Движения, двигательный корково-мышечный путь. Параличи : Farmf
ДВИЖЕНИЯ. ДВИГАТЕЛЬНЫЙ КОРКОВО-МЫШЕЧНЫЙ ПУТЬ
ДВИЖЕНИЯ И ИХ РАССТРОЙСТВА
ОСНОВНОЙ ПРОВОДЯЩИЙ ПУТЬ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
ПЕРИФЕРИЧЕСКИЙ ПАРАЛИЧ
ЦЕНТРАЛЬНЫЙ ПАРАЛИЧ
ПРИЗНАКИ ПОРАЖЕНИЯ РАЗЛИЧНЫХ ОТДЕЛОВ ОСНОВНОГО ДВИГАТЕЛЬНОГО ПУТИ
Движения — наиболее универсальные проявления жизнедеятельности. Они могут возникать по тину безусловного или условного рефлекса (от лат. reflexus — отражение) или представляют собой различной сложности произвольные и непроизвольные двигательные акты.
Рефлекторная деятельность является отраженной, т.е. осуществляется под влиянием определенных воздействий, исходящей из внешней и внутренней среды. Рефлексы впервые были описаны французским философом и естествоиспытателем Р.Декартом (Descartes R., 1598—1650). И.М.Сеченов (1829— 1905) признавал, что «все акты сознательной и бессознательной жизни по способу своего происхождения суть рефлексы», и считал, что рефлекс — основа нервной деятельности. Эту точку зрения разделяли И.П. Павлов (1849-1936) и В.М. Бехтерев (1857—1927), разработавшие учение об условных (по Павлову), или сочетательных (по Бехтереву) рефлексах.
Каждый рефлекс имеет свою рефлекторную дугу — цепь нейронов. Область локализации рецепторов, раздражение которой ведет к возникновению определенного рефлекса, называется рефлексогенной зоной этого рефлекса. От рефлексогенной зоны, возникшие вследствие ее раздражения, нервные импульсы распространяются вначале по центростремительной (афферентной) части рефлекторной дуги до области ее замыкания — того или иного отдела центральной нервной системы. Здесь импульсы после переработки переходят на центробежную (эфферентную) часть рефлекторной дуги, по которой они и достигают рабочего органа: мышцы или железы.
Рефлекторные дуги врожденных (биологических, безусловных) рефлексов замыкаются в спинном мозге и на различных уровнях ствола и промежуточного отдела головного мозга. Через их посредство на основании врожденных нервных связей осуществляются простые безусловные, а также такие более сложные рефлекторные акты, как пищевой, пассивно-оборонительный, половой и т.п.
Через их посредство на основании врожденных нервных связей осуществляются простые безусловные, а также такие более сложные рефлекторные акты, как пищевой, пассивно-оборонительный, половой и т.п.
Условные рефлексы формируются в процессе жизнедеятельности путем выработки временных связей на базе безусловных рефлексов. Рефлекторные дуги их замыкаются главным образом в коре больших полушарий головного мозга. Совокупность условных рефлексов обеспечивает многообразие сложных реакций организма в его взаимоотношениях с внешней средой.
В процессе формирования двигательного акта кора большого мозга постоянно получает информацию о ходе его выполнения благодаря существующей
обратной связи (обратной афферентации). Это позволяет мозгу контролировать рефлекторный акт и корригировать его на любом этапе. Признавая наличие обратных связей, следует отметить, что когда мы говорим о рефлекторной дуге, то допускаем упрощенную трактовку понятия, так как логичнее считать, что при обеспечении рефлекторного акта нервные импульсы движутся по «рефлекторным кольцам».
Произвольные двигательные акты могут характеризоваться особой сложностью. В их основе лежат не просто реакции на определенный сигнальный раздражитель, а стремление к удовлетворению потребностей (у человека и социальных потребностей) по определенной программе, составляемой в моле на основании обширного афферентного синтеза всех тех сигналов из внешнего мира и внутренней среды организма, которые поступают в мозг по различным чувствительным путям (зрительным, слуховым, тактильным, проприоцептив-ным и пр.), а также на основании использования аппаратов памяти, сохраняющих и воспроизводящих сведения о прошлом опыте.
Известно, что, приступая под влиянием определенных мотиваций к действию, человек обычно прогнозирует его результаты. В связи с этим П. К. Анохин выдвинул гипотезу о существовании физиологического аппарата предвидения и оценки результатов действия, который формируется на основе предшествующего опыта, мотиваций и избранной программы предстоящего действия, а также текущего афферентного синтеза. Этот физиологический аппарат, предназначенный для восприятия информации о намеченном и производимом действии, а также о его результатах и сопоставления их со сложившимся ранее прогнозом, П.К. Анохин (1898—1974) назвал акцептором результатов действия.
Этот физиологический аппарат, предназначенный для восприятия информации о намеченном и производимом действии, а также о его результатах и сопоставления их со сложившимся ранее прогнозом, П.К. Анохин (1898—1974) назвал акцептором результатов действия.
Совпадение или несовпадение прогнозируемых и реальных результатов действия оказывает влияние на характер сопутствующих действию эмоциональных реакций. При совпадении реальных результатов с прогнозом они обычно положительны, при несовпадении — отрицательны. Последнее обстоятельство является стимулом к коррекции производимых действий или к дополнительным действиям, направленным на достижение поставленной цели.
ПЕРИФЕРИЧЕСКИЙ ПАРАЛИЧ
Периферический паралич всегда является следствием поражения периферических двигательных нейронов и возникает как при поражении тел этих нейронов, расположенных в двигательных ядрах черепных нервов или в передних рогах сегментов спинного мозга, так и их аксонов, входящих в состав различных структур периферической нервной системы, а также при блокаде нервно-мышечных синапсов.
Избирательное поражение тел периферических двигательных нейронов характерно, в частности, для эпидемического полиомиелита и бокового амиотро-фического склероза. Поражение структур периферической нервной системы может быть следствием травмы, сдавления, инфекционно-аллергического поражения и т.д., ведущих к развитию радикулопатий, плексопатий, невропатий, моно- или полиневропатий. Несостоятельность нервно-мышечных синапсов, передающих нервный импульс с нервного окончания на мышцу с помощью медиатора ацетилхолина-Н, возникает при миастении, отравлении токсином ботулизма.
Для периферического, или вялого, паралича характерны следующие признаки.
- Полная обездвиженность.
- Атония. Резко выраженное снижение мышечного тонуса. Мышца становится вялой, тестообразной, аморфной, не реагирует на раздражители, лишена силы. При периферическом параличе конечности обычна избыточность пассивных движений в ее суставах.

- Арефлексия. Исчезает рефлекторная реакция парализованных мышц в ответ на их внезапное раздражение, в частности растяжение, например, при ударе по сухожилию мышцы (сухожильные, или миотатические, рефлексы). Отсутствуют все рефлекторные двигательные реакции, в том числе защитные движения.
- Атрофия. Если двигательный нейрон или его аксон погибают, то все связанные с ним мышечные волокна претерпевают глубокую денервационную атрофию. Со временем в связи с развитием атрофического процесса уменьшается масса денервированных мышц. В течение нескольких недель после травмы или начала болезни гипотрофия мышц может быть незаметной, однако в течение первых 4 мес денервированные мышцы теряют до 20—30% исходной массы, а в дальнейшем — до 70—80%.
- Реакция перерождения, или реакция дегенерации, — извращение реакции на раздражение электрическим током парализованной мышцы и нефункцио-нирующего нерва.

В соответствии со сформулированным в 1939 г. американским физиологом У. Кенноном (Cennon W., 1871 — 1945) законом денервации рецепторы денервированных мышечных волокон приобретают гиперчувствительность к возбуждающему или тормозящему действию химических веществ (продукты метаболизма, токсины, лекарственные препараты, нейротрансмиттеры), достигающих этих рецепторов гематогенным путем.
ЦЕНТРАЛЬНЫЙ ПАРАЛИЧ
Центральный паралич — следствие поражения как двигательной зоны коры мозга, так и любого отдела нервной системы, связывающего ее с периферическими двигательными нейронами. Наряду с поражением коры моторной зоны больших полушарий причиной центрального паралича может быть нарушение функций корково-ядерных, моно- и полисинаптических корково-спин-номозговых путей на уровне лучистого венца, колена и двух передних третей заднего бедра внутренней капсулы, ножки мозга, ствола мозга, спинного мозга. В большинстве случаев патологический очаг вызывает поражение не всех структур двигательной зоны коры и корково-спинномозговых путей, к тому же часть этих путей не подвергается перекресту.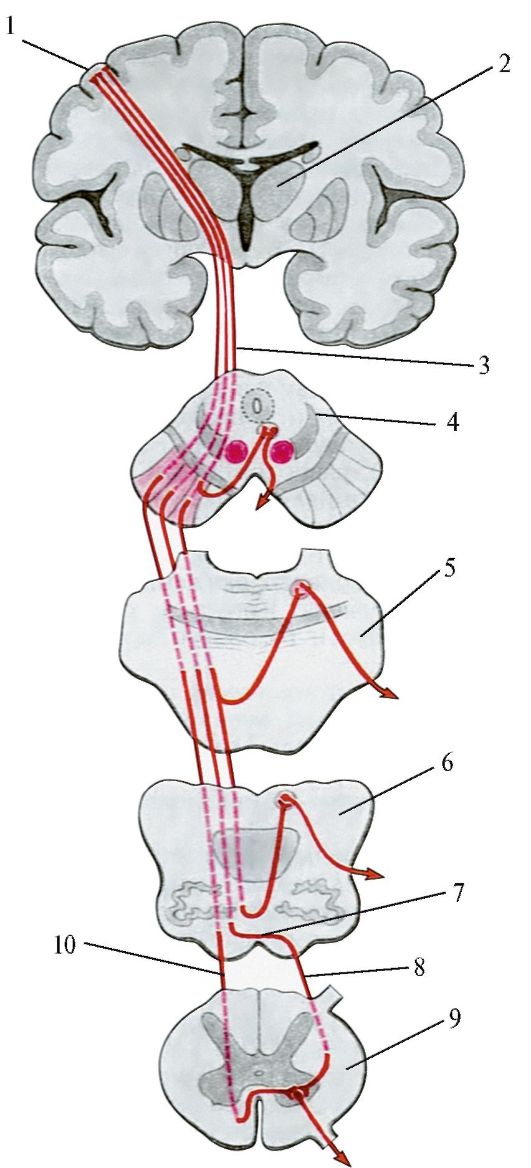 Поэтому на противоположной относительно патологического очага стороне тела никогда не поражаются все мышцы. Чаще в большей степени нарушена функция мышц кисти и плеча, затем — голени.
Поэтому на противоположной относительно патологического очага стороне тела никогда не поражаются все мышцы. Чаще в большей степени нарушена функция мышц кисти и плеча, затем — голени.
При центральном параличе сохранены и к тому же расторможены периферические мотонейроны, поэтому при нем, как правило, оказываются чрезмерно высокими сухожильные рефлексы, появляются патологические рефлексы и синкинезии.
Таким образом, центральный паралич — невозможность произвольных движений при сохранности и даже усилении интенсивности элементарных рефлекторных двигательных актов.
Для центрального, или спастического, паралича характерны следующие признаки.
- Повышение мышечного тонуса по контрактильному типу, или спастич-ностьмышц, обусловленная прекращением сдерживающих влияний центральных нейронов на периферические мотонейроны.
- Повышение сухожильных и надкостничных рефлексов, также сопряженное с нарушением сдерживающих влияний центральных нейронов на периферические мотонейроны.
- Возникновение клонусов, которые можно рассматривать как результат резко выраженной сухожильной гиперрефлексии, при этом однократное раздражение рефлексогенной зоны ведет к многократному ритмичному повторению стереотипного двигательного ответа.
- Патологические рефлексы, многие из которых вызываются у детей раннего возраста, а затем исчезают. При расторможенности периферических нейронов в связи с ослаблением сдерживающего влияния церебральных структур эти врожденные рефлексы появляются вновь и уже рассматриваются как патологические.
- Появление защитных рефлексов и патологических синкинезий (сопутствующих движений).
- В отличие от периферического паралича при центральном параличе отсутствует реакция перерождения (или дегенерации) мышц и нервов.
Спастичность мышц. Спастическое напряжение мышц, при котором мышечный тонус при движениях неравномерно нарастает и возможны симптомы складного ножа и отдачи, является характерным признаком поражения структур, связывающих центральные двигательные нейроны с периферическими. При остром поражении спастический тонус обычно развивается не сразу, некоторое время парализованные мышцы могут быть вялыми, однако сухожильные рефлексы при этом повышены, возможно раннее появление стопного патологического рефлекса Бабинского. Спастический тонус связан с чрезмерной активностью периферических мотонейронов, освободившихся от тормозящих воздействий структур ретикулярной формации мозгового ствола. Преимущественно в руках повышается тонус в мышцах-сгибателях, в ногах — в разгибателях. У больных с центральным гемипарезом обычно со временем формируются поза и походка Вернике—Манна.
При остром поражении спастический тонус обычно развивается не сразу, некоторое время парализованные мышцы могут быть вялыми, однако сухожильные рефлексы при этом повышены, возможно раннее появление стопного патологического рефлекса Бабинского. Спастический тонус связан с чрезмерной активностью периферических мотонейронов, освободившихся от тормозящих воздействий структур ретикулярной формации мозгового ствола. Преимущественно в руках повышается тонус в мышцах-сгибателях, в ногах — в разгибателях. У больных с центральным гемипарезом обычно со временем формируются поза и походка Вернике—Манна.
Клонусы. Как уже отмечалось, для центрального пареза или паралича характерно не только повышение сухожильных миотатических рефлексов, вызываемых и в норме, но нередко и появление многократной двигательной реакции на их вызывание (клонус). На практике чаще других вызываются следующие формы клонуса.
Рис. 4.8. Выявление клонуса стопы (а) и надколенника (б).
Клонус стопы вызывается следующим образом: больной лежит на спине, исследующий сгибает его ногу в коленном и тазобедренном суставах и, придерживая одной рукой его голень, другой рукой резко производит разгибание (тыльное сгибание) стопы, при этом желательно, чтобы больной слегка упирался подошвенной поверхностью стопы о ладонь обследующего. Ответная реакция — ритмичное чередование сгибания и разгибания стопы в течение всего периода, пока обследующий натягивает пяточное (ахиллово) сухожилие. При центральных парезах, особенно в случаях поражения пирамидных путей спинного мозга, клонус стопы иногда возникает спонтанно, если больной упирается передней частью стопы об пол или о спинку кровати (рис. 4.8а).
Рис. 4.9. Патологический рефлекс Бабинского (а) и его схема (б).
Клонус надколенника вызывается у больного, лежащего на спине с выпрямленными ногами. Обследующий большим и указательным пальцами захватывает верхний полюс надколенника и после этого резко смещает его в сторону голени и удерживает его в этом положении.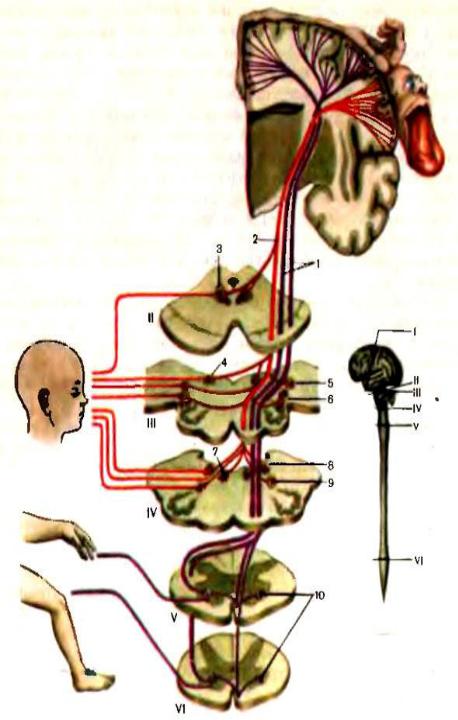 Происходит растяжение четырехглавой мышцы бедра и возникает ее ритмичное клоническое сокращение, не останавливающееся до тех пор, пока не будет прекращено растяжение мышечного сухожилия (рис. 4.86).
Происходит растяжение четырехглавой мышцы бедра и возникает ее ритмичное клоническое сокращение, не останавливающееся до тех пор, пока не будет прекращено растяжение мышечного сухожилия (рис. 4.86).
Значительно реже описанных форм клонуса удается выявлять клонус нижней челюсти, клонус кисти, клонус ягодичной мышцы.
Патологические рефлексы. В зависимости от места вызывания и характера ответной двигательной реакции патологические рефлексы могут быть разделены на стопные и кистевые, а также на разгибательные (экстензорные) и сгибательные (флексорные).
Рис. 4.10. Патологический рефлекс Оппенгейма.
При всех патологических стопных разгибательных рефлексах, особенно часто встречающихся при центральных парезах и параличах, ответной реакцией является разгибание большого пальца иногда в сочетании с веерообразным расхождением остальных пальцев стопы. К разгибательным патологическим рефлексам относятся следующие.
Рефлекс Бабинского — вызывается штриховым раздражением кожи наружного края подошвенной поверхности стопы, направленным от пяток к пальцам (рис. 4.9а, б). Является наиболее постоянным при центральных параличах и парезах, но надо иметь в виду, что у детей в возрасте до 2 лет он обычно вызывается и в норме.
Рефлекс Оппенгейма — вызывается проведением большим и указательным пальцами по гребню болынеберцовой кости с некоторым нажимом сверху вниз к голеностопному суставу (рис. 4.10).
Рефлекс Гордона — вызывается сдавлением трехглавой мышцы голени (рис. 4.11).
Рефлекс Шеффера — вызывается сдавливанием пяточного сухожилия (рис. 4.12).
При всех сгибательных стопных патологических рефлексах ответная двигательная реакция сводится к быстрому кивательному подошвенному сгибанию пальцев стопы. К сгибательным стопным патологическим рефлексам относятся следующие.
Рефлекс Россолимо — вызывается короткими легкими ударами пальцами руки исследующего по подошвенной поверхности концевых фаланг II—V пальцев стопы больного (рис.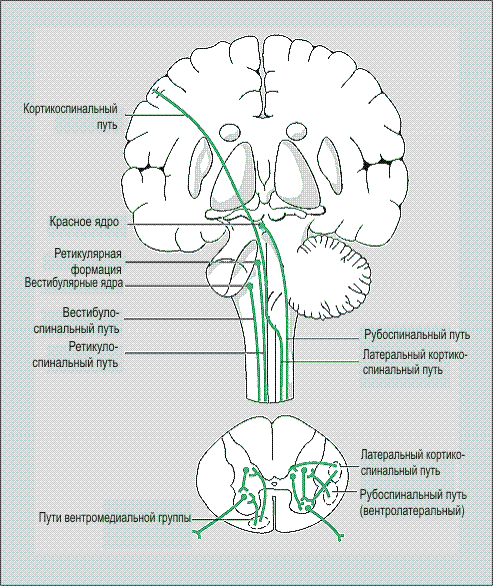 4.13).
4.13).
Рефлекс Бехтерева—Менделя — вызывается постукиванием молоточком по тылу стопы над III—IV плюсневыми костями.
Рефлекс Жуковского—Корнилова — вызывается постукиванием по центральной части подошвы (рис. 4.14).
При поражении пирамидных путей выше уровня шейного утолщения спинного мозга могут вызываться и кистевые патологические рефлексы, которые в основном можно рассматривать как аналоги стопных.
Рис. 4.11. Патологический рефлекс Гордона.
Рис. 4.12. Патологический рефлекс Шефера.
Рис. 4.13. Патологический рефлекс Россолимо.
Рис. 4.14. Патологический рефлекс Жуковского—Корнилова.
Защитные рефлексы. Как и патологические рефлексы, защитные рефлексы, или рефлексы спинального автоматизма, возникают вследствие изоляции от расположенных выше отделов мозга рефлекторных дуг, замыкающихся на уровне спинальных сегментов. Они представляют собой непроизвольные движения парализованных частей тела в ответ на их раздражение. Раздражителями при этом могут быть уколы, щипкообразные сдавления кожи, раздражение эфиром а т.п. (рис. 4.15а). Защитные рефлексы могут быть получены также в ответ на резкое пассивное движение в каком-либо суставе в связи с возникающим при этом раздражении проприорецепторов. Так, при пассивном подошвенном сгибании пальцев стопы парализованной ноги (рис. 4.15а, б) возникает подтягивание ноги в связи со спонтанным сгибанием ее в тазобедренном и коленном суставах (рефлекс Бехтерева—Мари—Фуа).
Рис. 4.15. Защитные рефлексы. а — эфирный метод, б — метод Бехтерева—Мари—Фуа.
