ISO, следы, помеченные как «ISO», были получены при моделировании, в котором учитывались индуцированные ISO изменения систолического и диастолического [Ca 2+ ] i отдельно. В моделях как центральных, так и периферических клеток систолический и диастолический уровни [Ca 2+ ] i были удвоены по ISO. При моделировании с использованием метода Boyett et al. В моделях изменения в [Ca 2+ ] i увеличили скорость на 10% для центральной модели и на 1% для модели периферических клеток.Ускоренная частота кардиостимуляции была связана с увеличением I NaCa во время фазы диастолической кардиостимуляции (Рисунки 8Aiii, Biii), которое возникло из-за повышенного диастолического уровня [Ca 2+ ] i из-за комплексного действия одного увеличивал высвобождение SR Ca 2+ и уменьшал поглощение SR Ca 2+ . Дальнейшее моделирование также было выполнено для исследования индивидуальной роли повышенного высвобождения SR Ca 2+ и пониженного поглощения SR Ca 2+ .Либо увеличивая высвобождение SR Ca 2+ отдельно, либо уменьшая только поглощение SR Ca 2+ , диастолический уровень [Ca 2+ ] i был повышен, что привело к увеличению I NaCa что приводит к ускоренной стимуляции ритма. Результаты Kurata et al. клеточные модели были качественно подобны результатам Boyett et al. модели.
Рис. 8. Влияние индуцированных ISO изменений переходного процесса Ca 2+ на спонтанную активность .На левой панели показаны модели центральных клеток, а на правой панели показаны модели периферических клеток (модели Boyett et al.) Для потенциалов действия (Ai, Bi) , внутриклеточных Ca 2+ переходных процессов (Aii, Bii) и I. NaCa (Aiii, Biii) в контрольных условиях и в присутствии ISO. В моделировании ISO все эффекты, кроме модуляции внутриклеточного переходного процесса Ca 2+ , опускаются. Моделирование показывает, что увеличение переходного процесса Ca 2+ играет важную роль в ответе модели центральной клетки на ISO.
Обсуждение
В этом исследовании мы включили набор уравнений известных действий ISO на I CaL , I f , I Kr и I Ks , I st и внутриклеточный Ca 2+ в две установленные модели потенциала действия центральных и периферических клеток SAN кролика для имитации хронотропного эффекта ISO. Использование этих моделей позволило нам манипулировать действиями ISO и анализировать относительный вклад различных ионных токов в модулирующее действие β-адренергической стимуляции.
Смоделированный эффект ISO на спонтанную активность в моделях клеток SAN, изученных здесь, очень похож на эффект, наблюдаемый экспериментально на клетках SAN кролика (Zaza et al., 1996; Lei et al., 2000, 2002). ISO увеличил скорость кардиостимуляции и повлиял на форму потенциалов действия в обеих используемых здесь моделях. Вычисленное увеличение скорости, зависящее от концентрации, сопоставимо с экспериментальными данными, полученными от кроличьего SAN (Zaza et al., 1996; Lei et al., 2000). Как Boyett et al.(2001) и Kurata et al. (2008), ISO увеличивает выброс и максимальный диастолический потенциал, а также сокращает продолжительность потенциалов действия. Эти изменения в форме потенциала действия согласуются с экспериментальными наблюдениями (Zaza et al., 1996; Lei et al., 2000).
В нашем моделировании мы смогли показать, что увеличение I CaL способствует увеличению превышения потенциала действия с ISO, однако это очень мало влияет на ускорение скорости стрельбы.В модели периферических клеток, когда рассматривается действие увеличения I только CaL , скорость возбуждения фактически замедляется. В соответствии с этим было обнаружено, что блокирование I CaL нифедипином увеличивает скорость стимуляции ритма экспериментально в маленьких шариках, вырезанных с периферии SAN кролика (Kodama et al., 1997). Предположительно, лежащий в основе механизм связан с эффектом увеличения I CaL на продолжительность потенциала действия, в результате чего увеличение I CaL приводит к увеличению временного интервала между двумя последовательными потенциалами действия.