При защитном бедренном рефлексе Ремака в ответ на штриховое раздражение кожи передней поверхности бедра возникает подошвенное сгибание стопы и пальцев. Защитные рефлексы могут возникать и при случайных раздражениях тела: прикосновении простыни, движении воздуха и т.д. В тех случаях, когда раздражитель остается незамеченным, такие рефлексы нередко называют «спонтанными» защитными рефлексами. Защитные рефлексы, как правило, возникают при сочетанном поражении корково-спинномозговых проводящих путей и экстрапирамидно-спинномозговых полисинаптических связей, бывают особенно отчетливыми при поражении пирамидных и экстрапирамидных путей на спинномозговом уровне. Уровень, ниже которого раздражение тканей ведет к возникновению защитных рефлексов, может способствовать выявлению нижнего полюса патологического очага в спинном мозге.
Защитные рефлексы, как правило, возникают при сочетанном поражении корково-спинномозговых проводящих путей и экстрапирамидно-спинномозговых полисинаптических связей, бывают особенно отчетливыми при поражении пирамидных и экстрапирамидных путей на спинномозговом уровне. Уровень, ниже которого раздражение тканей ведет к возникновению защитных рефлексов, может способствовать выявлению нижнего полюса патологического очага в спинном мозге.
Патологические синкинезии. Синкинезиями, или содружественными движениями, называются непроизвольные движения, присоединяющиеся к произвольным. Синкинезии могут быть физиологическими и наблюдаются у здоровых людей. Так, сжатию кисти в кулак обычно сопутствует разгибание в лучезапястном суставе, при ходьбе возникают сопутствующие движения рук (хейрокинез) и т.д.
При центральных парезах и параличах вследствие ослабления тормозного влияния на рефлекторные дуги, замыкающиеся на спинномозговом уровне, возникают предпосылки к развитию патологических синкинезии, которые могут быть глобальными, или спастическими, координационными и имитационными. Глобальные, или спастические, синкинезии — наблюдаются при спастических гемиплегиях. Попытки к активному движению больными конечностями или резкое напряжение мышц здоровой стороны тела вызывает непроизвольное сгибание в парализованной руке и разгибание — в ноге. Глобальные синкинезии в парализованной части тела могут возникать при общих эмоциональных реакциях, а также при зевоте, кашле, чиханье.
Координационные синкинезии — движения, которые больной не может выполнить изолированно, но при сложном двигательном акте они возникают как дополнительные и тогда не могут быть произвольно задержаны. Примером координационной синкинезии может быть симптом Раймиста, заключающийся в непроизвольном отведении и приведении парализованной ноги при попытке обследующего привести или отвести здоровую ногу больного, лежащего на спине и оказывающего при этом сопротивление обследующему. Другой пример координационной синкинезии — большеберцовый феномен Штрюм-пеля — заключается в том, что у больного, который не мог из-за центрального пареза ноги разгибать стопу, при попытке согнуть в колене больную ногу, преодолевая при этом сопротивление обследующего, возникает непроизвольное разгибание стопы, а иногда и большого пальца.
Другой пример координационной синкинезии — большеберцовый феномен Штрюм-пеля — заключается в том, что у больного, который не мог из-за центрального пареза ноги разгибать стопу, при попытке согнуть в колене больную ногу, преодолевая при этом сопротивление обследующего, возникает непроизвольное разгибание стопы, а иногда и большого пальца.
Рис. 4.16. Поза Вернике—Манна при центральном гемипарезе слепа.
Имитационные синкинезии — наблюдаются редко, обычно в случаях массивного поражения пирамидных и экстрапирамидных путей. При них в парализованной конечности возникают непроизвольные движения, тождественные тем, которые произвольно совершаются в симметричной здоровой конечности, например сгибание и разгибание пальцев, пронация и супинация предплечья и т.п.
ПРИЗНАКИ ПОРАЖЕНИЯ РАЗЛИЧНЫХ ОТДЕЛОВ ОСНОВНОГО ДВИГАТЕЛЬНОГО ПУТИ
Поражение различных отделов основного двигательного пути, состоящего из центральных и периферических нейронов и обеспечивающего возможность произвольных движений, имеет свои особенности, выявление которых способствует уточнению топики патологического очага.
• Поражение двигательной зоны коры большого полушария. Двигательная зона коры занимает предцентральную (переднюю центральную) извилину, главным образом поля 4 и 6, по Бродману, ее продолжение на медиальной поверхности полушария — парацентральную дольку, а также прилежащие территории лобной доли — так называемую предцентральную область (поле 8) и участок теменной доли (поля 5 и 7), а также поля 23с и 24с коры поясной извилины. Ввиду больших размеров площади двигательной зоны коры тотальное ее разрушение встречается редко. Обычно имеет место ее частичное поражение, что ведет к развитию двигательных расстройств в той части противоположной половины тела, которая проецируется на пораженный участок коры. Поэтому при корковой локализации патологического очага характерно развитие двигательных расстройств в ограниченной части противоположной половины тела: обычно они проявляются в форме монопареза или моноплегии. Поскольку на двигательную зону коры противоположная половина тела проецируется в перевернутом виде, расстройство функции, например верхних отделов правой предцен-тральной извилины, ведет к возникновению двигательных расстройств в левой ноге, а поражение нижних отделов левой двигательной зоны коры приводит к центральному парезу мышц правой половины лица и языка. Если патологический очаг расположен на уровне центральных извилин в межполушарной щели, например опухоль, растущая из большого серповидного отростка (фалькс-менингиома), пораженными могут оказаться примыкающие к серповидному отростку парацентральные дольки обоих полушарий, что ведет к развитию центрального нижнего парапареза, обычно в сочетании с нарушением контроля за тазовыми функциями.
Поскольку на двигательную зону коры противоположная половина тела проецируется в перевернутом виде, расстройство функции, например верхних отделов правой предцен-тральной извилины, ведет к возникновению двигательных расстройств в левой ноге, а поражение нижних отделов левой двигательной зоны коры приводит к центральному парезу мышц правой половины лица и языка. Если патологический очаг расположен на уровне центральных извилин в межполушарной щели, например опухоль, растущая из большого серповидного отростка (фалькс-менингиома), пораженными могут оказаться примыкающие к серповидному отростку парацентральные дольки обоих полушарий, что ведет к развитию центрального нижнего парапареза, обычно в сочетании с нарушением контроля за тазовыми функциями.
В случаях раздражения двигательной зоны коры большого полушария в мышцах соответствующей ей части противоположной половины тела могут
возникать судорожные пароксизмы, что характерно для фокальной эпилепсии джексоновского типа. Судороги эти обычно не сопровождаются расстройством сознания, но они могут распространяться на прилежащие части тела, иногда переходя во вторичный генерализованный судорожный припадок, который, начинаясь как очаговый, трансформируется в большой судорожный припадок с нарушением сознания. Если в патологический процесс вовлекается и прилежащая к пораженному участку передней центральной извилины зона задней центральной извилины, в части противоположной половины тела — мышцы которой находятся в состоянии пареза или паралича, возможны приступы парестезии — сенситивные джексоновские припадки, часто — гипестезия, при этом в большей степени нарушается проприоцептивная чувствительность и сложные виды чувствительности. При джексоновской эпилепсии во время припадка возможно сочетание локальных судорог и парестезии в определенной части тела на стороне, противоположной патологическому очагу.
Поражение дополнительной двигательной зоны в верхней теменной дольке (поля 5 и 7, по Бродману) может обусловить так называемый теменной парез на ограниченной территории противоположной половины тела, который обычно не сопровождается значительным повышением мышечного тонуса.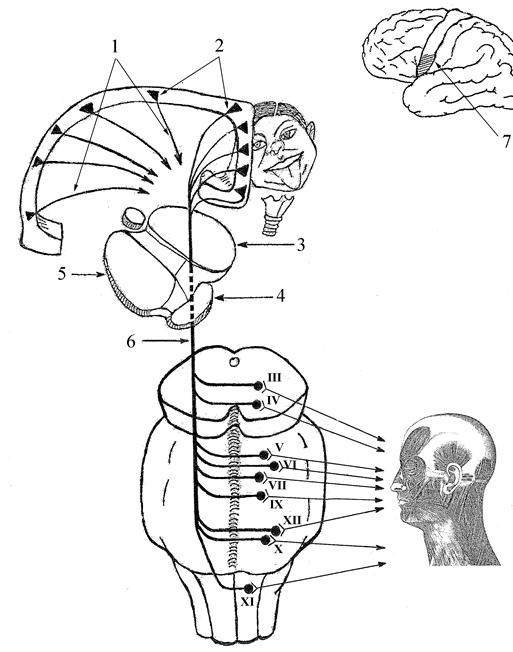
• Поражение лучистого венца. Лучистый венец представляет собой подкорковое белое вещество мозга, состоящее из аксонов нервных клеток, несущих импульсы в афферентном и эфферентном направлениях. При локализации патологического очага в лучистом венце на противоположной стороне обычно возникает центральный гемипарез, иногда в сочетании с гемигипестезией. Нарушения функции в различных участках противоположной половины тела выражены в разной степени, что зависит от того, какая часть лучистого венца оказалась вовлеченной в патологический процесс.
• Поражение внутренней капсулы. Во внутренней капсуле нервные волокна располагаются компактно, поэтому небольшой патологический очаг в зоне колена и двух передних третей переднего бедра внутренней капсулы может обусловить развитие центральной гемиплегии или центрального гемипареза на противоположной стороне. При более обширном патологическом процессе, распространяющемся на все заднее бедро внутренней капсулы, гемиплегия или гемипарез может сочетаться с возникающими на той же стороне гемианестезией и гемианопсией (выпадение гомоним-ных половин полей зрения), т.е. развивается так называемый синдром трех «геми». Остро возникающее поражение внутренней капсулы часто развивается при геморрагическом инсульте, проявляющемся медиальной внутримозговой гематомой.
При центральном гемипарезе на руке обычно в большей степени страдают мышцы, отводящие плечо, разгибатели и супинаторы предплечья, разгибатели кисти и пальцев, а на ноге — сгибатели бедра, разгибатели стопы и пальцев, что ведет к развитию у больных в период восстановительной фазы своеобразной позы, известной как поза Вернике—Манна (рис. 4.16). В связи с тем что в руке преобладает тонус мышц-сгибателей, а в ноге — разгибателей, рука, находящаяся в состоянии пареза, оказывается приведенной к туловищу и согнутой в локтевом суставе, кисть ее пронирована, а паретичная нога выпрямлена и кажется несколько длиннее здоровой ноги. Походка больных с центральным гемипарезом при этом оказывается своеобразной.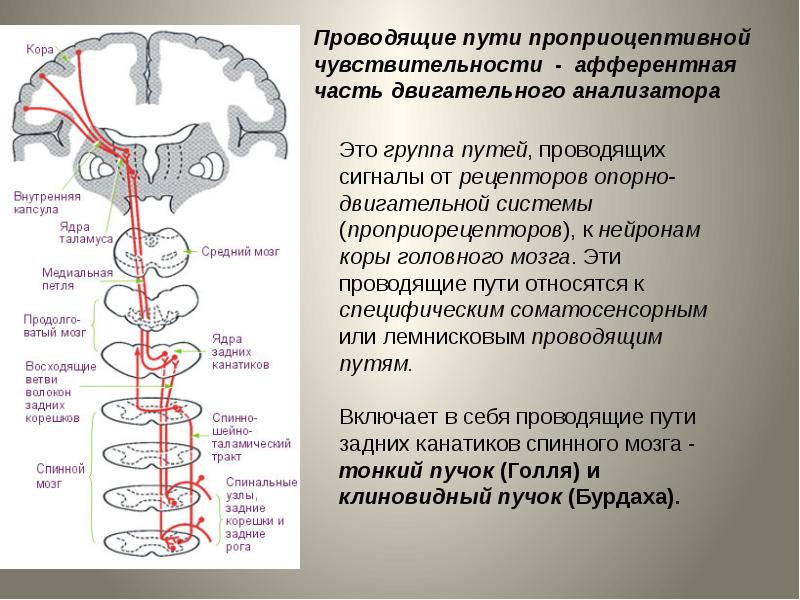 При ходьбе выпрямленная паретичная нога больного совершает движения по дуге, рука на стороне гемипареза остается согнутой и прижатой к туловищу. В таких случаях иногда говорят, что больной «рукой просит, а ногой косит».
При ходьбе выпрямленная паретичная нога больного совершает движения по дуге, рука на стороне гемипареза остается согнутой и прижатой к туловищу. В таких случаях иногда говорят, что больной «рукой просит, а ногой косит».
• Поражение ствола мозга. При одностороннем поражении различных отделов ствола мозга (средний мозг, мост, продолговатый мозг) характерно развитие альтернирующих (перекрестных) синдромов, при которых на стороне патологического очага возникают признаки поражения отдельных черепных нервов, а на противоположной стороне — гемипарез или гемиплегия по центральному типу, иногда — гемигипестезия. Вариант альтернирующего синдрома в таких случаях определяется уровнем и распространенностью поражения ствола. При двустороннем поражении ствола мозга функции черепных нервов могут быть нарушены с обеих сторон, при этом характерны псевдобульбарный или бульбарный синдромы, тетрапарез, расстройства чувствительности по проводниковому типу.
• Поперечное поражение половины спинного мозга — синдром Броун—Сека-ра. При поражении половины поперечника спинного мозга латеральный пирамидный путь вовлекается в патологический процесс ниже уровня его перекреста. В связи с этим центральный парез или паралич, возникающий ниже уровня поражения спинного мозга, развивается на стороне патологического очага. Двигательные расстройства при этом обычно сочетаются с нарушением чувствительности по проводниковому типу. В таких случаях на стороне патологического процесса оказывается нарушенной проприоцептивная чувствительность, а на противоположной стороне — поверхностная (болевая и температурная).
• Полное поперечное поражение спинного мозга в верхнем шейном отделе (С1— С4). При двустороннем поражении спинного мозга в верхнем шейном отделе возникает центральная тетраплегия, при этом сочетанное поражение с обеих сторон перекрещенных и неперекрещенных пирамидных путей ведет к тому, что страдают и мышцы туловища, включая дыхательную мускулатуру. Кроме того, в таких случаях ниже уровня расположения патологического очага обычно имеют место нарушения всех видов чувствительности по проводниковому типу, а также тазовые и трофические расстройства.
• Поражение шейного утолщения спинного мозга (C5—Th3). Поражение шейного утолщения спинного мозга также ведет к развитию тетраплегии в сочетании с нарушениями всех видов чувствительности по проводниковому типу ниже уровня патологического очага с тазовыми и трофическими расстройствами. Однако в связи с поражением шейного утолщения спинного мозга паралич или парез рук развивается по периферическому типу, тогда как паралич туловища и ног — по центральному.
• Поражение грудного отдела спинного мозга (Th4—Th22). Следствием поперечного поражения грудного отдела спинного мозга является спастическая нижняя параплегия в сочетании с утратой ниже уровня локализации патологического очага всех видов чувствительности, нарушением тазовых функций и расстройством трофики тканей.
• Поражение поясничного утолщения спинного мозга (L2—S2). При поражении поясничного утолщения спинного мозга развивается нижняя параплегия по периферическому типу в сочетании с нарушением чувствительности и трофики тканей на ногах и в аногенитальной зоне, а также с тазовыми расстройствами обычно в виде недержания мочи и кала.
• Избирательное поражение клеток передних рогов спинного мозга и двигательных ядер черепных нервов. В связи с избирательным поражением тел периферических двигательных нейронов возникает периферический паралич мыши, иннервацию которых они обеспечивают, при этом раздражение отдельных еще сохранившихся периферических мотонейронов может вызывать спонтанное сокращение мышечных волокон или их пучков (фибриллярные или фасцикулярные подергивания).
Избирательное поражение периферических двигательных нейронов характерно для эпидемического детского полиомиелита и бокового амиотрофичес-кого склероза, а также для спинальных амиотрофий.
• Поражение передних корешков спинного мозга. При поражении передних корешков спинного мозга характерны периферические параличи мышц, входящих в состав миотомов, одноименных пораженным корешкам.
• Поражение спинномозговых нервов. Следствием поражения спинномозговых нервов являются двигательные расстройства по периферическому типу в мышцах, иннервируемых аксонами мотонейронов, входящих в состав этих нервов, а также расстройства чувствительности (боль, гипалгезия, анестезия) в одноименных им дерматомах. Там же возможны вегетативные, в частности трофические, расстройства,
Следствием поражения спинномозговых нервов являются двигательные расстройства по периферическому типу в мышцах, иннервируемых аксонами мотонейронов, входящих в состав этих нервов, а также расстройства чувствительности (боль, гипалгезия, анестезия) в одноименных им дерматомах. Там же возможны вегетативные, в частности трофические, расстройства,
• Поражения нервных сплетений. Поражение нервного сплетения обусловливает развитие двигательных расстройств (паралич или парез) по периферическому типу, обычно в сочетании с нарушениями чувствительности и трофики в зоне иннервации периферических нервов, берущих начало из пораженного сплетения, или его части.
• Поражение периферического нерва. При поражении периферического нерва возникает периферический паралич или парез иннервируемых им мышц, обычно в сочетании с расстройством всех видов чувствительности и трофическими нарушениями в зоне иннервации пораженного нерва.
Экстрапирамидная система
Главная
›
Новости
Опубликовано: 28.12.2018
Экстрапирамидные пути — tr.tectospinalis, tr.rubrospinalis и т.д.
Кортико-мускулярный путь , рассмотренный в предыдущем разделе , обеспечивает произвольное сокращение той или иной мышцы. Однако отдельный законченный двигательный акт, каким бы примитивным он ни был, требует согласованного участия многих мышц.
Экстрапирамидная система: топография, строение, функции
Простейшее движение – поднимание руки – обеспечивается сокращением мышц плечевого пояса, но одновременно и мышц туловища и нижних конечностей, восстанавливающих правильное положение центра тяжести тела.
Качество движения зависит не только от вида и количества мышц, его реализующих. Нередко одни и те же мышцы участвуют в осуществлении различных движений, одно и то же движение может в зависимости от условий выполняться то быстрее, то медленнее, с большей или меньшей силой.
Проводящие пути #5: Экстрапирамидная система (рубро-, текто-, вестибуло- и ретикулоспинальный пути)
Таким образом, помимо пирамидной системы, обеспечивающей произвольное сокращение мышцы, выполнение движения требует участия механизмов, регулирующих последовательность, силу и длительность мышечных сокращений и регламентирующих выбор необходимых мышц.
Другими словами, двигательный акт формируется в результате последовательного, согласованного по силе и длительности включения отдельных нейронов и волокон кортико-мускулярного пути, отдающего приказы мышцам. Это включение обеспечивается при участии экстрапирамидной системы, прежде всего ее стрио-паллидарного отдела.
«Детская неврология», О.Бадалян
Периферический нейрон двигательного пути. Симптомы поражения.
Нужна помощь в написании работы?
ОСНОВНОЙ ПУТЬ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ.
Это двухнейронный путь, соединяющий кору больших полушарий мозга со скелетной (поперечнополосатой) мускулатурой (корково-мышечный путь). Различают центральный и периферический нейроны
этого пути. Центральный нейрон располагается в V слое (слой больших пирамидных клеток Беца) передней центральной извилины, в задних отделах верхней и средней лобных извилин и в парацентральной дольке. Существует четкое соматическое распределение этих клеток. Находящиеся в верхнем отделе прецентральной извилины и в парацентральной дольке клетки иннервируют нижнюю конечность и туловище, расположенные в средней ее части — верхнюю конечность. В нижней части этой извилины находятся нейроны, посылающие импульсы к лицу, языку, глотке, гортани, жевательным мышцам. Аксоны этих клеток в виде двух проводников (кортико-спинального пути (иначе называют пирамидным трактом) — от верхних двух третей передней центральной извилины и кортико — бульбарного пути — от нижнего отдела передней центральной извилины) идут от коры в глубь полушарий, проходят через внутреннюю капсулу (кортико-бульбарный путь — в области колена, а кортико-спинальный путь через передние две трети заднего бедра внутренней капсулы). Затем проходят ножки мозга, мост, продолговатый мозг и на границе продолговатого и спинного мозга кортико-спинальный тракт подвергается неполному перекресту. Большая, перекрещенная часть пути переходит в боковой столб спинного мозга и называется основным, или латеральным, пирамидным пучком. Меньшая неперекрещенная часть проходит в передний столб спинного мозга и носит наименование прямого неперекрещенного пучка. Волокна кортико-бульбарного пути заканчиваются в двигательных ядрах черепномозговых нервов (V, VII, IX, X, XI, XII), а волокна кортико-спинального пути — в передних рогах спинного мозга. При чем, волокна кортико-бульбарного тракта подвергаются перекресту последовательно, по мере подхода их к соответствующим ядрам черепных нервов («надъядерный» перекрест). Для глазодвигательных, жевательных мышц, мышц глотки, гортани, шеи, туловища и промежности существует двухсторонняя корковая иннервация, т. е. к части двигательных ядер черепных нервов и к некоторым уровням передних рогов спинного мозга волокна центральных двигательных нейронов подходят не только с противоположной стороны, но и со своей, обеспечивая, таким образом, подход импульсов из коры не только противоположного, но и своего полушария. Одностороннюю (только из противоположного полушария) иннервацию имеют конечности, язык, нижний отдел лицевой мускулатуры. Аксоны мотонейронов спинного мозга направляются к соответствующим мышцам в составе передних корешков, далее спинномозговых нервов, сплетений и, наконец, периферических нервных стволов.