Роль I F в хронотропном эффекте ISO была спорной. I f рассматривается некоторыми как основной ток кардиостимуляции в SAN кролика (DiFrancesco, 1995, 2010; DiFrancesco and Borer, 2007) и повышается I f рассматривается как основная ионная основа хронотропный эффект ИСО (Zaza et al., 1996; Liao et al., 2010). Однако роль I f в хронотропии, опосредованной β-адренорецепторами, в SAN не является однозначной: Cai et al.(1995) показали, что положительный хронотропный эффект ISO не подвергается значительному влиянию, когда I f блокируется Cs + . Кроме того, в венозном синусе земноводных, в котором нет I f , ISO по-прежнему увеличивал скорость стрельбы (Ju and Allen, 1999), в то время как у мышей не хватало преобладающего гена канала I f (HCN4). проявляют физиологические реакции сердечного ритма на изопротеренол (Herrmann et al., 2007). В нашем моделировании мы заметили, что I f действительно способствует хронотропному эффекту ISO, но его вклад невелик, поскольку удаление действия ISO на I f приводит к небольшому изменению моделируемого соотношение концентрация-ответ для увеличения скорости.
Была также рассмотрена роль I st в хронотропии. В обеих моделях устранение воздействия ISO на I st не оказывает значительного влияния на дозозависимое увеличение скорости. При моделировании устранение действия ISO оказало относительно большее влияние на хронотропию в модели центральной клетки, чем в модели периферической клетки. Это связано с тем, что при разработке моделей из-за отсутствия экспериментальных данных о региональной разнице I st в SAN кролика была принята одинаковая величина (плотность тока) I st для обоих модели центральных и периферических клеток.Следовательно, модели центральных ячеек были относительно более чувствительны к I st из-за относительно меньшей плотности тока других токов деполяризующих каналов, таких как I f , I Na и I CaT по сравнению с моделями периферических клеток (Zhang et al., 2000).
В этом исследовании мы показали, что повышенный уровень [Ca 2+ ] i в симулированном присутствии ISO вносит вклад в увеличение скорости за счет уменьшения продолжительности цикла кардиостимуляции в модели центральной клетки.Это наблюдение качественно аналогично результатам Maltsev and Lakatta (2009), которые свидетельствуют о том, что «часы Ca 2+ » играют важную роль в создании синусоидальной узловой стимуляции сердца. Количественные различия между нашими результатами и результатами Maltsev and Lakatta (2009) по вкладу «Ca 2+ clock» в кардиостимуляцию, вероятно, отражают различия в параметризации различных токов, участвующих в мембранных часах, между Boyett et al. (2001) и модель Мальцева и Лакатта (2009).Параметры токов ионных каналов в Boyett et al. (2001) унаследованы от Zhang et al. (2000) клеточные модели, которые были основаны на экспериментальных данных и подтверждены ими (Zhang et al., 2000). Разница в ответе на повышенный [Ca 2+ ] i между центральным и периферическим Boyett et al. (2001) модели также отражали экспериментально наблюдаемую внутреннюю разницу в плотности тока некоторых деполяризующих токов между двумя разными типами клеток, как обобщено в Boyett et al.(2000) и Zhang et al. (2000).
Хронотропный эффект ISO отражает совместное воздействие ISO на ионные токи I CaL , I f , I Kr , I Ks и I 8 9000 , среди которых сравнительно более важную роль играет акция на I Kr . Устранение действия ISO на скорость дезактивации I Kr значительно уменьшило хронотропный эффект ISO как в Boyett et al.(2001) и Kurata et al. (2008) моделирует и даже обращает хронтропию ISO с высокой концентрацией в периферийной модели. Увеличение скорости дезактивации I Kr увеличивало величину I Kr в фазе ранней реполяризации потенциала действия, но уменьшало величину I Kr в фазе поздней реполяризации и фаза деполяризации. Уменьшение внешнего I Kr в период деполяризации позволяет чистому внутреннему току быть относительно большим, что ускоряет деполяризацию и, следовательно, скорость стимуляции ритма.
Важный вклад I Kr в кардиостимуляцию был отмечен в предыдущих экспериментальных исследованиях. В своей работе Sato et al. (2000) показали, что частичная блокировка I Kr ибутилидом (хотя ибутилид не является полностью избирательным блокатором каналов I Kr ) незначительно замедляет скорость стимуляции кардиостимуляции кролика, но полностью блокирует I кр. более высокая концентрация отменяет его потенциал действия, стимулирующего кардиостимуляцию.Оно и Ито (1995) также отметили важную роль чувствительного к E-4031 I Kr в стимуляции сердца SAN кролика. Дальнейшие исследования продемонстрировали, что частичная блокировка I Kr (около 50%) с помощью E-4031 практически устраняет спонтанную кардиостимуляционную активность в центральной ткани SAN кролика, хотя активность кардиостимуляции в периферической ткани SAN сохраняется (Kodama et al., 1999). . Данные по клеткам SAN мышей также показали важный вклад I Kr в потенциалы действия кардиостимуляции, поскольку блок I Kr по E-4031 увеличивал продолжительность цикла кардиостимуляции примерно на 68% (Nikmaram et al., 2008). Наши данные моделирования добавлены к этим экспериментальным данным, чтобы показать важную роль I Kr в нормальной кардиостимуляции SAN, особенно в выяснении роли измененной скорости деактивации I Kr в потенциалах действия кардиостимуляции, а именно нелегко получить из фармакологического исследования.
Допущения, ограничения и выводы
В настоящем исследовании из-за отсутствия доступных экспериментальных данных мы предположили, что действие ISO, зависящее от концентрации, на текущую амплитуду синусового предсердия I Ks и I st было таким же, как у I Kr .Это предположение требует пересмотра, когда станет доступно больше экспериментальных данных. Кроме того, там, где данные по клеткам SAN кролика не были доступны, использовались данные других видов (например, некоторые параметры уравнения для I st были основаны на множественных источниках данных из клеток SAN морских свинок и крыс (Guo et al. ., 1995, 1997; Shinagawa and Noma, 2000; Toyoda et al., 2005). Было обнаружено, что внутриклеточная цикличность Ca 2+ модулирует скорость кардиостимуляции в настоящей модели, но не в такой значительной степени, как в предыдущем исследовании, проведенном Мальцева. и Лакатта (2009).Поскольку роль часов Ca 2+ в стимуляции ритма SAN обсуждается (например, Honjo et al., 2003; Lancaster et al., 2004; DiFrancesco and Borer, 2007; Herrmann et al., 2007; Joung et al. , 2009; DiFrancesco, 2010; Gao et al., 2010; Himeno et al., 2011; Sosunov, Anyukhovsky, 2012), дальнейшая экспериментальная количественная оценка внутриклеточных механизмов обработки Ca 2+ клеток SAN и их вклада в электрогенез была бы полезна. . Также из-за ограниченной доступности экспериментальных данных о клетках SAN кролика, моделирование зависящего от концентрации действия ISO на I f было основано на экспериментальных данных Zaza et al.(1996), которые показали максимальный сдвиг установившейся кривой активации на +9,62 мВ. Хотя это относительно меньше по сравнению с вызванным ISO сдвигом (от +7 до +18 мВ) кривой активации I f в мышиных клетках SAN (Alig et al., 2009; Baruscotti et al., 2010 ; Liao et al., 2010), это близко к наблюдаемым значениям 8,8 мВ (Accili et al., 1997a) и 5,3 мВ (Accili et al., 1997b) для 1 мкМ ISO-индуцированного сдвига в потенциалозависимой активации. из I f в клетках SAN взрослого кролика.Поскольку нет экспериментальных данных, показывающих другие вызванные ISO изменения кинетики I f , при моделировании наклон кривой установившейся активации и зависящая от напряжения постоянная времени процесса активации были приняты равными без изменений в состоянии ISO.