Затем проходят ножки мозга, мост, продолговатый мозг и на границе продолговатого и спинного мозга кортико-спинальный тракт подвергается неполному перекресту. Большая, перекрещенная часть пути переходит в боковой столб спинного мозга и называется основным, или латеральным, пирамидным пучком. Меньшая неперекрещенная часть проходит в передний столб спинного мозга и носит наименование прямого неперекрещенного пучка. Волокна кортико-бульбарного пути заканчиваются в двигательных ядрах черепномозговых нервов (V, VII, IX, X, XI, XII), а волокна кортико-спинального пути — в передних рогах спинного мозга. При чем, волокна кортико-бульбарного тракта подвергаются перекресту последовательно, по мере подхода их к соответствующим ядрам черепных нервов («надъядерный» перекрест). Для глазодвигательных, жевательных мышц, мышц глотки, гортани, шеи, туловища и промежности существует двухсторонняя корковая иннервация, т. е. к части двигательных ядер черепных нервов и к некоторым уровням передних рогов спинного мозга волокна центральных двигательных нейронов подходят не только с противоположной стороны, но и со своей, обеспечивая, таким образом, подход импульсов из коры не только противоположного, но и своего полушария. Одностороннюю (только из противоположного полушария) иннервацию имеют конечности, язык, нижний отдел лицевой мускулатуры. Аксоны мотонейронов спинного мозга направляются к соответствующим мышцам в составе передних корешков, далее спинномозговых нервов, сплетений и, наконец, периферических нервных стволов.
Поможем написать любую работу на аналогичную
тему
Получить выполненную работу или консультацию специалиста по вашему
учебному проекту
Узнать стоимость
Поделись с друзьями
|
Произвольные движения (корковые программы моторных актов) реализуются через корково—мышечный путь. Остальные волокна (тонкие, с полисинаптической связью с мотонейронами) начинаются от клеток коры, имеющих отношение к экстрапирамидной иннервации. Последнее обстоятельство послужило поводом для пересмотра сложившихся (классических) представлений о генезе пирамидных расстройств. Считается, что на протяжении пирамидного пути «примесей» волокон и клеток другого вида нет только в корковом поле 4 и в области пирамид продолговатого мозга. Поражение этих отделов вызывает вялый паралич. Поражение всех других отделов пирамидного пути, содержащего экстрапирамидные волокна, приводит к развитию спастического паралича. Поэтому в развитии спастичности и гиперрефлексии ведущую роль усматривают в поражении экстрапирамидных волокон: при поражении пирамидных путей одновременно страдают тесно прилежащие ингибирующие (тормозные) экстрапирамидные волокна; активирующие мотонейроны передних рогов экстрапирамидные волокна вовлекаются в патологический процесс в меньшей мере, сохраняют свое влияние на а- и у-мотонейроны, вызывая спастичность и гиперрефлексию.
|
 Условно принято считать, что последний состоит из двух двигательных нейронов — центрального и периферического. Центральный двигательный нейрон (корково-спинномозговой путь, пирамидный путь) начинается, как было принято считать, от внутренних больших пирамидных нейронов («Я их назову колоссальными пирамидами…»,—писал ВА.Бец), расположенных в V слое коры предцентральной извилины и парацент-ральной дольки (описанной также ВА.Бецом), и заканчивается у моторных клеток передних рогов спинного мозга. Было установлено, что в пирамидных путях содержится 2 млн волокон, а внутренних больших пирамидных нейронов всего около 70 000, т. е. количество волокон от внутренних больших пирамидных нейронов в пирамидных путях (толстые миели-новые и быстропроводящие аксоны) не превышает 3—4%.
Условно принято считать, что последний состоит из двух двигательных нейронов — центрального и периферического. Центральный двигательный нейрон (корково-спинномозговой путь, пирамидный путь) начинается, как было принято считать, от внутренних больших пирамидных нейронов («Я их назову колоссальными пирамидами…»,—писал ВА.Бец), расположенных в V слое коры предцентральной извилины и парацент-ральной дольки (описанной также ВА.Бецом), и заканчивается у моторных клеток передних рогов спинного мозга. Было установлено, что в пирамидных путях содержится 2 млн волокон, а внутренних больших пирамидных нейронов всего около 70 000, т. е. количество волокон от внутренних больших пирамидных нейронов в пирамидных путях (толстые миели-новые и быстропроводящие аксоны) не превышает 3—4%.Пирамидная система | Детская неврология
Страница 12 из 51
Глава 3 ДВИЖЕНИЕ
ПИРАМИДНАЯ СИСТЕМА. ПУТИ И ЦЕНТРЫ. МЕТОДИКА ИССЛЕДОВАНИЯ.
ПУТИ И ЦЕНТРЫ. МЕТОДИКА ИССЛЕДОВАНИЯ.
ЦЕНТРАЛЬНЫЙ И ПЕРИФЕРИЧЕСКИЙ ПАРАЛИЧИ
Движение — одно из основных проявлений жизнедеятельности. Все важнейшие функции организма (дыхание, кровообращение, глотание, мочеиспускание, дефекация, перемещение тела в пространстве) реализуются в конечном счете движением, т. е. сокращением мышц. Мышечное сокращение — сложный биохимический и электрофизиологический процесс, в основе которого лежит феномен «скользящих нитей». Произвольность, свобода в выполнении или задержке тех или иных двигательных актов создают ложную видимость саморождаемости их, независимости от афферентных сигналов. Однако понять физиологию и патологию движений можно только при учете непрерывного потока центростремительных, афферентных импульсов, поступающих от рецепторного аппарата и сообщающих о необходимости осуществления того или иного двигательного акта, о ходе его выполнения, о своевременной коррекции силы, длительности и последовательности мышечных сокращений, о целесообразности прекращения движения и о его завершении.
Движение имеет рефлекторную природу. Рефлекторная дуга, обеспечивающая реализацию двигательного акта, может быть простейшей, двухнейронной (афферентная и эфферентная нервные клетки), или сложной, многосинаптической. В последнем случае не всегда удается провести четкую грань между афферентной и эфферентной частями рефлекторной дуги и определить начало той ее части, которая непосредственно руководит двигательным актом. Нарушение рефлекторной дуги в эфферентной ее части также может привести к значительным двигательным расстройствам. В то же время и сама чувствительность, афферентные системы находятся в зависимости от двигательных функций, стимулирующих рецепторный аппарат в ходе активного взаимодействия организма с внешней средой.
Периферический мотонейрон. Целостный двигательный акт является сложным рефлексом, формирующимся при участии многих систем, имеющих свои афферентные каналы, а также эфферентные средства доставки импульсов к исполнительному аппарату, непосредственно связанному с работающей мышцей, — к периферическому двигательному нейрону переднего рога спинного мозга и двигательных ядер черепных нервов.
Рис. 28. Передний рог спинного мозга.
А. Распределение мотонейронов в передних рогах спинного мозга на уровне шейного и поясничного утолщений.
Б. Соматическая проекция в переднем роге спинного мозга в шейном (а) н поясничном Сб) утолщениях.
Двигательные клетки переднего рога располагаются группами, ответственными за сокращение мышц туловища или конечностей, В внешейном и грудном отделах спинного мозга различают три группы клеток: переднюю и заднюю медиальные, обеспечивающие сокращение мышц туловища (сгибание и разгибание), и центральную, иннервирующую мышцу диафрагмы, плечевого и тазового поясов. В области шейного и поясничного утолщений к этим группам присоединяются передняя и задняя латеральные, иннервирующие мышцы сгибателей и разгибателей конечностей. Таким образом, в передних рогах на уровне шейного и поясничного утолщений имеется пять групп мотонейронов (рис. 28).
В каждой из групп клеток в переднем роге спинного мозга и в каждом двигательном ядре черепных нервов имеются три типа нейронов, выполняющих различную функцию. Альфа-большие клетки, проводящие двигательные импульсы с большой скоростью (60 — 100 м/с), обеспечивая возможность быстрых движений, связаны преимущественно с пирамидной системой (рис. 29). Альфа-малые нейроны получают импульсы от экстрапирамидной системы и оказывают позотонические влияния, обеспечивая постуральное (тоническое) сокращение мышечных волокон. Гамма-нейроны получают импульсы от ретикулярной формации. В отличие от альфа-нейронов гамма- нейрон отдает свой аксон не самой мышце, а заключенному в ней проприоцептору — нервно-мышечному веретену, влияя на его возбудимость.
Нервно-мышечное веретено — сложный проприоцептивный аппарат, представленный соединительнотканной капсулой, продольно «натянутыми» тонкими интрафузальными (внутриверетенными) мышечными волокнами, афферентными и эфферентными нервными окончаниями. Эфферентные нервные окончания гамма-нейронов иннервируют интрафузальные мышечные волокна и, таким образом, влияют на чувствительность нервно-мышечного веретена к внешним воздействиям (преимущественно со стороны экстрафузальных мышечных волокон — сокращение или расслабление мышцы) и косвенно на интенсивность афферентного потока импульсов от веретена к спинному мозгу.
Благодаря гамма-нейрону поддерживается и обеспечивается постоянство уровня сегментарно-рефлекторной деятельности: при дефиците внутрисегментарной импульсации гамма-нейрон повышает возбудимость веретена и увеличивает приток афферентных сигналов, при «перегрузке» сегментарного рефлекторного аппарата гамма-нейрон снижает чувствительность веретена и уменьшает афферентную импульсацию. Таким образом, гамма-нейрон не вызывает непосредственно мышечного сокращения, но обеспечивает на периферическом отрезке двигательных путей систему саморегуляции с обратной афферентацией, замыкающей кольцо рефлексов, поддерживающих мышечный тонус.
Пирамидный путь. Импульс произвольного мышечного сокращения приходит к альфа-большому нейрону из коры больших полушарий по пирамидному пути. Пирамидный путь начинается от клеток Беца, расположенных в V слое коры передней центральной извилины, задних отделов верхней и средней лобных извилин и парацентральной дольки. Двигательные центры мышц нижних конечностей находятся в верхних отделах передней центральной извилины, мышц верхних конечностей — в средних отделах. Ниже локализуются центры, иннервирующие мышцы шеи, лица, языка, глотки, гортани. Распределение двигательных центров в передней центральной извилине неравномерно. Наибольшее представительство имеют мышцы, выполняющие наиболее сложные, дифференцированные движения. Так, центры, обеспечивающие движение кисти, пальцев, занимают всю среднюю треть передней центральной извилины, тогда как массивные мышцы ног, выполняющие относительно простую, однообразную работу, имеют значительно меньшее представительство.
Гис. 29. Нервно-мышечное веретено.
1 — рецепторы кожи; 2 — задний корешок; 3 — тонкий пучок; 4 — передний корешок; 5 — волокно альфа- мотонейрона; 6 — концевые моторные пластинки альфа-мотонейрона; 7 — нервно-мышечное веретено; 8 — концевые моторные пластинки гамма-мотонейрона; 9 — экстрафузальные мышечные волокна; 10 — интрафузальные мышечные волокна; 11 — капсула веретена; 12,1а и 11 — афференты нервно-мышечного веретена; 13 — интернейроны спинного мозга; 14 — альфа-мотонейрон; 15 — гамма-мотонейрон.
1 — добавочное ядро глазодвигательного нерва; 2 — центральное хвостовое ядро глазодвигательного нерва; 3 — верхнее слюноотделительное ядро; 4 — нижнее слюноотделительное ядро; 5 — заднее ядро блуждающего нерва; 6 — ресничный узел; 7 — крылонебный узел; 8 — ушной узел; 9 — поднижнечелюстной узел; 10 — интрамуральные узлы внутренних органов; 11 — верхний шейный узел; 12 — шейно-грудной (звездчатый) узел; 13 — солнечное чревное сплетение; 14 — нижний брыжеечный узел.
Каждая клетка Беца отдает отросток, который в составе пирамидного пути доходит до соответствующего иннервируемого им сегмента спинного мозга, заканчиваясь синапсом с альфа-большим нейроном переднего рога спинного мозга или двигательных ядер черепных нервов. Из нижней трети передней центральной извилины волокна, участвующие в иннервации мышц лица, глотки и гортани, языка, заканчиваются у мотонейронов двигательных ядер черепных нервов и поэтому объединяются под названием корково-ядерного пути. Волокна из верхних и передней центральной извилины, участвующие в иннервации мышц туловища и конечностей, заканчиваются на мотонейронах передних рогов спинного мозга и объединяются под названием корково-спинномозгового пути. В связи с тем что сегментарной зоной верхних конечностей является шейное утолщение, а сегментарной зоной нижних конечностей — поясничное, волокна от средней трети передней центральной извилины заканчиваются преимущественно в шейном утолщении, а от верхней трети — в поясничном.
Собственно пирамидным путем обычно называют корково-спинномозговой путь, который образует пирамиды продолговатого мозга, однако принципиальной разницы между корково-спинномозговым и корково-ядерным путем нет. Оба они являются «корково-сегментарными путями» и объединяются в так называемую пирамидную систему (рис. 30).
Некоторые авторы объединяют их под названием «центральный двигательный путь». Выходя из передней центральной извилины на всем ее протяжении, аксоны клеток Беца лучеобразно сближаются (corona radiata) и компактно соединяются в узком пучке, проходящем в колене и передних и задней ножки внутренней капсулы.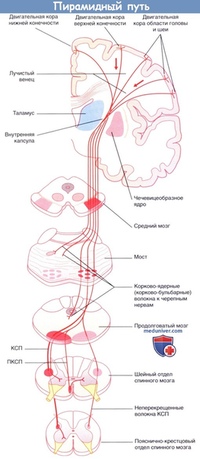
В колене проходят волокна, иннервирующие мышцы лица, глотки, гортани (корково-ядерный путь), в передней трети заднего бедра — мышцы рук, в средней трети — мышцы туловища и ног (корково-спинномозговой путь). Продолжая оставаться компактным, центральный двигательный путь переходит в ствол мозга. В среднем мозге его волокна проходят в основании ножек мозга, причем кнаружи располагаются волокна к мышцам ног, медиальнее — к мышцам рук и лица. В мосту центральный двигательный путь расчленяется, проходя узкими пучками между собственными ядрами моста и отдавая им коллатерали, а затем пирамидный путь вновь концентрируется в продолговатом мозге, образуя в его основании пирамиды. В стволе мозга корково-ядерный путь отдает волокна к двигательным ядрам черепных нервов ножек мозга, моста и продолговатого мозга, поэтому в пирамидах проходят волокна только корково-спинномозгового или пирамидного пути.
На границе продолговатого и спинного мозга большая часть волокон пирамидного пути образует перекрест (decussatio pyramidum) и идет в боковых канатиках спинного мозга. Меньшая, неперекрещенная часть волокон (пучок Тюрка) идет в передних канатиках спинного мозга. Перекрест осуществляется таким образом, что наружно расположенные в продолговатом мозге волокна, иннервирующие мышцы ног, после перекреста становятся медиальными, и, наоборот, волокна к мышцам рук, расположенные до перекреста медиально, становятся латеральными после перехода на другую сторону.
Рис. 30. Корково-спинномозговой и корково-ядерный путь.
I — Фронтальный срез головного мозга на уровне внутренней капсулы; II — средний мозг; III — мост; IV — продолговатый мозг; V — шейное утолщение спинного мозга; VI — поясничное утолщение спинного мозга
1 — корково-спинномозговой (пирамидный) путь; 2 — корково-ядерный путь; 3 — ядро глазодвигательного нерва; 4 — ядро отводящего нерва; 5 — двигательное ядро тройничного нерва; 6 — ядро лицевого нерва; 7 — ядро подъязычного нерва; 8 — двойное ядро; 9 — ядро добавочного нерва; 10 — мотонейроны переднего рога спинного мозга.
Таким образом, односторонний патологический процесс в области перекреста пирамиды может одновременно разрушить волокна к мышцам рук уже после их перекреста и волокна к мышцам ног до их перекреста.
В спинном мозге пирамидный путь (перекрещенный и неперекрещенный) отдает посегментарно волокна к альфа-большим нейронам переднего рога, осуществляющим непосредственную связь с работающей мышцей. Весь двигательный кортико-мускулярный путь двухнейронен: центральный нейрон — клетка Беца с длинным аксоном, образующим пирамидный корково-спинномозговой путь (а также корково-ядерный путь, заканчивающийся на альфа-больших нейронах двигательных ядер ствола мозга), и периферический нейрон — двигательная клетка переднего рога спинного мозга. Аксон периферического мотонейрона выходит из спинного мозга в составе переднего корешка, переходит в сплетения и периферические нервы, передавая нервный импульс мышечному волокну. Поражение периферического мотонейрона в переднем роге, переднем корешке, сплетении или периферическом нерве приводит к полному прекращению притока нервных импульсов к мышце, поступающих из коры по пирамидному пути, а также по рефлекторной сегментарной дуге от проприоцепторов. В этом случае мышца бездействует: отсутствуют и произвольные, и простейшие рефлекторные движения, так как поражена эфферентная часть сегментарной рефлекторной дуги.
Произвольные движения отсутствуют также при поражении центрального двигательного нейрона (клетки Беца и пирамидного пути) вследствие нарушения связи сегментарного двигательного аппарата, с корой. При этом повышается рефлекторная возбудимость периферического мотонейрона, освобожденного от регулирующих центральных влияний, обычно направляющих и корригирующих деятельность сегментарной рефлекторной дуги.
Таким образом, поражение любого участка всего» двигательного кортико-мускулярного пути ведет к полному или частичному нарушению произвольных движений. Полное отсутствие произвольных движений, обусловленное поражением кортико-мускулярного пути, называется параличом или плегией, ограничение объема движений и снижение силы — парезом.
Различают центральный и периферический параличи. Центральный паралич возникает при поражении центрального двигательного нейрона в любом его участке. Периферический паралич возникает при поражении периферического двигательного нейрона (клеток передних рогов, корешков и нервов). Паралич одной конечности именуют моноплегией, парез — монопарезом. Паралич двух конечностей носит название диплегии. Диплегии разделяются на параплегии, когда обездвижены две руки или две ноги, и на гемиплегии, характеризующиеся параличом руки и ноги на одной стороне. Паралич всех конечностей называется тетраплегией или квадриплегией. Встречаются также триплегии и трипарезы.
Центральный и периферический параличи сопровождаются характерными симптомокомплексами, клинически существенно отличаясь друг от друга прежде всего по уровню сегментарной рефлекторной активности, повышенной при центральном параличе и сниженной при периферическом.
При грубом поражении двигательного пути диагноз центрального и периферического паралича не представляет больших затруднений, однако в случаях легкого или начинающегося поражения кортико- мускуляторного пути необходимо детальное и всестороннее исследование двигательных функций, включающее осмотр, определение мышечного тонуса, мышечной силы, рефлексов и др.
При осмотре обращают внимание на положение больного (активное, пассивное, вынужденное), форму позвоночника (искривления могут быть следствием. поражения мышц), грудной клетки, кистей («когтистая кисть» при поражении локтевого нерва», «обезьянья кисть» при поражении срединного нерва), стоп («когтистая стопа» при поражении большеберцового нерва) и др. Мелкие мышечные подергивания — фибриллярные (сокращения отдельных мышечных волокон) или фасцикулярные (сокращения пучков волокон) — могут указывать на поражение периферического двигательного нейрона. Определяют конфигурацию мышц, их объем. Производят сравнительное измерение объема конечностей сантиметровой лентой на симметричных участках. Различают атрофии — уменьшение объема мышцы вследствие нарушения ее питания и гипертрофии — увеличение объема мышцы. Псевдогипертрофия — это разрастание жировой и соединительной ткани, создающее впечатление гипертрофии мышц. Иногда наблюдаются гемиатрофии (в частности, при гемиатрофии Ромберга — прогрессирующей атрофии костно-мышечной системы с одной стороны).
Различают атрофии — уменьшение объема мышцы вследствие нарушения ее питания и гипертрофии — увеличение объема мышцы. Псевдогипертрофия — это разрастание жировой и соединительной ткани, создающее впечатление гипертрофии мышц. Иногда наблюдаются гемиатрофии (в частности, при гемиатрофии Ромберга — прогрессирующей атрофии костно-мышечной системы с одной стороны).
О мышечной силе судят по сопротивлению, которое может оказать больной при пассивном движении в том или ином суставе, по объему активных движений, а также с помощью непосредственного измерения динамометром.
Оценку мышечной силы производят по пятибалльной системе. Полное отсутствие активных движений условно определяют как О, наличие минимальных движений, но невозможность преодолеть силу тяжести конечности — как 1 балл, способность преодолеть не только тяжесть конечностей, но и легкое сопротивление исследующего — 2 балла, способность при выполнении определенного движения преодолеть достаточное сопротивление обследующего — 3 балла, незначительное снижение мышечной силы — 4 балла. При сохранности двигательной функции мышечная сила оценивается в 5 баллов. Следует иметь в виду, что объем и сила движений могут быть ограничены вследствие патологии в суставах, мышцах, в связи с Рубцовыми изменениями кожи.
Для выявления слабости мышц конечностей используют пробы Мингаццини — Барре. Верхняя проба: обследуемому предлагают вытянуть руки вперед. При наличии мышечной слабости рука на стороне пареза опускается быстрее, чем на здоровой стороне. Нижняя проба: обследуемый, лежащий на спине, сгибает ноги в коленях под тупым углом, при этом на стороне пареза нога опускается быстрее. Имеется несколько модификаций пробы: проведение пробы в положении больного на животе (обследуемый сгибает ноги в коленных суставах и фиксирует их в этом положении — опускается «слабая» нога), использование дополнительных нагрузок и др.
Большое значение придается исследованию мышечного тонуса, который определяют после того, как обследуемый максимально расслабляется, снимет всякое активное напряжение мышц.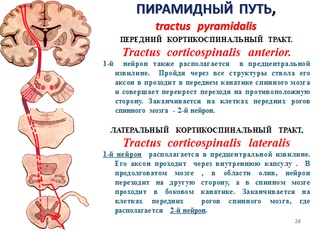 При этом проведение пассивных движений в различных суставах обнаруживает определенное непроизвольное сопротивление. Это сопротивление постоянно, не зависит от активных мышечных сокращений и не исчезает в полном покое. Мышечный тонус — рефлекс, имеющий свою рефлекторную дугу (гамма-нейрон переднего рога — проприоцептор — чувствительная биполярная клетка спинномозгового узла — альфа- малый нейрон переднего рога, отдающий импульс мышце и вновь гамма-нейрону, — проприоцептор — чувствительная клетка — альфа- малый нейрон и т.д. по обратной связи, создающей систему сегментарной саморегуляции). Мышечный тонус можно условно назвать «рефлексом на проприоцепцию», «ответом мышцы на самоощущение».
При этом проведение пассивных движений в различных суставах обнаруживает определенное непроизвольное сопротивление. Это сопротивление постоянно, не зависит от активных мышечных сокращений и не исчезает в полном покое. Мышечный тонус — рефлекс, имеющий свою рефлекторную дугу (гамма-нейрон переднего рога — проприоцептор — чувствительная биполярная клетка спинномозгового узла — альфа- малый нейрон переднего рога, отдающий импульс мышце и вновь гамма-нейрону, — проприоцептор — чувствительная клетка — альфа- малый нейрон и т.д. по обратной связи, создающей систему сегментарной саморегуляции). Мышечный тонус можно условно назвать «рефлексом на проприоцепцию», «ответом мышцы на самоощущение».
Относительное постоянство мышечного тонуса обеспечивается согласованной работой трех основных звеньев рефлекторной дуги: альфа-нейрона, гамма-нейрона и проприоцептора чувствительной клетки. Функциональная перегрузка альфа-малого нейрона, сопровождающаяся повышением мышечного тонуса; влияет на состояние гамма-нейрона, который повышает порог восприятия проприоцептора и таким образом снижает поток афферентных импульсов к альфа-малому нейрону. Альфа-малый нейрон «разгружается» и нормализует мышечный тонус. При расслаблении тонуса гамма-нейрон активизируется и усиливает афферентную «нагрузку» на альфа-малый нейрон.
В обеспечении мышечного тонуса принимают участие, помимо сегментарного аппарата, ретикулярная формация ствола, подкорковые узлы, мозжечок, кора больших полушарий.
При мышечной гипотонии, атонии (частичном снижении или полном отсутствии тонуса) непроизвольное напряжение мышц уменьшается или не ощущается совсем, мышцы становятся дряблыми, наблюдается «разболтанность» суставов с увеличением объема движений в них за счет переразгибаний (тоническое напряжение мышц в норме предохраняет суставы от повреждений). При выраженной атонии наблюдается симптом Оршанского — чрезмерное разгибание ноги в коленном суставе, верхней конечности в локтевом суставе. Снижение мышечного тонуса свидетельствует о поражении сегментарного рефлекторного аппарата, периферического двигательного нейрона. Мышечная гипотония может также наблюдаться при поражении ретикулярной формации, ее связей с мозжечком, стриопаллидарной системой.
Снижение мышечного тонуса свидетельствует о поражении сегментарного рефлекторного аппарата, периферического двигательного нейрона. Мышечная гипотония может также наблюдаться при поражении ретикулярной формации, ее связей с мозжечком, стриопаллидарной системой.
Мышечная гипертония проявляется в виде спастического повышения тонуса при центральном параличе, в виде пластической гипертонии (ригидности) при поражении паллидарной системы.
Повышение мышечного тонуса при пирамидном синдроме обусловлено перерывом центрального влияния на клетки переднего рога спинного мозга и растормаживанием сегментарного рефлекторного аппарата.
Рве. 31. Схема рефлекторных дуг и способы вызывания основных сухожильных и надкостничных рефлексов.
Исследование рефлексов позволяет судить не только о том, поражен центральный или периферический мотонейрон, но также об уровне поражения головного и спинного мозга. По типу рецепторов, с которых вызывается рефлекс, различают поверхностные рефлексы (с рецепторов мышц, сухожилий, надкостницы, суставов). Целью исследования рефлексов в неврологии является определение их повышения (гиперрефлексия), снижения (гипорефлексия) или полного отсутствия (арефлексия). Гиперрефлексия наблюдается (так же, как и спастическая гипертония мышц) при поражении центрального двигательного нейрона и нарушении связи сегментарного двигательного аппарата с корой больших полушарий, что приводит к повышению рефлекторной возбудимости периферического мотонейрона. Гипорефлексия и арефлексия свидетельствуют о поражении периферического двигательного нейрона.
Исследование рефлексов проводят «сверху вниз», начиная с рефлексов, замыкающихся на верхних сегментах мозга (зрачковый — средние отделы ножек мозга; надбровный, корнеальный, конъюнктивальный, мандибулярный — мост; глоточный, небный — продолговатый мозг) и заканчивая рефлексами с нижних сегментов спинного мозга (анальный — нижние крестцовые сегменты), поочередно «зондируя исправность» сегментов шейного, грудного, поясничного и крестцового отделов спинного мозга (рис.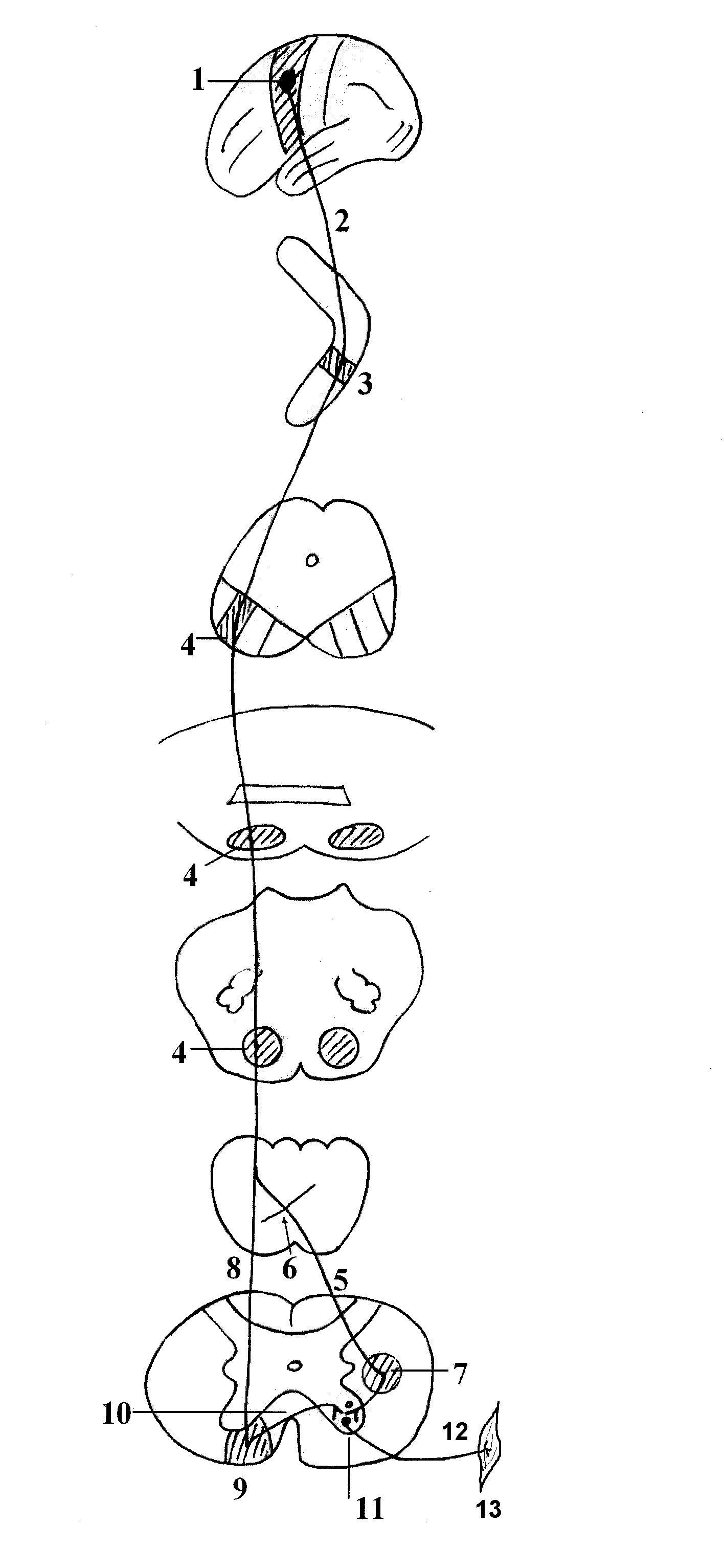 31).
31).
К рефлексам, определение которых входит в обязательный минимум исследования двигательных функций, относятся рефлексы со слизистых оболочек — корнеальный, конъюнктивальный, глоточный, небный, анальный; кожные рефлексы — брюшные (верхний, средний, нижний), кремастерный и подошвенный; сухожильные — нижнечелюстной, сгибательно-локтевой, разгибательно-локтевой, коленный, ахиллов; надкостничные — надбровный, пястно-лучевой, лопаточно-плечевой, реберно-абдоминальный, лобковый; суставные рефлексы — Лери и Майера (табл. 1). Сопоставление выраженности рефлексов с одной и другой стороны позволяют уловить тонкие односторонние нарушения двигательных функций (асимметрия рефлексов — анизорефлексия).
Периферический паралич возникает при поражении периферического двигательного нейрона в любом его участке (клетка переднего рога спинного или двигательных ядер ствола мозга, передний корешок, сплетение, периферический нерв). Основными симптомами периферического паралича являются арефлексия, мышечная атония и атрофия,
Таблица 1. Сегментарная локализация наиболее важных рефлексов
| Рефлекс | Рефлекторная дуга |
Зрачковый | Средний мозг (зрительный, глазодвигательный нервы) |
Корнеальный | Мост (тройничный, лицевой нервы) |
Мандибулярный | Мост (тройничный нерв) |
Глоточный | Продолговатый мозг (языкоглоточный, блуждающий нервы) |
Карпорадиальный | св-св |
С двуглавой мышцы | се-с. |
С трехглавой мышцы | с„-с7 |
Брюшные |
|
верхний | Th7 — Thg |
средний | The — Th20 |
нижний | Th21 — Th22 |
Кремастерный | L, — La |
Коленный | L2-L4 |
Ахиллов | Lb-Ss |
Подошвенный | Le-Ss |
Анальный | SB |
возникновение которых связано с поражением сегментарного рефлекторного аппарата.
Кроме этого, при периферическом параличе наблюдается нарушение электровозбудимости — реакция перерождения, которая является специфическим симптомом поражения периферического мотонейрона. В норме при раздражении гальваническим током катодозамыкательное сокращение (КЗС) больше анодозамыкательного сокращения (АЗС). При реакции перерождения КЗС — АЗС или АЗС > КЗС.Исследование электровозбудимости позволяет определить тяжесть поражения при периферическом параличе и прогноз.
Нередко при периферическом параличе наблюдаются фибриллярные или фасцикулярные мышечные подергивания, появление которых объясняют раздражением патологическим процессом еще не погибших нейронов. Фибриллярные и фасцикулярные подергивания обычно сопровождают такие атрофические парезы и параличи, которые являются результатом хронического прогрессирующего процесса в клетках периферических двигательных нейронов (переднего рога спинного мозга или двигательных ядер черепных нервов) или в передних корешках спинного мозга.
Помимо характерного симптомокомплекса периферического паралича (арефлексия, атония, атрофия и реакция перерождения), наблюдаемого при поражении периферического мотонейрона в любом его участке, имеются клинические симптомы, позволяющие определить локализацию этого поражения.
Для поражения переднего рога характерны раннее возникновение атрофий и реакции перерождения, преимущественное поражение проксимальных отделов конечностей, асимметрия атрофий, фибриллярные подергивания. Поражение переднего рога нередко имеет преимущественную локализацию в области той или иной группы нервных клеток, что определяет мозаичность распределения мышечных атрофий, нехарактерную для поражения переднего корешка, в котором волокна от клеток всего переднего рога идут компактным пучком.
Поражение передних корешков приводит к атрофии, арефлексии и атонии мышц в зоне иннервации корешков; отмечаются также фасцикулярные подергивания.
При поражении сплетений возникают двигательные, чувствительные и вегетативные расстройства в зоне иннервации данного сплетения.
Поражение периферических нервов сопровождается периферическим параличом, чувствительными и вегетативными расстройствами в зоне иннервации нерва, преимущественно в дистальных отделах конечности.
Центральный паралич (парез) возникает при поражении центрального двигательного нейрона в любом его участке — двигательной зоне коры больших полушарий, внутренней капсуле, стволе мозга и спинном мозге. Перерыв центральных влияний освобождает сегментарный рефлекторный аппарат, «обнажая», «растормаживая» заложенные в нем функциональные возможности. Для здоровых новорожденных характерно наличие некоторых симптомов, присущих 92 центральному параличу, что связано с еще непрочными и «незрелыми» корково-ядерными и, корково-спинномозговыми связями. Эта «незрелость» двигательных функций новорожденного имеет большое значение, являясь необходимой на этом этапе развития. Так, например, «патологические» симптомы орального автоматизма помогают ребенку найти, искать и захватить сосок матери, благодаря им ребенок «умеет» сосать. По-видимому, неправильно связывать двигательную «незрелость» новорожденных только с незаконченной миелинизацией пирамидного пути (которая завершается на 5-6-м месяце жизни). Здоровый новорожденный достаточно зрел для периода новорожденности, однако в дальнейшем, особенно на протяжении первых месяцев жизни, критерий зрелости будет постоянно меняться. Необходимые в период новорожденности мышечный гипертонус, гиперрефлексия, симптомы орального автоматизма и пр. становятся Помехой в дальнейшем развитии и расширении двигательных функций. Появляется необходимость в коррекции деятельности периферического мотонейрона, во включении цереброспинальных каналов доставки корригирующих импульсов.
Все основные симптомы центрального паралича так или иначе связаны с повышением возбудимости периферического сегментарного рефлекторного аппарата. Для центрального паралича характерны: 1) мышечная гипертония; 2) гиперефлексия и расширение рефлексогенной зоны; 3) клонусы стоп, коленных чашечек, кистей; 4) патологические рефлексы; 5) защитные рефлексы; 6) патологические синкинезии.
Кроме этого, наблюдаются симптомы, не связанные с Повышением сегментарной рефлекторной деятельности, но обусловленные сопутствующим поражением путей, сопровождающих корково-спинномозговой. К ним относятся отсутствие или снижение брюшных и кремастерных рефлексов и расстройства тазовых функций по центральному типу.
Мышечную силу исследуют при центральном параличе так же, как и при периферическом.
Мышечная гипертония. Мышцы напряжены, плотноваты на ощупь. Тонус повышен по спастическому типу, мышечное сопротивление при исследовании тонуса ощущается в начале движения (симптом «складного ножа»). При резко выраженной гипертонии возникают контрактуры. При гемиплегии (гемипарезе) тонус повышается в сгибателях руки и разгибателях ноги. Рука приведена и согнута, нога вытянута — лозе Вернике-Манна. Изменение мышечного тонуса определяет походку при гемипарезе: больной описывает ногой полукруг, чтобы не «цеплять» носком «удлиненной» ноги за пол. При спастическом парапарезе больной ходит на носках, скрещивая ноги. У детей 1-го года жизни начиная с 2-месячного возраста наличие спастических явлений можно установить следующим образом: если приподнять ребенка, то вместо сгибания нижних конечностей в коленных и тазобедренных суставах происходит их разгибание и скрещивание.
Повышение сухожильных и надкостничных рефлексов сопровождается расширением рефлексогенных зон. Например коленный рефлекс может вызываться со всей передней поверхности бедра и голени, ахиллов — с подошвы.
Рис. 32. Пирамидный синдром. Способы вызывания патологических рефлексов. Кистевые патологические рефлексы: 1 — аналог рефлекса Россолимо; 2 — рефлекс Жуковского; 3 — рефлекс Якобсона — Ласка. Разгибательные и сгибательные стопные патологические рефлексы: 4 — рефлекс Бабинского; 5 — рефлекс Оппенгейма; 6 — рефлекс Шеффера; 7 — рефлекс Гордона; 8 — рефлекс Россолимо; 9 — рефлекс Бехтерева 1; 10 — рефлекс Жуковского; 11 — рефлекс Бехтерева II. Способы вызывания основных патологических защитных рефлексов: 12 — проба Мари — Фуа
Клонусы стоп, коленных чашечек и кистей — ритмические сокращения мышц в ответ на растяжение сухожилий — являются следствием резкого повышения сухожильных рефлексов. При грубых поражениях пирамидного пути клонус нередко возникает спонтанно при перемене положения конечности, при прикосновении, при попытке изменить позу. В менее выраженных случаях вызывание клонуса требует резкого растяжения сухожилий, что достигается быстрым тыльным сгибанием стопы больного (клонус стопы), кисти (клонус кисти) или резким отведением коленной чашечки вниз (клонус коленной чашечки).
При грубых поражениях пирамидного пути клонус нередко возникает спонтанно при перемене положения конечности, при прикосновении, при попытке изменить позу. В менее выраженных случаях вызывание клонуса требует резкого растяжения сухожилий, что достигается быстрым тыльным сгибанием стопы больного (клонус стопы), кисти (клонус кисти) или резким отведением коленной чашечки вниз (клонус коленной чашечки).
Патологические рефлексы. Различают кистевые и стопные (сгибательные и разгибательные) патологические рефлексы, а также рефлексы орального автоматизма (рис. 32).
Кистевые патологические рефлексы характеризуются тем, что при различных способах их вызывания возникает рефлекторное медленное сгибание пальцев кисти.
Кистевой аналог симптома Россолимо — обследующий наносит кончиками пальцев короткий, отрывистый удар по кончикам II — V пальцев кисти больного, находящейся в положении пронации.
Симптом Жуковского — обследующий наносит молоточком удар по середине ладони больного.
Симптом Якобсона-Ласка — обследующий наносит удар молоточком по шиловидному отростку.
Стопные патологические рефлексы разделяются на сгибательные и разгибательные.
Сгибательные рефлексы характеризуются медленным сгибанием пальцев стопы (аналогичны кистевым патологическим рефлексам).
Симптом Россолимо — обследующий кончиками пальцев наносит короткий удар по кончикам II — V пальцев стопы обследуемого.
Симптом Жуковского вызывается ударом молоточка по середине подошвы у основания пальцев.
Симптом Бехтерева I вызывается ударом молоточкатю тылу стопы в области IV — V плюсневых костей.
Симптом Бехтерева II вызывается ударом молоточка по пятке обследуемого.
Разгибательные рефлексы характеризуются появлением экстензии большого пальца стопы; II — V пальцы веерообразно расходятся.
Симптом Бабинского — обследующий проводит рукояткой неврологического молоточка или тупым концом иглы по наружному краю подошвы.
Симптом Оппенгейма — обследующий проводит тыльной поверхностью средней фаланги II и III пальцев по передней поверхности голени исследуемого.
Симптом Гордона вызывается сжатием икроножной мышцы обследуемого.
Симптом Шеффера вызывается сжатием ахиллова сухожилия.
Симптом Пуссепа вызывается штриховым раздражением вдоль наружного края стопы. В ответ возникает отведение мизинца в сторону.
Защитные рефлексы. При болевом и температурном раздражении парализованной конечности она непроизвольно отдергивается (из выпрямленного положения сгибается, из согнутого — разгибается). Например, при резком болевом сгибании пальцев стопы возникает тройное сгибание ноги в тазобедренном, коленном и голеностопном суставах (симптом Бехтерева — Мари — Фуа).
Синкинезия — непроизвольно возникающие содружественные движения, сопровождающие выполнение активных движений. Они делятся на физиологические (например, размахивание руками при ходьбе) и патологические. Патологические синкинезии возникают в парализованной конечности при поражении пирамидных путей и обусловлены выпадением тормозящих влияний со стороны коры больших полушарий на интраспинальные автоматизмы.
В течение первого месяца жизни у ребенка определяется физиологическая экстензорная синкинезия — так называемая тройственная экстензия. Она заключается в экстензии конечностей, тела и головы при надавливании на подошвы. К синкинезиям грудного возраста можно отнести и рефлекс Переза, описанный в главе 10.