Хотя важно, чтобы потенциальные ограничения моделей, используемых в этом исследовании, были четко обозначены, все же вероятно, что результаты нашего исследования действительны, показывая, что хронотропный эффект ISO включает интегрированное действие ISO на I CaL I f , I st , I Kr , I Ks и [Ca 2+ ] i ) для соответствия предварительным экспериментальным данным Заза и др.(1996) и Lei et al. (2000). Наше исследование имеет важное значение, подчеркивая относительный вклад обработки I f , Ca 2+ и модуляцию кинетики дезактивации I Kr в общую реакцию на ISO.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана грантами на проекты Совета инженерных и физических исследований Великобритании (EP / J00958X / 1; EP / I029826 / 1), British Heart Foundation (FS / 08/021) и Фонда естественных наук Китая ( 61179009).
Список литературы
Абрамочкин Д. В., Кузьмин В. С., Сухова Г. С., Розенштраух Л. В. (2009). Модуляция последовательности активации синоатриального узла кролика ацетилхолином и изопротеренолом исследована с помощью метода оптического картирования. Acta Physiol. (Oxf.) 196, 385–394.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Accili, E. A., Redaelli, G., and DiFrancesco, D. (1997a). Дифференциальный контроль тока, активируемого гиперполяризацией (i (f)), путем стробирования цАМФ и ингибирования фосфатазы в миоцитах сино-предсердного узла кролика. J. Physiol. (Лондон) 500 (Pt 3), 643–651.
Аччили, Э. А., Робинсон, Р. Б., и ДиФранческо, Д. (1997b). Свойства и модуляция I f у новорожденного по сравнению с сердечным узлом SA у взрослых. г. J. Physiol. 272 (3 Pt 2), h2549 – h2552.
Алиг, Дж., Марджер, Л., Месирка, П., Эмке, Х., Мангони, М. Э. и Исбрандт, Д. (2009). Контроль ЧСС по цАМФ чувствительности каналов HCN. Proc. Natl. Акад. Sci. США 106, 12189–12194.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Барбути, А., Терраньи, Б., Бриоски, К., и ДиФранческо, Д. (2007). Локализация f-каналов в кавеолах опосредует специфическую модуляцию скорости бета2-адренорецептора в синоатриальных миоцитах. J. Mol. Клетка. Кардиол. 42, 71–78.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Барускотти, М., Букки, А., Вискоми, К., Манделли, Г., Консалес, Г., Некки-Рускони, Т., Монтано, Н., Казали, К. Р., Микелони, С., Барбути, А. ., и ДиФранческо, Д. (2010). Глубокая брадикардия и блокада сердца, вызванные индуцибельным кардиоспецифическим нокаутом гена канала кардиостимулятора Hcn4. Proc. Natl. Акад. Sci. США 108, 1705–1710.
CrossRef Полный текст
Бойетт, М.Р., Чжан, Х., Гарни, А., Холден, А. В. (2001). Контроль активности кардиостимулятора внутриклеточным Ca 2+ : эксперименты и моделирование. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 359, 1091–1110.
Букки А., Барускотти М., Робинсон Р. Б. и ДиФранческо Д. (2003). I (f) -зависимая модуляция частоты кардиостимуляторов, опосредованная цАМФ, в присутствии рианодина в клетках сино-предсердного узла кролика. J. Mol. Клетка. Кардиол. 35, 905–913.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Цай, К., Лей, М., и Браун, Х. Ф. (1995). Ответы узла / предсердия СА морской свинки на ацетилхолин и адреналин в присутствии блокаторов i f и i K, ACh . J. Physiol. (Лондон) 483, 21.