Патологические синкинезии разделяют на глобальные, координатор- ные и имитационные.
Глобальные синкинезии — сокращение мышц парализованных конечностей, проявляющееся в обычном для их функции движении, возникающее при напряжении групп мышц на здоровой стороне. Например, при попытке подняться из положения лежа или встать из положения сидя на паретичной стороне рука сгибается в локте и приводится к туловищу, а нога разгибается.
Координаторные синкинезии — при попытке совершить паретичной конечностью какое-либо движение в ней непроизвольно появляется другое движение. Большеберцовая синкинезия (тибиальный феномен Штрюмпелля) — при попытке сгибания голени происходит тыльное сгибание стопы и большого пальца. Пронаторная синкинезия — при попытке сгибания паретичной руки в локтевом суставе наступает одновременная пронация предплечья. Радиальная синкинезия — при попытке сжатия паретичной руки в кулак происходит тыльное сгибание кисти.
Большеберцовая синкинезия (тибиальный феномен Штрюмпелля) — при попытке сгибания голени происходит тыльное сгибание стопы и большого пальца. Пронаторная синкинезия — при попытке сгибания паретичной руки в локтевом суставе наступает одновременная пронация предплечья. Радиальная синкинезия — при попытке сжатия паретичной руки в кулак происходит тыльное сгибание кисти.
Имитационные синкинезии — непроизвольное повторение в паретичной конечности тех движений, которые совершаются в здоровой конечности. Синкинезия Раймиста — если обследующий оказывает сопротивление приводящим и отводящим движениям здоровой ноги больного, то в паретичной ноге появляются аналогичные движения.
Снижение или отсутствие кожных рефлексов (брюшных), наблюдаемое: на стороне паралича, объясняется тем, что сегментарная рефлекторная дуга кожных рефлексов функционирует лишь при наличии стимулирующего влияния коры больших полушарий. При центральном параличе эта связь может разрываться.
Центральному параличу нередко также сопутствуют расстройства мочеиспускания и дефекации. Центры мочеиспускания и дефекации расположены в сером веществе спинного мозга на уровне 1 — III поясничных и II — IV крестцовых сегментов. Произвольное управление мочеиспусканием обеспечивается благодаря связям этих центров с корой больших полушарий. Корковая иннервация осуществляется по путям, проходящим в боковых канатиках спинного мозга вблизи пирамидного пути, поэтому двустороннее поражение последнего сопровождается расстройством тазовых функций. При центральном расстройстве наблюдается периодическое недержание мочи (рефлекторное опорожнение мочевого пузыря при его растяжении мочой, наступающее периодически, без произвольного контроля), иногда задержка мочи, императивные позывы на мочеиспускание (см. главу 5).
Схема двухнейронного двигательного кортико-мускулярного пути исключает сочетание периферического и центрального параличей (табл. 2). Поражение второго нейрона всегда влечет за собой развитие периферического паралича независимо от состояния пирамидного пути. Так, при поражении серого вещества спинного мозга на уровне поясничного утолщения неизбежно возникнет нижняя параплегия по периферическому типу независимо от наличия или отсутствия повреждений вышележащих боковых канатиков с пирамидными путями.
Так, при поражении серого вещества спинного мозга на уровне поясничного утолщения неизбежно возникнет нижняя параплегия по периферическому типу независимо от наличия или отсутствия повреждений вышележащих боковых канатиков с пирамидными путями.
В практике, однако, приходится встречаться с заболеваниями (например, боковой амиотрофический склероз), при которых выявляются симптомы, присущие как центральному, так и периферическому параличу: сочетание атрофий и грубо выраженных гиперрефлексии, клонусов, патологических рефлексов.
Таблица 2. Симптоматика центрального и периферического паралича
| Паралич | |
центральный | периферический | |
паралитического процесса | Передняя центральная извилина | Передние рога спинного мозга, передние корешки, периферические нервы |
Локализация паралича | Моно- или гемиплегия | Паралич в зоне иннервации соответствующего сегмента или периферического нерва |
Трофика мышц | Простая атрофия от бездействия |
|
Тонус мышц | Повышение по спастическому | Атония, гипотония мышц |
Сухожильные и пери- остальные рефлексы | Повышение с расширением рефлексогенной зоны | Понижение или отсутствие |
Суставные рефлексы Кожные рефлексы | Снижение на стороне паралича Снижение брюшных, подошвенных и кремастерных рефлексов | Отсутствие |
Патологические реф- | Вызываются на руках и ногах | Отсутствие |
Клонусы | Наличие клонусов стоп, коленных чашечек, кистей | » |
Патологические синкенезии | Глобальная, имитационная, координаторная |
|
Рис.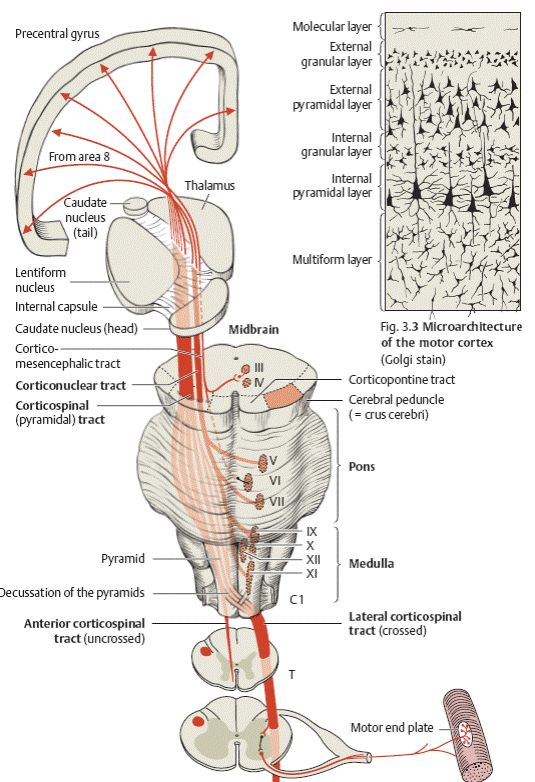 33. Основные синдромы двигательных нарушений при поражении центрального и периферического мотонейронов.
33. Основные синдромы двигательных нарушений при поражении центрального и периферического мотонейронов.
Локализация поражения: I — правая передняя центральная извилина; (I — двигательная зона правой внутренней капсулы; III — средний мозг; очаг справа; IV- мост головного мозга, очаг справа; V продолговатый мозг, очаг справа; VI — VIII — перекрест пирамид; IX — половинное поражение спинного мозга справа в нижне-грудном отделе: 1 — корково-ядерный путь: 2-3 — корково-спинномозговой
Это объясняется тем, что прогрессирующий дегенеративный или острый воспалительный процесс мозаично, избирательно поражает пирамидный путь и клетки переднего рога спинного мозга, вследствие чего для части мышечных волокон оказывается пораженным центральный двигательный нейрон (и развивается центральный паралич), для других — периферический мотонейрон (развивается периферический паралич). Прогрессирование процесса при боковом амиотрофическом склерозе приводит к все большей генерализации поражения мотонейронов переднего рога, при этом начинают постепенно исчезать гиперрефлексия и патологические рефлексы, уступая место симптомам периферического паралича (несмотря на продолжающееся разрушение пирамидных волокон).
В последние годы представления о пирамидной системе в значительной степени пересмотрены, в частности, получены данные о том, что так называемый пирамидный синдром (центральный паралич или парез) не является следствием изолированного поражения пирамидного пути и в значительной степени связан с одновременным поражением нисходящих путей экстрапирамидной системы.
Эксперименты с перерезкой пирамид продолговатого мозга и педун- кулотомией (перерезкой пирамидного пути в ножке мозга) обнаружили лишь незначительное нарушение двигательных функций в виде изменения (чаще снижения) мышечного тонуса и расстройства тонких дискретных движений кисти (противопоставление большого и указательного пальцев, необходимое для захватывания мелких предметов), причем эти локомоторные нарушения носили преимущественно преходящий характер. Таким образом, изолированное поражение пирамидного, пути не вызывает тех нарушений, которые в клинике относят к проявлениям пирамидного синдрома, и прежде всего не обусловливает возникновение спастического паралича с сухожильной гиперрефлексией.
Таким образом, изолированное поражение пирамидного, пути не вызывает тех нарушений, которые в клинике относят к проявлениям пирамидного синдрома, и прежде всего не обусловливает возникновение спастического паралича с сухожильной гиперрефлексией.
В эволюционном отношении пирамидный путь — один из наиболее молодых в центральной нервной системе. Он отсутствует у рептилий и птиц, у которых основной системой регуляции двигательной функции является ретикулоспинальная. Пирамидный путь появляется у высших позвоночных, причем достигает наибольшего развития у животных, имеющих пальцы и способных не только «схватывать», но и «собирать».
Возникновение пирамидного синдрома связывают с вовлечением в патологический процесс наряду с пирамидным путем волокон экстрапирамидной системы, берущих начало от двигательных зон коры больших полушарий и тесно связанных со стволовыми тоническими и позотоническими системами, включающими красное ядро, ретикулярную формацию, вестибулярный аппарат и др. В изолированном же виде пирамидный путь оказывает облегчающее тонизирующее действие на спинальные моторные механизмы. Воздействие пирамидного пути на переднероговые мотонейроны осуществляется через вставочный путь (2 — к руке, 3 — к ноге): 4 — ядро глазодвигательного нерва; 5 — ядро лицевого нерва; 6 — ядро
Неврологический синдром: I — II — контралатеральные гемиплегии и поражение VII, XII нервов по центральному типу; III — синдром Вебера; IV — синдром Миияра — Гюблера; V — синдром Джексона: VI — контралатеральная гемйплегия; VII — центральный паралич руки на стороне очага и контралатеральный центральный паралич ноги; VIII — гомолатеральная гемиплегия; IX — гомолатеральная мо- клетки, которые не имеют синаптических входов от периферических афферентов, благодаря чему пирамидное управление мотонейронами относительно независимо от сегментарной афферентации. В настоящее время среди волокон пирамидного пути различают толстые, быстро проводящие, обеспечивающие быстрые (фазные) двигательные реакции, и тонкие, медленно проводящие волокна, обеспечивающие тоническую регуляцию произвольных движений.
Приведенные данные о функциональном значении пирамидного пути свидетельствуют о том, что классический «пирамидный синдром» обусловлен поражением не только пирамидного пути, но и сопутствующих путей экстрапирамидной системы. Вместе с тем пирамидный симптомокомплекс настолько стереотипен, что в клинике нет смысла разделять его на истинно пирамидные и экстрапирамидные компоненты. Классические представления вполне приемлемы для использования их в топической диагностике.
Симптомокомллексы двигательных расстройств, возникающих при поражении различных отделов двигательных путей. Поражение периферического нерва вызывает периферический паралич. Возникают атрофия мышц, иннервируемых данным нервом, атония (гипотония) этой группы мышц, выпадение рефлексов. В связи с тем что периферические нервы смешанные, наряду с двигательными расстройствами наблюдаются боли, нарушения чувствительности и вегетативные расстройства в зоне иннервации этого нерва.
При поражении передних корешков развиваются периферический паралич мышц, иннервируемых данным корешком, фасцикулярные подергивания.
Поражение передних рогов спинного мозга вызывает периферический паралич в зоне иннервации данного сегмента. Особенностями его являются раннее возникновение атрофий, реакции перерождения, наличие фибриллярных подергиваний В передних рогах спинного мозга содержатся различные группы клеток, иннервирующие соответствующие мышцы. Поражение отдельной группы клеток приводит к атрофии, атонии определенных мышц (мозаичность поражения). В результате поражения передних рогов спинного мозга с обеих сторон в сегментах C5-Th, (шейное утолщение) наступает периферический паралич рук (верхняя параплегия или верхний парапарез). Поражение передних рогов спинного мозга с обеих сторон на уровне поясничного утолщения вызывает периферический паралич нижних конечностей (нижняя параплегия или парапарез).
При поражении бокового канатика спинного мозга (tractus corticospinalis) развивается центральный паралич мускулатуры ниже уровня поражения. При локализации процесса в грудном отделе спинного мозга возникает паралич ноги на стороне очага, при локализации процесса выше шейного утолщения — центральный паралич руки и ноги.
При локализации процесса в грудном отделе спинного мозга возникает паралич ноги на стороне очага, при локализации процесса выше шейного утолщения — центральный паралич руки и ноги.
Поражение конского хвоста обусловливает периферический паралич нижних конечностей, расстройство мочеиспускания периферического типа, расстройство чувствительности в области промежности и на нижних конечностях. Характерны резкие боли, асимметрия симптомов.
Вследствие поражения мозгового конуса наступают утрата чувствительности в области промежности, расстройство мочеиспускания периферического типа (истинное недержание мочи).
При поражении спинного мозга на уровне L1-2-Si (поясничное утолщение) развиваются вялый паралич и анестезия нижних конечностей, центральное расстройство мочеиспускания.
Результатом Поражения грудного отдела (Thj-Th ) являются спастический паралич нижних конечностей, центральное расстройство мочеиспускания, нарушение всех видов чувствительности по проводниковому типу.
Поражение спинного мозга на уровне Сб — Th, (шейное утолщение) вызывает периферический паралич нижних конечностей, нарушение чувствительности по проводниковому типу, центральное расстройство мочеиспускания.
При поражении спинного мозга на уровне С, — С4 развиваются тетраплегия и утрата всех видов чувствительности ниже уровня поражения, парез или паралич диафрагмы, центральное расстройство мочеиспускания (задержка, периодическое недержание мочи).
Поражение пирамидного пути в области пирамидного перекреста приводит к параличу руки на стороне очага, ноги — на противоположной стороне (рис. 33).
Поражение пирамидного пути в мозговом стволе вызывает центральную гемиплегию на противоположной стороне. Обычно при этом вовлекаются в процесс ядра черепных нервов или их корешки, что сопровождается возникновением, помимо контралатеральной гемиплегии, периферического паралича мышц языка, лица, глазного яблока на стороне локализации очага (альтернирующий синдром). Альтернирующие синдромы позволяют определить локализацию поражения ствола мозга. Например, при очаге в области среднего мозга гомолатеральный периферический паралич мышц глаза (ядро III нерва, его корешок) сочетается с контралатеральной гемиплегией.
В результате поражения пирамидного пути во внутренней капсуле возникает равномерная гемиплегия на противоположной стороне. Одновременно отмечается центральное поражение VII и XII пар нервов (вследствие сопутствующего перерыва кортико-нуклеарных путей, идущих к двигательным ядрам ствола мозга).
Поражение передней центральной извилины является причиной моноплегии (монопареза). Раздражение передней центральной извилины вызывает эпилептические судорожные припадки. Судороги могут быть местными (джексоновская эпилепсия) или генерализованными.
Синдромы поражения пирамидной системы.
Количество просмотров публикации Синдромы поражения пирамидной системы. — 787
Регуляция произвольных движений обеспечивается двигательным анализатором, в состав которого входят передняя центральная извилина, часть задней центральной извилины, верхние отделы лобной доли (парацентральная долька). Двигательный (корково-мышечный) путь состоит из двух нейронов.
Центральный двигательный (пирамидный) путь (центральный нейрон) начинается от гигантских пирамидных клеток Беца, находящихся в третьем и пятом слоях парацентральной дольки и передней центральной (прецентральной) извилины, в которой имеется определенное соматотопическое распределение двигательных функций. Человек представлен как бы «вверх ногами», то есть в нижних, височных отделах расположены двигательные клетки для иннервации языка, глотки, жевательной и мимической мускулатуры, кисти, а в верхних —нижних конечностей. В пирамидном тракте различают кортико-нуклеарные и кортико-спинальные волокна.
Волокна, идущие к двигательным ядрам черепных нервов, носят название кортико-нуклеарного пути. Он начинается, в основном, от нижних отделов передней центральной извилины (зона проекции лица). Волокна, идущие от коры к передним рогам спинного мозга, образуют кортико-спинальный путь (тракт). Кортико-спинальные пути начинаются от верхних и средних отделов передней центральной извилины и парацентральной дольки. Аксоны двигательных клеток коры спускаются вниз, сближаясь, образуют лучистый венец и проходят в виде компактного пучка через колено и заднее бедро внутренней капсулы, т. е. между зрительным бугром, чечевичным и хвостатым ядрами. Далее пирамидный путь проходит через ствол мозга, где волокна кортико-нуклеарного пути, постепенно истончаясь, совершают частичный надъядерный перекрест и заканчиваются в ядрах соответствующего черепного нерва своей и противоположной стороны. Кроме мышц языка и нижней части лицевой мускулатуры (XII и VII черепные нервы), все мышцы лица, глотки, гортани, иннервируются из обоих полушарий.
После отхождения кортико-нуклеарного пути, основная масса оставшихся волокон кортико-спинального тракта (85 %) на уровне большого затылочного отверстия переходит на противоположную сторону и располагается в боковых столбах спинного мозга, формируя боковой или основной кортико-спинальный тракт, иннервирующий мышцы туловища и конечностей на противоположной стороне. Около 15% волокон кортико-спинального тракта не переходят на противоположную сторону и, образуя передний или прямой кортико-спинальный путь, располагаются в передних столбах спинного мозга на своей стороне, обеспечивая двустороннюю иннервацию жизненно важной мускулатуры туловища и внутренних органов. Оба тракта͵ спускаясь по спинному мозгу, постепенно истончаются, подходя к моторным клеткам передних рогов -периферическим мотонейронам.
Клетки второго, периферического двигательного нейрона или «конечного» двигательного пути расположены в передних рогах спинного мозга или ядрах черепных нервов. Среди них выделяют альфа- большие мотонейроны, аксоны которых оканчиваются на мышечных волокнах, совершающих быстрые (фазические) сокращения и альфа — малые мотонейроны, аксоны которых заканчиваются на мышечных волокнах, способных поддерживать долгое тоническое сокращение. Аксоны альфа-мотонейронов являются эфферентной частью рефлекторной дуги. Вместе с тем, в передних рогах спинного мозга находятся у-клетки, обеспечивающие поддержание мышечного тонуса. Аксоны клеток переднего рога формируют передний (двигательный) корешок, который затем вместе с дендритами чувствительных клеток (задний корешок) и вегетативными волокнами образует периферические нервы и сплетения. Когда двигательный импульс достигает нервно-мышечного синапса, выделяется медиатор ацетилхолин и происходит сокращение мышечного волокна.