Чой, Х. С., Ван, Д. Ю., Ноубл, Д., и Ли, К. О. (1999). Влияние изопреналина, карбахола и Cs + на активность Na + и потенциал кардиостимулятора в клетках узла SA кролика. г. J. Physiol. 276 (1 Pt 2), h305 – h314.
Pubmed Реферат | Pubmed Полный текст
Duchatelle-Gourdon, I., Hartzell, H.C., and Lagrutta, A.A. (1989). Модуляция калиевого тока замедленного выпрямителя в кардиомиоцитах лягушки бета-адренергическими агонистами и магнием. J. Physiol. (Лондон) 415, 251–274.
Pubmed Реферат | Pubmed Полный текст
Фриман, Л. К., и Касс, Р. С. (1993).Калиевые каналы замедленного выпрямления в желудочке и синоатриальном узле морской свинки: молекулярные и регуляторные свойства. Cardiovasc. Наркотики Ther. 7 (Приложение 3), 627–635.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Гао, З., Чен, Б., Джоинер, М.Л., Ву, Ю., Гуан, X., Коваль, О.М., Чаудхари, А.К., Кунья, С.Р., Молер, П.Дж., Мартинс, Дж. Б., Сонг, Л.С., и Андерсон, Мэн (2010). Высвобождение I (f) и SR Ca (2 + ) вносит вклад в активность кардиостимулятора в клетках синоатриального узла собак. J. Mol. Клетка. Кардиол. 49, 33–40.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Джайлс В., Накадзима Т., Оно К. и Шибата Э. Ф. (1989). Модуляция изопреналином тока запаздывающего выпрямителя K + в миоцитах предсердий лягушки быка. J. Physiol. (Лондон) 415, 233–249.
Pubmed Реферат | Pubmed Полный текст
Гуо, Дж., Оно, К., и Нома, А. (1995). Устойчивый внутренний ток, активированный в диапазоне диастолического потенциала в клетках сино-предсердного узла кролика. J. Physiol. (Лондон) 483 (Pt 1), 1–13.
Pubmed Реферат | Pubmed Полный текст
Хабучи, Ю., Хан, X., и Джайлз, В. Р. (1995). Сравнение токов выпрямителя, активируемого гиперполяризацией, и токов с задержкой в атриовентрикулярном узле кролика и синоатриальном узле. Heart Vessels Suppl. 9, 203–206.
Хагивара, Н. И. Х. (1989). Модуляция внутриклеточным Ca 2+ активируемых гиперполяризацией внутренних токов в клетках одиночных сино-предсердных узлов кролика. J. Physiol. (Лондон) 409, 121–141.
Pubmed Реферат | Pubmed Полный текст
Херрманн, С., Штибер, Дж., Штокл, Г., Хофманн, Ф. и Людвиг, А. (2007). HCN4 обеспечивает «резерв деполяризации» и не требуется для ускорения сердечного ритма у мышей. EMBO J. 26, 4423–4432.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Химено, Ю., Тойода, Ф., Сато, Х., Амано, А., Ча, К. Ю., Мацуура, Х., и Нома, А. (2011). Незначительный вклад цитозольных транзиентов Ca 2+ в ритм кардиостимулятора в клетках синоатриального узла морской свинки. г. J. Physiol. Heart Circ. Physiol. 300, h351 – h361.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Хондзё, Х., Инада, С., Ланкастер, М.К., Ямамото, М., Нива, Р., Джонс, С.А., Сибата, Н., Мицуи, К., Хориучи, Т., Камия, К., Кодама , I., и Boyett, MR (2003). Высвобождение Са 2+ саркоплазматического ретикулума не является доминирующим фактором в активности водителя ритма синоатриального узла. Circ. Res. 92, e41 – e44.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ито, Х., Оно, К. (1995). Быстро активирующийся канал выпрямителя с задержкой K + в клетках синоатриального узла кролика. г. J. Physiol. 269 (2 Pt 2), h543 – h552.