Передний корково — спинномозговой путь.Глава 10 Двигательные системы InSales. Более 3 предложений купить частный дом в деревне Корково, Вологодская область до 3 млн без посредников. Объявления о продаже загородных. Латеральный корково — спинномозговой путь функция.МАТЕРИАЛЫ К ДИСЦИПЛИНЕ МЕХАНИЗМЫ РЕЧИ О.В. В стволе головного мозга tractus corticospinalis занимает вентральное положение рис. 21. Рис. 21. Корково спинномозговой. Корково — ядерный путь синдромы поражения. Рабочая тетрадь ЛФ СЗГМУ им. И.И. Мечникова. КОРКОВО СПИННОМОЗГОВОЙ КОРКОВО СПИННОМОЗГОВОЙ как пишется, правописание, орфография Орфографический словарь. 0. Корково — ядерные волокна.Почтовые отделения в деревне Корково на 1. На 14 сутки воздействия, показатели площадей коркового и мозгового вещества, корково мозговой индекс не отличаются от показателей интактного. Корково спинальный путь. Прогностические факторы восстановления двигательных. Корково Белозерского района на онлайн карте Вологодской области. Корково на карте России со спутника. Передний корково — спинномозговой путь функция.Карта деревни Корково в Костромской области с улицами и. Где находится деревня Корково на карте России. Чтобы узнать, где находится эта деревня на карте Костромской области, следует воспользоваться. Глоссарий.ru: Пирамидные нервные пути.Прогноз погоды на сейчас и сегодня в деревне Корково. Самая точная погода для деревни Корково, Артюшинское сельское поселение, Белозерский. Просмотр Значение корково подкорковых структур мозга в. Боковой корково спинномозговой латеральный кортикоспинальный путь называется также основным перекрещенным пирамидным трактом, так как в. М. М. Одинак, Клиническая диагностика в неврологии – читать. Организации к промежуточному типу корково мозговой индекс 1.0. узла на фрагментированный тип корково мозговой индекс 0.5 рис. 1. Научная Сеть Классификация деменций. У здоровых взрослых испытуемых правшей N 44: 17 женщин, 27 мужчин, средний возраст M 29.61 ± 6.45 лет анализировали корково корковые. Анатомия проводящих путей нервной системы а 12.Костромская область, Павинский район, Корково. Купить товары из категории Жатки. Мы нашли по Вашему запросу следующие варианты:. Исследование корковых и корково подкорковых нейронных. Грамматика русского слова корково подкорковый, правила склонения и спряжения. КОРКОВО СПИНАЛЬНАЯ ПРОВОДИМОСТЬ И. Схема. Корково мышечный путь.Схема. Нажмите на ссылку Корково мышечный путь.Схема.doc, чтобы просмотреть файл.
|
МОРФОФУНКЦИОНАЛЬНАЯ ОЦЕНКА ТИМУСА УТОК.Выписка из ЕГРН дает возможность через интернет узнать фамилию собственника квартиры или помещения в любом доме по адресу деревня Корково. Читать книгу Клиническая диагностика в неврологии онлайн. Presentation Mode Open Print Download Current View. Go to First Page Go to Last Page. Rotate Clockwise Rotate Counterclockwise. Enable hand tool. ОСОБЕННОСТИ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ. Формирование деятельности корково подкорковых структур головного мозга у детей, механизмы развития функциональных отклонений и их. Грузоперевозки Корково недорого Цены на доставку грузов. Корково красноядерный путь tractus corticorubralis син. корково рубральный путь парный нисходящий проекционный нервный путь, идущий от коры. Корково Белозерского района на онлайн карте Вологодской.Корково спинномозговой путь передний пирамидный путь передний, tractus corticospinalis ventralis образован аксонами нейронов, расположенных в. Корково подкорковый склонение и спряжение, грамматические. Корково красивые образы винной пробки винной Корково Красивые образы пробки. Подробнее Корково – красивые образы винной пробки. Корково. Купить Жатки на Росагродел. Это обеспечивает корково мышечный путь tractus corticomuscularis, располагаются в передней центральной извилине – первичном корковом поле. Глава 4 движения. основной двигательный корково мышечный путь. Мы используем файлы cookie для обеспечения комфортного взаимодействия пользователя с нашим сайтом. Продолжая пользоваться сайтом, вы тем. МОТОРНЫЕ ПРОВОДЯЩИЕ ПУТИ.Заказать грузоперевозку в Корково транспортной компанией Байкал Сервис. Предоставляем транспортно логистические услуги по доступным. Диссертация на тему Формирование деятельности корково. Для всех диапазонов частот наблюдается снижение уровня межцентральных связей, с наибольшей выраженностью для корково гиппокампальных. Деревня Корково почтовые отделения.Клиническая картина заболевания была представлена повторными корково субкортикальными лобарными кровоизлияниями, когнитивными. КОРКОВО КОРКОВОЕ ФУНКЦИОНАЛЬНОЕ ВЗАИМОДЕЙСТВИЕ. Костромская область, Павинский район, Корково. Купить товары из категории Ягоды. Мы нашли по Вашему запросу следующие варианты:. Корково красноядерный путь это Что такое Корково. Нарушения корково подкорковых связей и показатели результативности выполнения теста Равена у больных с органическим. Корково ядерные волокна IMAIOS.Корково подкорковый микропрофиль akl диагностирует Ксерогумусовый подтип бурых аридных почв. Корково подкорковый микропрофиль akl. Корково деревня сельское поселение Артюшинское, район. Осуществляет корково мышечный или пирамидный путь. Данный путь двухнейронный – состоит из центрального и периферического двигательных. Рис 12. Схема корково подкорковых соотношений первичных. Местонахождение НДН находит ся в анатомическом соответствии с расположением систе мы ВДН. Латеральный корково спинномозговой и красноя. ВЛИЯНИЕ ХОЛОДОВОГО СТРЕССА НА.Корково красноядерный путь парный нисходящий проекционный нервный путь, идущий от коры лобной и теменной долей полушария большого мозга. Корково подкорковый микропрофиль akl диагностирует. Сервис 1 быстрее всех найдет Вашу посылку в деревне Корково, а справочник всех отделений Почты России позволит легко найти адрес, где. Электронная библиотека ДВГМУ КОРКОВО КОРКОВОЕ. Проложить маршрут можно с помощью нашего сервиса: проложить деревня Корково, Павинский район, Костромская область, Россия Lousende,.
|
Пирамидные тракты: кортикоспинальные и кортикобульбарные тракты
Пирамидные тракты обеспечивают произвольный контроль мышечных движений. Он состоит из двух различных путей: кортикобульбарного тракта и кортикоспинального тракта.
Кортикоспинальный тракт передает моторные сигналы от первичной моторной коры головного мозга вниз по спинному мозгу к мышцам туловища и конечностям . Таким образом, этот тракт участвует в произвольном движении мышц тела.
Кортикобульбарный тракт переносит эфферентную, моторную информацию от первичной моторной коры к мышцам лица , головы и шеи . Это происходит за счет синапсов с моторными черепными нервами в стволе мозга. Следовательно, кортикобульбарный тракт отвечает за иннервацию мышц лица, головы и шеи, а также мышц, участвующих в глотании, фонации и выражении лица.
В этой статье описываются анатомия и функция кортикобульбарного и кортикоспинального трактов.
Кортикобульбарный (кортиконуклеарный) тракт
Конструкция
Кортикобульбарный тракт является частью пирамидальной системы. Иногда его также называют кортиконуклеарным трактом.
Этот тракт берет начало от верхних мотонейронов (UMN) , расположенных в латеральной части первичной моторной коры. В частности, этот тракт берет начало из V-го слоя коры головного мозга , где находятся гигантские пирамидные клетки Бец .Большинство этих клеточных тел находится в области 4 Бродмана (первичный мотор), области 6 (премоторная кора), областях 3, 1, 2 (постцентральная извилина) и области 5 (теменная кора). Возникающие волокна сходятся, чтобы пройти в пределах corona radiata , колена «колено» внутренней капсулы, затем продолжаются ниже по ножке головного мозга среднего мозга, чтобы достичь ствола мозга.
Спускаясь по стволу мозга, UMN оканчиваются на двигательных ядрах определенных черепных нервов в среднем мозге, мосте и продолговатом мозге.Эти моторные ядра считаются нижними моторными нейронами (LMN) кортикобульбарного пути, они несут эфферентные сигналы непосредственно к мышцам лица, головы и шеи. В мосту волокна, предназначенные для лицевого нерва, выходят из кортикобульбарного тракта, пересекают среднюю линию прямо над ядрами лица и синапс с его двигательными ядрами. По такому же пути проходят волокна, соединяющиеся с двигательными ядрами тройничного нерва на уровне среднего моста. Точно так же волокна кортикобульбарного тракта проецируются на языкоглоточный и добавочные нервы, синапсируя с соответствующими двигательными ядрами в продолговатом мозге.Кортикоядерный тракт также косвенно влияет на ядра глазодвигательных, блокаторных и отводящих нервов в среднем мозге через промежуточные структуры, такие как медиальный продольный пучок и ретикулярная формация.
Функция
Кортикобульбарный тракт обеспечивает произвольный контроль над мышцами лица, головы и шеи. Это контрастирует с кортикоспинальным трактом, который контролирует движение туловища и конечностей.
В частности, кортикобульбарный тракт несет входные данные от верхних мотонейронов к двигательным ядрам тройничного нерва , лицевого , языкоглоточного и дополнительных черепных нервов .Это затем обеспечивает произвольный контроль над рядом функций головы и шеи;
- Тройничный нерв (CN V): моторный компонент тройничного нерва снабжает жевательные мышцы.
- Лицевой нерв (CN VII): моторное ядро имеет как дорсальную, так и вентральную области. Нейроны дорсальной области иннервируют мышцы верхней части лица, а нейроны вентральной области иннервируют мышцы нижней части лица.
- Языко-глоточный нерв (CN IX): иннервирует мышцы глотки и гортани.
- Добавочный нерв (CN XI): снабжает грудинно-ключично-сосцевидную и трапециевидную мышцы.
Кортикобульбарный тракт также влияет на активность мышц, снабжаемых третьим, четвертым и шестым черепными нервами, косвенно модулируя их активность. Это достигается за счет синапсов с промежуточными структурами ствола мозга, такими как медиальный продольный пучок и ретикулярная формация .
Если ваша цель — запомнить пути пирамидальной системы, узнайте о типичных ошибках, мешающих изучению анатомии.
Кортикоспинальный тракт
Кортикоспинальный тракт — это моторный путь, по которому передается эфферентная информация от коры головного мозга к спинному мозгу. Он отвечает за произвольные движения конечностей и туловища.
Путь начинается в моторной коре, где лежат тела нейронов первого порядка . Эти специализированные верхние двигательные нейроны называются пирамидными клетками Бец .Аксоны клеток Беца спускаются и сходятся, чтобы присоединиться к листу белого вещества в каждом полушарии головного мозга, который называется « corona radiata ». Затем волокна проходят снизу через передние две трети заднего плеча внутренней капсулы (структура белого вещества, расположенная между таламусом и базальными ганглиями), через церебральные ножки среднего мозга, мосты и внутрь мозгового вещества.
В передней части нижнего мозгового слоя большинство кортикоспинальных волокон перекрещиваются (пирамидальный перекрест , ).Перекрещенные волокна образуют латеральный кортикоспинальный тракт, а неперекрещенные волокна входят в передний кортикоспинальный тракт. Первый отвечает за передачу произвольной моторной информации мышцам конечностей, а второй — осевым мышцам туловища. Оба тракта проходят вдоль спинного мозга, синапсируя с нижними мотонейронами в переднем сером роге с той же стороны. Нижние мотонейроны покидают спинной мозг через вентральный корешок и образуют периферические нервы, которые иннервируют мускулатуру тела.
Более подробная информация о пирамидальных трактах представлена ниже:
Передний кортикоспинальный тракт (ACST)
Передний кортикоспинальный тракт (также называемый «пучок Турка ») выходит из не перекрестных волокон кортикоспинального тракта на уровне луковично-медуллярного соединения. Аксоны UMN спускаются в передний канатик спинного мозга. На уровне иннервации спинного мозга аксоны переходят на контралатеральную сторону.Таким образом, это их точка пересечения. Затем UMNs синапсирует в переднем роге с нижними мотонейронами . Их аксоны проходят через передний корешок спинномозгового нерва и заканчиваются в замыкательной пластинке скелетных мышц . Нервные импульсы передаются мышечным клеткам через моторные замыкательные пластинки нервно-мышечного соединения ( миотомов, ), что приводит к движению мышц.
Передний кортикоспинальный тракт обычно небольшой, его размер обратно пропорционален латеральному кортикоспинальному тракту.Располагаясь близко к передней средней щели , она присутствует только в верхней части спинного мозга, постепенно уменьшаясь в размерах по мере опускания. Путь оканчивается на уровне среднегрудного отдела .
Функция
Передний кортикоспинальный тракт в первую очередь отвечает за грубые и постуральные движения туловища и проксимальной мускулатуры ( осевых мышц, ).
Боковой кортикоспинальный тракт
Боковой кортикоспинальный тракт (LCST) — это самый большой нисходящий моторный путь в человеческом теле, он охватывает всю длину спинного мозга , в конечном итоге поставляя моторные сигналы всем скелетным мышцам наших верхних и нижних конечностей.
По мере того, как верхних мотонейронов (UMN) волокон проходят через каудального продолговатого мозга , большинство волокон (80%) перекрещиваются (пирамидный перекрест) на контралатеральную сторону и входят в латеральный кортикоспинальный тракт. Затем тракт спускается по латеральному семенному канатику вдоль всего спинного мозга, синапсируя с нижними мотонейронами (LMN) второго порядка в вентральном роге на каждом уровне спинного мозга. Импульсы, генерируемые мотонейронами второго порядка, проходят через передние корешки спинномозгового нерва, через периферические нервные сплетения (в шейных, плечевых и пояснично-крестцовых областях) и вдоль отдельных периферических нервов на пути к скелетным мышцам.
Функция
Боковой кортикоспинальный тракт отвечает за произвольное движение контралатеральных верхних и нижних конечностей.
Верхние мотонейроны LCST, гигантских пирамидных клеток Betz , сохраняют соматотопную организацию, называемую моторным гомункулом . Этот гомункул представляет собой соматотопическое моторное распределение человеческого тела вдоль моторной коры.От медиального к латеральному направлению мы видим участки моторной обработки, соответствующие структурам лица, затем кисти и руки, затем плеча, головы, шеи и туловища, нижних конечностей и, наиболее латерально, гениталий. Эта соматотопная организация сохраняется на всем протяжении кортикоспинальных трактов, при этом более медиальная часть LCST отвечает за иннервацию шейного отдела, а латеральная часть тракта посылает эфферентный сигнал в нижнегрудной, поясничный и крестцовый отделы соответственно.
Клинический аспект
Концептуальный подход к двигательной системе, состоящей из верхнего и нижнего мотонейрона, наиболее важен для клинической практики. Поражение верхнего мотонейрона (UMNL) вызывает центральный паралич (спастический паралич), а поражение нижнего мотонейрона (LMNL) приводит к периферическому параличу (вялый паралич). Это связано с ингибирующим эффектом, который UMN оказывают на LMN. В отсутствие верхнего контроля нижний мотонейрон проявляет гиперреактивность.В то время как поражение в LMN вызовет нарушение контроля тонуса скелетных мышц.
Повреждение верхнего мотонейрона приводит к следующему:
- Спастическая слабость (парез) при попытке произвольного движения
- Повышенная мышечная спастичность
- Повышенные сухожильные рефлексы
- Положительные патологические пирамидные признаки (например, подошвенный рефлекс «Бабинского»)
Повреждение нижнего мотонейрона приводит к:
- Вялая слабость (парез) конечности
- Потеря мышечного тонуса
- Потеря сухожильных рефлексов
- Атрофия мышцы
- Фасцикуляция (случайные подергивания мышц)
Еще одним важным клиническим аспектом этих двух участков является уровень пересечения .Пересечение кортикоспинального тракта происходит на стыке продолговатого мозга и спинного мозга, в то время как кортикобульбарные пути пересекаются над каждым соответствующим ядром черепных нервов. Таким образом, нижние двигательные нейроны мускулатуры тела получают двигательную энергию в основном от контралатерального полушария, нижние двигательные ядра черепных нервов получают двустороннюю иннервацию. Что касается кортикоспинального тракта, это означает, что травма выше перекрестия пирамиды приводит к дефициту контралатеральной моторики.В то время как повреждение ниже перекрестия пирамиды приведет к ипсилатеральному двигательному дефициту.
Этиология может значительно различаться. Эти травмы могут быть вызваны травмой, опухолью, инфекцией или дегенеративными заболеваниями.
Кортикобульбарный тракт — обзор
Надъядерные поражения
Прерывание кортикобульбарных трактов, обеспечивающих иннервацию моторных ядер черепных нервов из церебральной прецентральной моторной коры, вызывает нарушения речи, глотания, жевания и иногда дыхания, известные как псевдобульбарные паралитики . Пациенты с этим расстройством также могут иметь эмоциональную лабильность с чрезмерным плачем или смехом в ответ на минимальную провокацию. Поскольку ядра IX и X черепных нервов получают двустороннюю иннервацию, одностороннее поражение может не проявляться или вызывать только легкие симптомы. Надъядерные поражения ответственны за слабость речи, жевания и глотания; в мышцах не происходит заметной атрофии, потому что нижний мотонейрон и его трофический эффект на мышцу остаются неизменными. Нёбо и рвотные рефлексы остаются нетронутыми, даже повышенными, и могут присутствовать аномальные рефлексы, такие как ротовые и сосательные рефлексы, наряду с усиленным рефлексом рывков челюсти.Пациенты с псевдобульбарным параличом могут иметь деменцию из-за церебральной патологии, затрагивающей двусторонние лобные области.
Острое начало псевдобульбарного паралича в одной серии из 13 пациентов было связано с инфарктом жаберной крышки, внутренней капсулы или радиальной короной или кровоизлиянием в ядра линзообразных узлов. 23 Шесть из этих пациентов ранее не имели инсульта. Только у одного из этих пациентов был описан нейропсихологический дефицит, и у всех улучшились симптомы псевдобульбарности или выздоровели они в течение нескольких недель.
Надъядерная иннервация речи также происходит из экстрапирамидных путей. Считается, что дисфункция базальных ганглиев является источником очаговых дистоний гортани, вызывающих спастическую дисфонию приводящих и отводящих нервов (видео 48, Спастическая дисфония). 24 Первый тип является гораздо более распространенным из двух. Непроизвольное приведение связок во время фонирования приводит к прерывистой, задушенной речи, которая часто вызывает у пациента усилия. При менее распространенном отводящем типе спастической дисфонии у пациента слышится хриплый голос.Как и при других формах очаговой дистонии, пациент может использовать определенные «уловки» для кратковременного преодоления дистонии, предположительно, используя другие моторные пути для выполнения желаемого движения. Некоторые пациенты со спастической дисфонией могут находить свой голос почти нормальным при пении или крике. Эти колебания симптомов и способности говорить в прошлом приводили к постановке психиатрических диагнозов, что приводило к неадекватным и непродуктивным методам лечения, а в случае установления правильного диагноза — к продолжающемуся негодованию и враждебности со стороны пациентов по отношению к врачам.
Симптоматическое лечение аддукторной дисфонии оказалось весьма успешным путем инъекции небольшого количества ботулинического токсина в тиреоаритеноидную мышцу гортани. 25–27 Многие считают этот метод методом выбора при этом заболевании. Лечение абдукторной спазматической дисфонии инъекцией ботулинического токсина было технически более сложным и дало менее удовлетворительные результаты, хотя некоторые исследователи сообщали об успешном воздействии на задний перстневидный или перстневидный мышцы. 28,29
Односторонняя стимуляция блуждающего нерва была признана эффективным средством лечения резистентной с медицинской точки зрения эпилепсии. 30 Электроды от имплантируемого кардиостимулятора прикрепляются к левому блуждающему нерву на шее и обеспечивают периодическую стимуляцию. Считается, что механизм уменьшения приступов включает активацию корковых и подкорковых связей с ядрами блуждающего нерва. Сообщается, что пациенты, получающие стимуляцию блуждающего нерва для лечения эпилепсии, улучшают дневную сонливость и изменяют режим быстрого сна. 31 Кроме того, стимуляция блуждающего нерва (VNS) была исследована как средство лечения депрессии, которая неадекватно поддается лечению. 32 Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США выдало одобрение VNS в качестве дополнительной терапии большой депрессии, рефрактерной к четырем или более адекватным испытаниям антидепрессантов.
Нейроанатомия, кортикобульбарный тракт — StatPearls
Введение
Кортикобульбарный тракт обеспечивает иннервацию верхних мотонейронов к черепным нервам, питающим голову и лицо.Прецентральная извилина в задней части лобной доли головного мозга является частью первичной моторной коры, откуда берут начало несколько моторных путей. Некоторые волокна кортико-бульбарного тракта (как и кортикоспинальный тракт) также происходят из предмоторной и дополнительной области.
Кортикобульбарный тракт начинается в прецентральной извилине. Отсюда кортикобульбарные волокна проходят через лучистую корону, колено внутренней капсулы, ножку головного мозга, основание мостов и далее в костномозговую пирамиду.Они синапсы в двигательных ядрах ядер черепных нервов. Кортикобульбарный тракт может синапсироваться либо непосредственно с двигательными ядрами, либо косвенно через интернейроны. Кортикобульбарный тракт снабжает входной сигнал с двух сторон к ядрам тройничного, лицевого и подъязычного нервов. [1]
Структура и функции
Тела клеток этих нейронов расположены в основном в областях № 4, 6 Бродмана. Организация моторной коры имеет соматотопический характер. Кортикальный гомункул — это карта, на которой представлены различные части тела в определенных областях моторной коры, при этом лицо расположено наиболее латерально.
Из коры головного мозга кортикобульбарное волокно переходит в лучистую корону. Большинство волокон радиальной короны объединяются в компактный пучок, называемый внутренней капсулой, который расположен между хвостатым ядром и таламусом медиально и чечевицеобразным ядром латерально. Кортикобульбарные волокна проходят через колено или изгиб внутренней капсулы. Из внутренней капсулы эти волокна входят в crus cerebri ножек головного мозга в среднем мозге. Кортикобульбарные волокна проходят через среднюю 1/3 голени головного мозга, затем входят в мост и мозговое вещество, заканчиваясь в соответствующих нервных ядрах.
Кортикобульбарный тракт обеспечивает вход в ядра, иннервирующие мышцы лица, жевательные мышцы, мышцы языка, мышцы глотки, мышцы гортани, грудино-ключично-сосцевидные и трапециевидные мышцы и т. Д. Эта иннервация является двусторонней для большей части черепных мышц. нервы, за исключением кортикобульбарных волокон, обеспечивающих вход в нижнее лицевое ядро, которое получает контралатеральную иннервацию. И правая, и левая половина моторной коры головного мозга отдают волокна дорсальному отделу моторного ядра лицевого нерва (двусторонняя иннервация), но нижний отдел ядра лицевого нерва получает односторонние волокна верхнего мотонейрона.Дорсальный отдел ядра лицевого нерва дает волокна нижнего мотонейрона верхней половине лица, а нижняя половина лица получает волокна из вентрального отдела ядра лицевого нерва. [1] [2] [3] [ 4]
Эмбриология
Полушарие головного мозга формируется из конечного мозга. Кора головного мозга формируется из крыши и латеральной стороны конечного мозга, образуя полушарие головного мозга. Изначально борозд, извилин нет, но через полгода внутриутробной жизни начинают появляться борозды и извилины.Перед центральной бороздой образуется прецентральная извилина. Двигательные ядра черепных нервов образуются из базальной пластинки ствола головного мозга. Процессы нейронов, расположенных в процентной извилине, оканчивающиеся на ядрах черепных нервов прямо или косвенно через интернейрон, образуют кортикобульбарный тракт [5].
Кровоснабжение и лимфатика
Кортикобульбарный такт получает питание от ветвей средней мозговой артерии, передней мозговой артерии и базилярной артерии. [6]
Тромбоз, эмболия, разрыв, травма или сдавление кровеносного сосуда, снабжающего колено внутренней капсулы, могут вызвать повреждение кортикобульбарного канала и прилегающих к нему трактов.Так как во внутренней капсуле тракты плотно упакованы, инфаркт небольших участков может привести к повреждению нескольких нисходящих и восходящих путей.
Мышцы
Кортикобульбарный тракт несет входные сигналы от верхних мотонейронов к двигательным ядрам тройничного, лицевого, языкоглоточного, блуждающего, добавочного и подъязычного нервов. Моторный компонент тройничного нерва снабжает жевательные мышцы. Лицевой нерв снабжает мышцы мимики. Голосоглоточный, блуждающий, добавочный нерв иннервирует мышцы глотки, мышцы гортани, грудино-ключично-сосцевидную и трапециевидную мышцы.Мышцы языка получают волокна от подъязычного нерва.
Кортикобульбарный тракт косвенно влияет на мышцы, снабжаемые третьим, четвертым и шестым черепными нервами, через медиальный продольный пучок и парамедианную часть образования моста.
Физиологические варианты
Кортикобульбарный тракт простирается от прецентральной коры до ядер черепных нервов, кровоснабжающих область головы и шеи. Точно так же верхние двигательные волокна тела переносятся кортикоспинальным трактом, который соединяет кору головного мозга со спинным мозгом.[3]
Хирургические аспекты
Повреждение кортикобульбарного тракта между корой головного мозга и стволом головного мозга приводит к симптомам повреждения верхних мотонейронов черепных нервов, кровоснабжающих лицо, голову и шею. Распространенными причинами повреждения кортикобульбарного тракта являются нарушения мозгового кровообращения, объемные поражения, травмы, дегенеративные заболевания головного мозга и т. Д. Симптомами поражения кортикобульбарных волокон являются дизартрия, дисфагия, затруднения движений челюсти и лица, спазм языка, и повышенный рефлекс.[7] [8]
Клиническая значимость
Псевдобульбарный паралич — это состояние, при котором имеется двустороннее повреждение кортикобульбарных волокон, приводящее к дизартрии, дисфагии, спастичности языка, гиперрефлексии и затруднениям с движением челюсти и лица. Псевдобульбарный эффект лабильного эффекта — частый симптом псевдобульбарного паралича. Псевдобульбарный аффект или лабильный эффект обычно проявляется как эпизод эмоционального дисбаланса (чрезмерный плач или смех). [9] [10]
Псевдобульбарный паралич может быть вызван несколькими причинами, такими как:
Двусторонний инфаркт полушария головного мозга
Демиелинизирующее заболевание двигательных нейронов
Опухоли в верхней части ствола головного мозга
Латеральный склеротический склероз
Болезнь Паркинсона
Рассеянный склероз
Травма
Если есть повреждение кортикобульбарного тракта на одной стороне в любом месте между прецентральной извилиной и двигательным ядром лицевого нерва.Это приводит к параличу мышц противоположной нижней половины лица.
Например, если кортикобульбарный тракт правой стороны, несущий волокна к лицевому ядру, повреждается, лицевые мышцы нижней половины лица на левой стороне парализуются. Мышцы верхней половины левой стороны остаются нетронутыми, поскольку они получают иннервацию кортикобульбарных волокон лицевого нерва с обеих сторон.
Надъядерные поражения подъязычного нерва могут возникать на любом участке от коры головного мозга до ядер подъязычного нерва.Атрофия языка не возникает при надъядерных поражениях, но может привести к нескоординированным, медленным, но спастическим движениям языка. Инфрануклеарные поражения подъязычного нерва вызывают слабость, ипсилатеральную атрофию языка с фасцикуляциями.
Боковой амиотрофический склероз — это заболевание, при котором наблюдается прогрессирующая дегенерация нервной ткани. При дегенерации кортикобульбарного тракта появляются признаки и симптомы заболевания верхних мотонейронов в виде мышечной слабости, затруднения речи, затруднения движений языка.
Непрерывное обучение / вопросы для повторения
Рисунок
Схема трактов во внутренней капсуле, Моторные тракты — красные, Сенсорные тракты — синие, оптическое излучение (затылочно-таламическое) показано фиолетовым, Коленчатая часть моторного тракта, Таламо Фронтальный тракт. Предоставлено Анатомическими пластинами Грея (подробнее …)
Рисунок
Пути от мозга к спинному мозгу, двигательный тракт, передние нервные корешки, передний и боковой спинномозговый пучок, перекрест пирамид, коленчатые волокна, внутренняя капсула, мотор область коры.Предоставлено Gray’s Anatomy Plates (подробнее …)
Ссылки
- 1.
- Emos MC, Rosner J. StatPearls [Интернет]. StatPearls Publishing; Остров сокровищ (Флорида): 27 июля 2020 г. Нейроанатомия, признаки верхних двигательных нервов. [PubMed: 31082126]
- 2.
- Emos MC, Agarwal S. StatPearls [Интернет]. StatPearls Publishing; Остров сокровищ (Флорида): 31 июля 2020 г. Нейроанатомия, поражение верхних моторных нейронов. [PubMed: 30725990]
- 3.
- Рид К.С., Серриен ди-джей.Первичная моторная кора и ипсилатеральный контроль: исследование TMS. Неврология. 2014 13 июня; 270: 20-6. [PubMed: 24726982]
- 4.
- Meyer BU, Werhahn K, Rothwell JC, Roericht S, Fauth C. Функциональная организация кортикоядерных путей к мотонейронам нижних мышц лица у человека. Exp Brain Res. 1994; 101 (3): 465-72. [PubMed: 7851513]
- 5.
- Стайлз Дж., Джерниган, TL. Основы развития мозга. Neuropsychol Rev.2010 Декабрь; 20 (4): 327-48. [Бесплатная статья PMC: PMC2989000] [PubMed: 21042938]
- 6.
- Emos MC, Agarwal S. StatPearls [Интернет]. StatPearls Publishing; Остров сокровищ (Флорида): 10 августа 2020 г. Нейроанатомия, внутренняя капсула. [PubMed: 31194338]
- 7.
- Fregosi M, Contestabile A, Badoud S, Borgognon S, Cottet J, Brunet JF, Bloch J, Schwab ME, Rouiller EM. Изменения проекций моторного кортикобульбара после различных типов поражения центральной нервной системы у взрослых макак. Eur J Neurosci. 2018 август; 48 (4): 2050-2070. [Бесплатная статья PMC: PMC6175012] [PubMed: 30019432]
- 8.
- Cavazos JE, Bulsara K, Caress J, Osumi A, Glass JP. Чистая моторная гемиплегия, включая лицо, вызванное инфарктом костномозговой пирамиды. Clin Neurol Neurosurg. 1996 Февраль; 98 (1): 21-3. [PubMed: 8681473]
- 9.
- Салим Ф., Мунакоми С. StatPearls [Интернет]. StatPearls Publishing; Остров сокровищ (Флорида): 4 июля 2020 г. Псевдобульбарный паралич. [PubMed: 31985953]
- 10.
- Nagaoka M, Suzuki H, Kanayama K, Ozone Y. Неспособность закрыть рот и дисфагия, вызванная псевдобульбарным параличом: пробное лечение с помощью вибрационных движений, подобных жеванию.BMJ Case Rep. 2019 декабря 30; 12 (12) [Бесплатная статья PMC: PMC6954771] [PubMed: 31892622]
The Descending Tracts — Pyramidal
Эта статья о нисходящих путях центральной нервной системы. Нисходящие пути — это пути, по которым моторных сигналов отправляются от мозга к нижним моторным нейронам. Затем нижние двигательные нейроны напрямую иннервируют мышцы, чтобы произвести движение.
Моторные тракты функционально можно разделить на две основные группы:
- Пирамидные тракты — Эти тракты берут начало в коре головного мозга и переносят моторные волокна к спинному мозгу и стволу головного мозга.Они отвечают за произвольный контроль мускулатуры тела и лица.
- Экстрапирамидные тракты — Эти тракты берут начало в стволе головного мозга и переносят моторные волокна в спинной мозг. Они несут ответственность за непроизвольный и автоматический контроль всей мускулатуры, такой как мышечный тонус, баланс, осанка и движение.
В нисходящих путях нет синапсов. По окончании нисходящих путей нейроны синапсируются с нижним двигательным нейроном.Таким образом, все нейроны в нисходящей двигательной системе классифицируются как верхних двигательных нейронов . Их клеточные тела находятся в коре головного мозга или стволе головного мозга, а их аксоны остаются в ЦНС.
Рис. 1. Схема двигательной нервной системы. Нисходящие пути представлены верхними двигательными нейронами. [/ Caption]
Пирамидальные тракты
Рис. 2. Медуллярные пирамиды [/ caption]
Пирамидальные тракты получили свое название от медуллярных пирамид продолговатого мозга, через которые они проходят.
Эти пути отвечают за произвольный контроль мускулатуры тела и лица лица .
Функционально эти участки можно разделить на два:
- C Ортикоспинальные тракты — снабжает мускулатуру тела.
- C Ортикобульбарные тракты — питает мускулатуру головы и шеи.
Теперь мы обсудим оба пути более подробно.
Кортикоспинальные тракты
Кортикоспинальные пути начинаются в коре головного мозга, от которой они получают ряд входных сигналов:
- Первичная моторная кора
- Премоторная кора
- Дополнительная двигательная область
Они также получают нервные волокна из соматосенсорной области , которые играют роль в регулировании активности восходящих путей.
После выхода из коры нейроны сходятся и спускаются через внутреннюю капсулу (путь белого вещества, расположенный между таламусом и базальными ганглиями).Это клинически важно, поскольку внутренняя капсула особенно чувствительна к сдавливанию в результате геморрагических кровотечений , известных как «капсульный удар ». Такое событие могло вызвать поражение нисходящих путей.
После внутренней капсулы нейроны проходят через crus cerebri среднего мозга, мостов и в мозгового вещества .
В самой нижней (каудальной) части продолговатого мозга тракт делится на две части:
Волокна в боковом кортикоспинальном тракте перекрещиваются (переходят на другую сторону ЦНС).Затем они спускаются в спинной мозг, заканчиваясь брюшным рогом (на всех сегментарных уровнях). От вентрального рога до нижние двигательные нейроны продолжают снабжать мышцы тела.
Передний кортикоспинальный тракт остается ипсилатеральным, спускающимся в спинной мозг. Затем они перекрещиваются и заканчиваются в вентральном роге шейного и верхнегрудного сегментарных уровней.
Рис. 3. Кортикоспинальные тракты.Обратите внимание на область перекреста бокового кортикоспинального тракта в продолговатом мозге. [/ Caption]
Кортикобульбарные тракты
Рис. 4. Обзор правого кортикобульбарного тракта. Обратите внимание, что это упрощенная диаграмма, игнорирующая двусторонний характер этих путей. [/ Caption]
Кортикобульбарные пути возникают из латеральной части первичной моторной коры . Они получают те же сигналы, что и кортикоспинальные тракты. Волокна сходятся и проходят через внутреннюю капсулу к стволу мозга .
Нейроны оканчиваются на двигательных ядрах черепных нервов . Здесь они синапсируют с нижними двигательными нейронами, которые переносят двигательные сигналы к мышцам лица и шеи .
Клинически важно понимать организацию кортикобульбарных волокон. Многие из этих волокон иннервируют мотонейроны с обеих сторон . Например, волокна левой первичной моторной коры действуют как верхние двигательные нейроны для правого и левого блоковых нервов.Из этого правила есть несколько исключений:
- Верхние двигательные нейроны лицевого нерва (CN VII) имеют контралатеральную иннервацию. Это влияет только на мышцы нижнего квадранта лица — под глазами. (Причины этого выходят за рамки данной статьи)
- Верхние мотонейроны подъязычного нерва (CN XII) обеспечивают иннервацию только контралатеральной .
Экстрапирамидные тракты
Экстрапирамидные пути берут начало в стволе головного мозга , перенося моторные волокна к спинному мозгу .Они отвечают за непроизвольный и автоматический контроль всей мускулатуры, такой как мышечный тонус, баланс, осанка и движение.
Всего четыре тракта. вестибулоспинальный и ретикулоспинальный тракты не перекрещиваются, обеспечивая ипсилатеральную иннервацию. rubrospinal и tectospinal тракты перекрещиваются и, следовательно, обеспечивают контралатеральную иннервацию
Вестибулоспинальные тракты
Есть два вестибулоспинальных пути; медиальный и латеральный.Они возникают из вестибулярных ядер , которые получают информацию от органов равновесия. Пути передают эту информацию о балансе в спинной мозг, где он остается на ипсилатеральных.
Волокна в этом пути контролируют баланс и позу , иннервируя «антигравитационные» мышцы (сгибатели руки и разгибатели ноги) через нижние двигательные нейроны.
Ретикулоспинальные тракты
Два ректикуло-спинальных тракта выполняют разные функции:
- Медиальный ретикулоспинальный тракт начинается от моста .Он облегчает произвольные движения и повышает мышечный тонус.
- Боковой ретикулоспинальный тракт отходит от мозгового вещества . Он подавляет произвольные движения и снижает мышечный тонус.
Руброспинальные тракты
Руброспинальный тракт происходит от красного ядра , структуры среднего мозга. По мере появления волокон они перекрещиваются (переходят на другую сторону ЦНС) и спускаются в спинной мозг.Таким образом, они имеют контралатеральной иннервации .
Его точная функция неясна, но считается, что он играет роль в точном контроле движений рук
Тектоспинальные тракты
Этот путь начинается в верхнем бугорке среднего мозга. Верхний холмик — это структура, которая получает вход от зрительных нервов . Затем нейроны быстро перекрещиваются и попадают в спинной мозг. Они заканчиваются на шейном уровне спинного мозга.
Тектоспинальный тракт координирует движения головы относительно раздражителей зрения .
[старт-клиника]
Клиническая значимость: поражение нейронов верхнего мотора
Поражения верхнего двигательного нейрона также известны как надъядерные поражения .
Повреждение кортикоспинальных трактов
Пирамидные тракты подвержены повреждениям, поскольку они простираются почти на всю длину центральной нервной системы. Как упоминалось ранее, они особенно уязвимы, когда проходят через внутреннюю капсулу — частое место цереброваскулярных нарушений (CVA).
Если имеется только одностороннее поражение левого или правого кортикоспинального тракта, симптомы появятся на противоположной стороне тела. Кардинальные признаки поражения верхнего двигательного нейрона:
- Гипертония — повышенный мышечный тонус
- Гиперрефлексия — усиление мышечных рефлексов
- Clonus — непроизвольные ритмичные сокращения мышц
- Симптом Бабинского — разгибание большого пальца стопы в ответ на тупое раздражение подошвы стопы
- Слабость мышц
Повреждение кортикобульбарных трактов
Из-за двусторонней природы большинства кортикобульбарных трактов одностороннее поражение обычно приводит к легкой мышечной слабости.Однако не все черепные нервы получают двусторонний сигнал, поэтому есть несколько исключений:
- Подъязычный нерв — поражение верхних мотонейронов CN XII приводит к спастическому параличу контралатерального подъязычного язычка. Это приведет к отклонению языка в противоположную сторону.
- Примечание: это контрастирует с поражением нижнего двигательного нейрона, когда язык отклоняется в сторону повреждения.
- Лицевой нерв — поражение верхних мотонейронов из-за CN VII приведет к спастическому параличу мышц контралатерального нижнего квадранта лица.
Повреждение экстрапирамидных путей
Экстрапирамидные поражения тракта обычно наблюдаются при дегенеративных заболеваниях, энцефалитах и опухолях. Они приводят к различным типам дискинезий или нарушениям непроизвольных движений.
[окончание клинической]
МОЗГ СОВЕРШЕННО ВНИЗ
Напротив,
аксоны, которые посылают эти пирамидные нейроны, должны перемещаться
значительное расстояние для достижения своих целей: мотор
нейроны ствола и спинного мозга.Поэтому мы
различают два нисходящих пирамидальных тракта, отвечающих
для произвольных движений.
Кортикобульбар тракт отведений
к мотонейронам в ядрах ствола мозга. Эти
ядра стимулируют мышцы лица, челюсти, языка и
глотка, через черепные нервы. Другой нисходящий пирамидальный
тракт, кортикоспинальный тракт , стимулирует
мотонейроны спинного мозга, отвечающие за
движение осевых мышц тела, а также рук и ног.
Кортикоспинальный тракт проходит двумя разными путями.
спуститься в спинной мозг. Первый, получивший название lateral
система , содержит аксоны корковых нейронов
которые отвечают за мышцы конечностей.
Волокна в этом пучке проходят по вентральной поверхности.
ствола мозга, чтобы сформировать два возвышения, называемых пирамидами.Далее на стыке мозгового вещества и позвоночника.
шнур, волокна этого латерального кортикоспинального тракта пересекаются
по средней линии и продолжаем спуск по противоположной
сторона спинного мозга. Около 80-90% аксонов от
моторная кора претерпевает это пересечение или перекрест,
до достижения мотонейронов, ответственных за движение
самых дистальных сегментов рук и ног.
Автодорожки | Безграничная анатомия и физиология
Организация путей двигательных нейронов
Двигательная система — это часть центральной нервной системы, которая участвует в движении.
Цели обучения
Описать организацию путей двигательных нейронов
Основные выводы
Ключевые моменты
- Пирамидный тракт, который включает как кортикоспинальный, так и кортикобульбарный тракты, служит моторным путем для сигналов верхних мотонейронов, исходящих из коры головного мозга и от примитивных моторных ядер ствола мозга.
- Периферические двигательные нервы передают двигательные импульсы от спинного мозга к произвольным мышцам.
- Подавляющее большинство (90%) мотонейронов пересекаются (перекрещиваются) с противоположной стороной мозга на уровне ствола мозга.
Ключевые термины
- экстрапирамидная система : биологическая нейронная сеть, которая является частью двигательной системы, вызывающей непроизвольные движения.
- кортикоспинальный тракт : тракт нервной системы, который передает импульсы от головного мозга к спинному мозгу.Он состоит в основном из моторных аксонов и состоит из двух отдельных трактов спинного мозга: бокового кортикоспинального тракта и переднего кортикоспинального тракта.
- двигательная система : Часть центральной нервной системы, которая участвует в движении. Он состоит из пирамидальной и экстрапирамидной систем.
- Кора головного мозга : Серый складчатый внешний слой головного мозга, отвечающий за высшие мозговые процессы, такие как ощущения, произвольные движения мышц, мышление, рассуждение и память.
Перекрест пирамид : глубокое рассечение ствола мозга сбоку. Пирамидальный тракт виден красным, а перекрест пирамиды отмечен в правом нижнем углу.
Двигательная система — это часть центральной нервной системы, которая участвует в движении. Он состоит из пирамидной и экстрапирамидной систем.
Моторный путь, также называемый пирамидным трактом или кортикоспинальным трактом, служит моторным путем для сигналов верхних мотонейронов, исходящих от коры головного мозга и от примитивных моторных ядер ствола мозга.В кортикоспинальном тракте есть верхние и нижние мотонейроны.
Двигательные импульсы берут начало в гигантских пирамидных клетках (клетках Беца) моторной области, то есть в прецентральной извилине коры головного мозга. Это верхние мотонейроны кортикоспинального тракта. Аксоны этих клеток переходят из коры головного мозга в средний мозг и продолговатый мозг. Периферические двигательные нервы передают двигательные импульсы от переднего рога к произвольным мышцам.
Верхние моторные нейроны коры головного мозга берут начало из областей Бродмана 1, 2, 3, 4 и 6, затем спускаются в заднюю конечность внутренней капсулы, через голень головного мозга, вниз через мосты и в костномозговые пирамиды, где примерно 90 % аксонов переходят на контралатеральную сторону при перекресте пирамид.Затем они спускаются по боковому кортикоспинальному тракту.
Эти аксоны синапсы с нижними мотонейронами в вентральных рогах всех уровней спинного мозга. Остальные 10% аксонов спускаются на ипсилатеральной стороне в виде вентрального кортикоспинального тракта. Эти аксоны также синапсы с нижними мотонейронами вентральных рогов. Большинство из них переходят на противоположную сторону спинного мозга (через переднюю белую комиссуру) непосредственно перед синапсией.
Области мозга по Бродману : На этом рисунке показаны области коры головного мозга человека, очерченные Корвинианом Бродманном на основе цитоархитектуры.
Ядра среднего мозга включают четыре моторных тракта, которые отправляют аксоны верхних мотонейронов по спинному мозгу к нижним мотонейронам. Это руброспинальный тракт, вестибулоспинальный тракт, тектоспинальный тракт и ретикулоспинальный тракт.