Pubmed Реферат | Pubmed Полный текст
Джунг, Б., Тан, Л., Маруяма, М., Хан, С., Чен, З., Стаки, М., Джонс, Л. Р., Фишбейн, М. К., Вайс, Дж.Н., Чен П. С. и Лин С. Ф. (2009). Внутриклеточная динамика кальция и ускорение синусового ритма при бета-адренергической стимуляции. Тираж 119, 788–796.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Джу, Ю. К., и Аллен, Д. Г. (1999). Как бета-адренергическая стимуляция увеличивает частоту сердечных сокращений? Роль внутриклеточного высвобождения Ca 2+ в пейсмекерных клетках амфибий. J. Physiol. (Лондон) 516 (Pt 3), 793–804.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ке Ю., Лей, М., Коллинз, Т. П., Ракович, С., Маттик, П. А., Ямасаки, М., Броди, М. С., Террар, Д. А., и Соларо, Р. Дж. (2007). Регуляция кальциевого канала L-типа и задержанная активность калиевого канала выпрямителя с помощью p21-активированной киназы-1 в пейсмекерных клетках синоатриального узла морской свинки. Circ. Res. 100, 1317–1327.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Харче, С., Ю, Дж., Лэй, М., и Чжан, Х. (2011). Математическая модель потенциалов действия клеток синоатриального узла мыши с молекулярными основаниями. г. J. Physiol. Heart Circ. Physiol. 301, H945 – H963.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Кодама И., Бойетт М. Р., Никмарам М. Р., Ямамото М., Хондзё Х. и Нива Р. (1999). Региональные различия в эффектах E-4031 в синоатриальном узле. г. J. Physiol. 276, H793 – H802.
Pubmed Реферат | Pubmed Полный текст
Кодама И., Никмарам М. Р., Бойетт М. Р., Сузуки Р., Хондзё Х. и Оуэн Дж. М. (1997). Региональные различия в роли токов Ca 2+ и Na + в активности водителя ритма в синоатриальном узле. г. J. Physiol. 272 (6 Pt 2), h3793 – h3806.
Pubmed Реферат | Pubmed Полный текст
Курата Ю., Мацуда Х., Хисатом И. и Шибамото Т. (2008).Региональные различия в динамических свойствах пейсмейкинга синоатриального узла: роль тока канала Na + . Biophys. J. 95, 951–977.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ланкастер, М. К., Джонс, С. А., Харрисон, С. М., и Бойет, М. Р. (2004). Внутриклеточный Ca 2+ и кардиостимуляция в синоатриальном узле кролика: неоднородность роли и контроля. J. Physiol. (Лондон) 556 (Pt 2), 481–494.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Лей, М., и Браун, Х. Ф. (1996). Две составляющие калиевого тока запаздывающего выпрямителя, IK, в клетках сино-предсердного узла кролика. Exp. Physiol. 81, 725–741.
Pubmed Реферат | Pubmed Полный текст
Лей, М., Купер, П. Дж., Камеллитти, П., и Коль, П. (2002). Роль 293b-чувствительного, медленно активирующегося выпрямителя тока калия, i (Ks), в пейсмекерной активности изолированных клеток синоатриального узла кролика. Cardiovasc. Res. 53, 68–79.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ляо, З., Локхед, Д., Ларсон, Э. Д., и Проенца, К. (2010). Фосфорилирование и модуляция активированных гиперполяризацией каналов HCN4 протеинкиназой А в синоатриальном узле мыши. J. Gen. Physiol. 136, 247–258.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Мальцев В.А., Лакатта Э.Г. (2009). Синергизм связанных субсарколеммальных часов Ca 2+ и сарколеммальных часов напряжения обеспечивает надежную и гибкую функцию кардиостимулятора в новой модели кардиостимулятора. г. J. Physiol. Heart Circ. Physiol. 296, H594 – H615.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Никмарам М. Р., Лю Дж., Абдельрахман М., Добжински Х., Бойетт М. Р. и Лей М. (2008). Характеристика эффектов рианодина, ТТХ, E-4031 и 4-AP на синоатриальные и атриовентрикулярные узлы. Прог. Биофиз. Мол. Биол. 96, 452–464.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Оно, К., Ито, Х. (1995). Роль быстро активирующегося выпрямителя с задержкой K + в активности кардиостимулятора синоатриального узла. г. J. Physiol. 269, h553 – H62.
Pubmed Реферат | Pubmed Полный текст
Сато Н., Танака Х., Хабучи Ю. и Джайлз В. Р. (2000). Электрофизиологические эффекты ибутилида на K (+) тока замедленного выпрямителя в клетках синоатриального и атриовентрикулярного узла кролика. Eur. J. Pharmacol. 22, 281–288.
CrossRef Полный текст
Шеннон Т. Р., Ван Ф. и Берс Д. М. (2005). Регуляция высвобождения Са в саркоплазматическом ретикулуме сердца за счет просвета [Са] и измененного стробирования оценивается с помощью математической модели. Biophys. J. 89, 4096–4110.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Шеннон, Т. Р., Ван, Ф., Пуглиси, Дж., Вебер, К., и Берс, Д. М. (2004).Математическая обработка интегрированной динамики Ca в миоците желудочка. Biophys. J. 87, 3351–3371.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Синагава, Ю.С. Х., и Нома, А. (2000). Устойчивый внутренний ток и внутренний выпрямительный ток K + в пейсмекерных клетках диссоциировали от синоатриальных клеток крысы. J. Physiol. (Лондон) 523, 593–605.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Сосунов, Э.А., Анюховский Э. П. (2012). Дифференциальные эффекты ивабрадина и рианодина на активность кардиостимулятора в синусном узле собаки и волокнах Пуркинье. J. Cardiovasc. Электрофизиол . (в прессе). DOI: 10.1111 / j.1540-8167.2011.02285.x
CrossRef Полный текст
Тойода, Ф., Динг, В. Г., и Мацуура, Х. (2005). Ответы устойчивого внутреннего тока на вегетативные агонисты в пейсмекерных клетках сино-предсердного узла морской свинки. руб. J. Pharmacol. 144, 660–668.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Виноградова Т.М., Богданов К.Ю., Лакатта Э.Г. (2002). Бета-адренергическая стимуляция модулирует высвобождение Ca (2+) рианодинового рецептора во время диастолической деполяризации, чтобы ускорить активность кардиостимулятора в синоатриальных узловых клетках кролика. Circ. Res. 90, 73–79.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Виноградова Т.М., Сиренко С., Ляшков, А. Э., Юнес, А., Ли, Ю., Чжу, В., Ян, Д., Рукнудин, А. М., Сперджен, Х., Лакатта, Э. Г. (2008). Конститутивная активность фосфодиэстеразы ограничивает частоту спонтанных сокращений кардиостимуляторов, подавляя локальные выбросы Ca 2+ . Circ. Res. 102, 761–769.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ву, Ю., Гао, З., Чен, Б., Коваль, О. М., Сингх, М. В., Гуан, X., Хунд, Т. Дж., Кучке, В., Сарма, С., Грумбах, И. М., Веренс, X. Х., Молер, П. Дж., Сонг, Л. С., и Андерсон, М. Е. (2009). Кальмодулинкиназа II необходима для физиологии синоатриального узла. Proc. Natl. Акад. Sci. США 106, 5972–5977.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ядзава, К., и Камеяма, М. (1990). Механизм рецепторно-опосредованной модуляции отсроченного выходящего калиевого тока в желудочковых миоцитах морских свинок. Дж.Physiol. (Лондон) 421, 135–150.
Pubmed Реферат | Pubmed Полный текст
Заза А., Робинсон Р. Б. и ДиФранческо Д. (1996). Базальные ответы Ca 2+ L-типа и токов, активируемых гиперполяризацией, на вегетативные агонисты в сино-предсердном узле кролика.