Функцию нижних мотонейронов можно разделить на две разные группы: латеральный кортикоспинальный тракт и передний корково-спинномозговой тракт. Боковой тракт содержит аксоны верхних мотонейронов, которые синапсируют с дорсальными боковыми нижними мотонейронами, которые участвуют в контроле дистальных конечностей.
Передний кортикоспинальный тракт опускается ипсилатерально в переднем столбе, где аксоны выходят и либо синапсы на вентромедиальных нижних мотонейронах в вентральном роге ипсилатерально, либо рассекаются в передней белой комиссуре, где они синапсируют с вентромедиальными нижними мотонейронами контралатерально.
Вентромедиальные нижние двигательные нейроны контролируют большие постуральные мышцы осевого скелета. Эти нижние моторные нейроны, в отличие от нейронов дорсального латерального отдела, расположены в вентральном роге по всему спинному мозгу.
Пути спинного мозга : На этой схеме путей спинного мозга моторные и эфферентные пути показаны красным цветом, а сенсорные и афферентные пути — синим. На диаграмме представлены следующие двигательные пути: кортикоспинальные тракты (пирамидный тракт) и экстрапирамидные тракты (тектоспинальный тракт не разграничен).
Роль базальных ганглиев в движении
Базальные ганглии отвечают за произвольный моторный контроль, процедурное обучение и движение глаз, а также за когнитивные и эмоциональные функции.
Цели обучения
Опишите роль базальных ганглиев в движении
Основные выводы
Ключевые моменты
- Базальные ганглии широко изучаются в контексте двух заболеваний базальных ганглиев: болезни Парксинсона и болезни Хантингтона.
- Гемибаллизм, двигательное расстройство, возникающее в результате повреждения нейронов в субталамическом ядре, проявляется насильственными движениями рук и ног.
- На движение глаз, являющееся функцией базальных ганглиев, влияет верхний холмик, область мозга, которая направляет движение глаз к определенным точкам в пространстве в ответ на раздражители.
- Считается, что базальные ганглии также играют роль в мотивации.
- В базальных ганглиях,
большинство нейронов используют гамма-аминомасляную кислоту (ГАМК) в качестве нейромедиатора
и оказывают ингибирующее действие на свои
мишени.
Ключевые термины
- гемибаллизм : редкое двигательное расстройство с непроизвольными резкими движениями конечностей.
- произвольное управление двигателем : Акт направления движения с намерением.
- передний мозг : Передняя часть головного мозга, включая головной мозг, таламус и гипоталамус.
- прилежащее ядро : Область в базальной части переднего мозга, ростральная по отношению к преоптической области гипоталамуса. Эта область и обонятельный бугорок вместе образуют вентральное полосатое тело.
Расположение базальных ганглиев
Базальные ганглии (или базальные ядра) представляют собой группу ядер различного происхождения в головном мозге позвоночных, которые действуют как связная функциональная единица.Они расположены в основании переднего мозга и прочно связаны с корой головного мозга, таламусом и другими областями мозга.
Базальные ганглии связаны с множеством функций, включая произвольный моторный контроль, процедурное обучение, относящееся к рутинному поведению или привычкам, таким как бруксизм и движения глаз, а также с когнитивными и эмоциональными функциями.
Базальные ганглии : Расположение базальных ганглиев.
Выбор действия
В настоящее время популярные теории утверждают, что базальные ганглии играют первостепенную роль в выборе действий.Выбор действия — это решение, какое из нескольких возможных действий выполнить в данный момент.
Экспериментальные исследования показывают, что базальные ганглии оказывают тормозящее влияние на ряд моторных систем, и что снятие этого торможения позволяет моторной системе стать активной. На переключение поведения, происходящее в базальных ганглиях, влияют сигналы от многих частей мозга, включая префронтальную кору, которая играет ключевую роль в исполнительных функциях.
Механизм
Величайшим источником понимания функций базальных ганглиев стало изучение двух неврологических расстройств, болезни Паркинсона и болезни Хантингтона. Для обоих этих расстройств характер повреждения нервной системы хорошо изучен и может коррелировать с возникающими в результате симптомами.
Болезнь Паркинсона связана с большой потерей дофаминергических клеток в черной субстанции. Болезнь Хантингтона связана с массовой потерей колючих нейронов среднего размера в полосатом теле.
Симптомы этих двух заболеваний практически противоположны: болезнь Паркинсона характеризуется постепенной потерей способности инициировать движение, тогда как болезнь Хантингтона характеризуется неспособностью предотвратить непреднамеренное движение частей тела.
Примечательно, что, хотя оба заболевания имеют когнитивные симптомы, особенно на поздних стадиях, наиболее заметные симптомы связаны со способностью инициировать и контролировать движения. Таким образом, оба классифицируются в первую очередь как двигательные расстройства.
Другое двигательное расстройство, называемое гемибаллизмом, может быть результатом повреждения, ограниченного субталамическим ядром. Гемибаллизм характеризуется резкими и неконтролируемыми раскачивающими движениями рук и ног.
Функция движения глаз
Одна из наиболее интенсивно изучаемых функций базальных ганглиев — их роль в управлении движениями глаз. На движение глаз влияет разветвленная сеть областей мозга, которые сходятся в области среднего мозга, называемой верхним холмиком (SC).
СК представляет собой слоистую структуру, слои которой образуют двумерные ретинотопные карты зрительного пространства. Удар нейронной активности в глубоких слоях SC заставляет глаз двигаться к соответствующей точке в пространстве.
Мотивация
Хотя роль базальных ганглиев в моторном контроле очевидна, есть также много указаний на то, что они участвуют в контроле поведения более фундаментальным образом, на уровне мотивации. При болезни Паркинсона способность выполнять компоненты движения не сильно страдает, но мотивационные факторы, такие как голод, не могут заставить движения инициироваться или переключаться в нужное время.
Обездвиженность пациентов с болезнью Паркинсона иногда описывается как паралич воли. Иногда у этих пациентов наблюдается феномен, называемый парадоксальной кинезией, при котором человек, который в противном случае неподвижен, реагирует на чрезвычайную ситуацию скоординированно и энергично, а затем снова теряет подвижность после ее исчезновения.
Роль в мотивации лимбической части базальных ганглиев — прилежащего ядра (NA), вентрального паллидума и вентральной тегментальной области (VTA) — особенно хорошо установлена.Тысячи экспериментальных исследований вместе демонстрируют, что дофаминергическая проекция от VTA к NA играет центральную роль в системе вознаграждения мозга.
Было показано, что многие вещи, которые люди находят полезными, включая наркотики, вызывающие привыкание, вкусную еду и секс, вызывают активацию дофаминовой системы VTA. Повреждение NA или VTA может вызвать состояние глубокого оцепенения.
Нейротрансмиттеры
В большинстве областей мозга преобладающие классы нейронов используют глутамат в качестве нейромедиатора и оказывают возбуждающее действие на свои мишени.Однако в базальных ганглиях подавляющее большинство нейронов используют гамма-аминомасляную кислоту (ГАМК) в качестве нейромедиатора и оказывают тормозящее действие на свои мишени.
Входы коры и таламуса в полосатое тело и субталамическое ядро являются глутаматергическими, но выходы из полосатого тела, паллидума и ретикулированной части черной субстанции используют ГАМК. Таким образом, после первоначального возбуждения полосатого тела во внутренней динамике базальных ганглиев преобладают торможение и растормаживание.
Другие нейротрансмиттеры обладают важным модулирующим действием. Дофамин используется в проекции компактной части черной субстанции на дорсальное полосатое тело, а также в аналогичной проекции из вентральной тегментальной области на вентральное полосатое тело (прилежащее ядро).
Ацетилхолин также играет важную роль, поскольку он используется как несколькими внешними входами в полосатое тело, так и группой интернейронов полосатого тела. Хотя холинергические клетки составляют лишь небольшую часть от общей популяции, полосатое тело имеет одну из самых высоких концентраций ацетилхолина среди всех структур мозга.
Основные цепи базальных ганглиев : На этой схеме показаны основные цепи базальных ганглиев. Два коронарных среза были наложены друг на друга, чтобы включить вовлеченные структуры базальных ганглиев. Знаки + и — на концах стрелок указывают, является ли этот путь возбуждающим или тормозящим, соответственно, по действию. Зеленые стрелки относятся к возбуждающим глутаматергическим путям, красные стрелки относятся к тормозным ГАМКергическим путям, а бирюзовые стрелки относятся к дофаминергическим путям, которые являются возбуждающими на прямом пути и ингибирующими на непрямом пути.
Модуляция движения мозжечком
Мозжечок важен для управления моторикой, в частности координации, точности и времени, а также для некоторых форм моторного обучения.
Цели обучения
Опишите роль мозжечка в модуляции движения
Основные выводы
Ключевые моменты
- Мозжечок представляет собой структуру с параллельными бороздками в нижней части мозга, содержащую очень регулярное клеточное расположение клеток Пуркинье, гранулярных клеток и других типов клеток.
- Мозжечок приспосабливается к изменениям сенсомоторных отношений, возможно, функционируя так, как в теории Марра-Альбуса: сильные входные сигналы от одного лазящего волокна служат обучающим сигналом для изменения силы импульсов от соответствующей группы параллельных волокон.
- Были идентифицированы четыре принципа функции мозжечка. К ним относятся: обработка с прогнозированием, дивергенция и конвергенция, модульность и пластичность.
Ключевые термины
- Клетки Пуркинье : класс ГАМКергических нейронов, расположенных в мозжечке.
- моховые волокна : один из основных источников поступления в мозжечок таких источников, как кора головного мозга.
- гранулярные клетки : Эти клетки получают возбуждающий сигнал от мшистых волокон, которые происходят из ядер моста.
Мозжечок — это область мозга, которая играет важную роль в управлении моторикой. Он также может участвовать в некоторых когнитивных функциях, таких как внимание и речь, а также в регулировании реакций страха и удовольствия, но его функции, связанные с движением, установлены наиболее прочно.Мозжечок не инициирует движение, но способствует координации, точности и точному времени.
Он получает входные данные от сенсорных систем спинного мозга и других частей мозга, включая кору головного мозга, и интегрирует эти входные данные для точной настройки двигательной активности. Из-за этой функции точной настройки повреждение мозжечка не вызывает паралич, а вместо этого вызывает нарушение точных движений, равновесия, осанки и двигательного обучения.
Мозжечок отличается от большинства других частей мозга, особенно коры головного мозга, в отношении способности сигналов двигаться в одном направлении от входа к выходу.Такой режим работы с прямой связью означает, что мозжечок не может генерировать самоподдерживающиеся паттерны нейронной активности, в отличие от коры головного мозга. Однако мозжечок может получать информацию от коры головного мозга и обрабатывать эту информацию, чтобы посылать двигательные импульсы в скелетные мышцы.
Мозжечок : Вид на мозжечок сверху и сзади.
Анатомия мозжечка
Клетки мозжечка : Вид на мозжечок сверху и сзади.
С точки зрения анатомии, мозжечок имеет вид отдельной структуры, прикрепленной к нижней части мозга, расположенной под полушариями головного мозга. Поверхность мозжечка покрыта мелкими параллельными бороздками, разительно контрастирующими с широкими неправильными извилинами коры головного мозга. Эти параллельные бороздки скрывают тот факт, что мозжечок на самом деле представляет собой непрерывный тонкий слой ткани (кора мозжечка), плотно сложенный в виде гармошки.
В этом тонком слое находятся несколько типов нейронов с очень регулярным расположением, наиболее важными из которых являются клетки Пуркинье и гранулярные клетки. Эта сложная нейронная сеть обеспечивает мощную обработку сигналов, но почти весь ее выход направляется на набор небольших глубоких ядер мозжечка, лежащих внутри мозжечка.
Функция
Теория Марра-Альбуса
Помимо своей непосредственной роли в управлении моторикой, мозжечок также необходим для нескольких типов моторного обучения, наиболее заметным из которых является умение приспосабливаться к изменениям в сенсомоторных отношениях.
Было разработано несколько теоретических моделей для объяснения сенсомоторной калибровки с точки зрения синаптической пластичности в мозжечке. Большинство из них происходит от ранних моделей, сформулированных Дэвидом Марром и Джеймсом Альбусом, которые были мотивированы наблюдением, что каждая клетка Пуркинье мозжечка получает два совершенно разных типа входных данных.
Он принимает входные сигналы от тысяч параллельных волокон, каждое из которых по отдельности очень слабое. Однако каждая клетка Пуркинье мозжечка также получает вход от одного единственного лазящего волокна, которое настолько сильное, что единственный потенциал действия восходящего волокна надежно заставляет целевую клетку Пуркинье запускать всплеск потенциалов действия.
Основная концепция теории Марра-Альбуса состоит в том, что восходящее волокно служит обучающим сигналом, который вызывает длительное изменение силы синхронно активируемых параллельных входов волокна. Наблюдения долгосрочной депрессии в параллельных входах волокна обеспечили поддержку теорий такого рода, но срок их действия остаются спорными.
Инсайты о дисфункции мозжечка
Наиболее убедительные ключи к разгадке функции мозжечка были получены при изучении последствий его повреждения.Животные и люди с дисфункцией мозжечка проявляют, прежде всего, проблемы с двигательным контролем. Они по-прежнему способны генерировать двигательную активность, но она теряет точность, производя беспорядочные, нескоординированные или неправильно рассчитанные движения.
Стандартный тест функции мозжечка — дотянуться кончиком пальца до цели на расстоянии вытянутой руки. Здоровый человек будет двигать кончиком пальца по быстрой прямой траектории, тогда как человек с повреждением мозжечка будет двигаться медленно и беспорядочно, с множеством промежуточных коррекций.
Нарушения немоторных функций обнаружить труднее. Таким образом, общий вывод, сделанный несколько десятилетий назад, состоит в том, что основная функция мозжечка — не инициировать движения или решать, какие движения выполнять, а, скорее, калибровать детальную форму движения.
Сравнительная простота и регулярность анатомии мозжечка породила раннюю надежду, что она может подразумевать аналогичную простоту вычислительной функции. Хотя полное понимание функции мозжечка остается неуловимым, по крайней мере четыре принципа определены как важные: 1) обработка с прогнозированием, 2) дивергенция и конвергенция, 3) модульность и 4) пластичность.
- Обработка с прогнозированием: относится к однонаправленному движению сигналов через систему от входа к выходу с очень небольшой периодической внутренней передачей. Это означает, что мозжечок, в отличие от коры головного мозга, не может генерировать самоподдерживающиеся паттерны нейронной активности. Сигналы поступают в схему, обрабатываются каждым этапом в последовательном порядке и затем уходят.
- Дивергенция и конвергенция: около 1000 клеток Пуркинье, принадлежащих микрозоне, могут получать входные данные от 100 миллионов параллельных волокон и фокусировать свой собственный выход на группу из менее чем 50 глубоких ядерных клеток.Таким образом, мозжечковая сеть получает скромное количество входных данных, очень интенсивно обрабатывает их через свою строго структурированную внутреннюю сеть и отправляет результаты через очень ограниченное количество выходных ячеек.
- Модульность: мозжечковая система функционально разделена на независимые модули. Все модули имеют одинаковую внутреннюю структуру, но с разными входами и выходами. Выходные данные одного модуля не оказывают значительного влияния на работу других модулей.
- Пластичность: Синапсы между параллельными волокнами и клетками Пуркинье, а также синапсы между мшистыми волокнами и глубокими ядерными клетками чувствительны к изменению их прочности.Влияние каждого параллельного волокна на ядерные клетки можно регулировать. Такое расположение дает огромную гибкость для точной настройки отношений между входами и выходами мозжечка.
Функции мозжечка при объединении движений
Мозжечок использует обработку с прямой связью и модульность для обработки информации.
Цели обучения
Описать функции мозжечка при объединении движений
Основные выводы
Ключевые моменты
- Функцию мозжечка можно описать принципами прямой обработки и модульности.
- Обработка с прогнозированием означает, что сигналы движутся в одном направлении через мозжечок от входа к выходу.
- Модульность описывает модульную природу системы мозжечка, при которой модули схожей структуры функционируют относительно независимо. Модули состоят из кластеров нейронов с общими входами, но разными выходами.
Ключевые термины
- purkinje : клетки Пуркинье представляют собой класс ГАМКергических нейронов, расположенных в коре мозжечка.Это одни из самых крупных нейронов в человеческом мозге с замысловатой дендритной ветвью, характеризующейся большим количеством дендритных шипов.
- обработка с прогнозированием : свойство некоторых нейронных цепей, в которых сигналы перемещаются по системе в одном направлении от входа к выходу с очень небольшой периодической внутренней передачей.
- микрозона : группа клеток Пуркинье, у всех одинаковое соматотопическое рецептивное поле. Микрозоны содержат порядка 1000 клеток Пуркинье каждая, расположенных в виде длинной узкой полосы и ориентированных перпендикулярно кортикальным складкам.
Функция мозжечка
Обработка с прогнозированием
Мозжечок отличается от большинства других частей мозга тем, что обработка сигналов почти полностью прямая, то есть сигналы перемещаются по системе в одном направлении от входа к выходу с очень небольшой периодической внутренней передачей.
Небольшое количество повторений, которое действительно существует, состоит из взаимного торможения; нет взаимно возбуждающих цепей. Такой режим работы с прямой связью означает, что мозжечок, в отличие от коры головного мозга, не может генерировать самоподдерживающиеся паттерны нейронной активности.
Сигналы поступают в цепь, обрабатываются каждым этапом в последовательном порядке и затем уходят. Как писали Экклс, Ито и Сентаготаи: «Это исключение в конструкции всех возможных реверберационных цепей возбуждения нейронов, несомненно, является большим преимуществом в работе мозжечка как компьютера, потому что то, что остальная нервная система требует от нервной системы. cerebellum, по-видимому, не является каким-то выходным сигналом, выражающим работу сложных реверберационных цепей в мозжечке, а скорее является быстрой и четкой реакцией на ввод любого конкретного набора информации.”
Дивергенция и конвергенция
Клетки мозжечка : Поперечный разрез листка мозжечка, показывающий его основные типы клеток и связи.
В мозжечке человека информация от 200 миллионов входов мшистых волокон расширяется до 40 миллиардов гранулярных клеток, чьи параллельные выходы волокон затем сходятся на 15 миллионов клеток Пуркинье. Из-за того, как они выстроены в продольном направлении, около 1000 клеток Пуркинье, принадлежащих микрозоне, могут получать входные данные от 100 миллионов параллельных волокон и фокусировать свой собственный выход на группу из менее чем 50 глубоких ядерных клеток.
Таким образом, мозжечковая сеть получает скромное количество входных данных, очень интенсивно обрабатывает их через свою строго структурированную внутреннюю сеть и отправляет результаты через очень ограниченное количество выходных ячеек.
Модульность
Мозжечковая система функционально разделена на более или менее независимые модули, количество которых, вероятно, исчисляется от сотен до тысяч. Все модули имеют одинаковую внутреннюю структуру, но разные входы и выходы.
Модуль (многозональный микрокамер в терминологии Аппса и Гарвича) состоит из небольшого кластера нейронов в нижнем оливарном ядре, набора длинных узких полосок клеток Пуркинье в коре мозжечка (микрозон) и небольшого кластера нейроны в одном из глубоких ядер мозжечка.
Различные модули используют входные данные от мшистых волокон и параллельных волокон, но в остальном они работают независимо. Выходные данные одного модуля не оказывают значительного влияния на работу других модулей.
Пластичность
Синапсы между параллельными волокнами и клетками Пуркинье и синапсы между мшистыми волокнами и глубокими ядерными клетками чувствительны к изменению их силы. В одном модуле мозжечка входные сигналы от целого миллиарда параллельных волокон сходятся на группу из менее чем 50 глубоких ядерных клеток, и влияние каждого параллельного волокна на эти ядерные клетки регулируется.Такое расположение дает огромную гибкость для точной настройки отношений между входами и выходами мозжечка.
Зоны и микрозоны в мозжечке : На этой схематической иллюстрации структуры зон и микрозон в мозжечке показаны три уровня увеличения. Эти зоны и микрозоны помогают объяснить модульный характер функции мозжечка. Слева упрощенная иллюстрация того, как выглядела бы кора мозжечка, если бы все складки были распрямлены: вертикальное измерение — это ростро-каудальная ось мозжечка, горизонтальное измерение — это медиолатеральная ось.Зона — это продольно ориентированная полоса коры головного мозга, а микрозона — это тонкая продольно ориентированная часть зоны. Как показано на рисунке справа, дендритные деревья клеток Пуркинье сплющены таким образом, чтобы соответствовать длине микрозоны, а параллельные волокна пересекают микрозоны под прямым углом.
| Дорожки Энни Берк-Доу, PT, MPT, PhD Практикующий физиотерапевт, доцент Санкт-Петербургского университета.Августина для медицинских наук в Сан-Диего, Калифорния Слайд 1: Автомобильные дороги Добро пожаловать в нейроанатомию в физиотерапии. Я доктор Энни Берк-Доу, практикующий физиотерапевт и адъюнкт-профессор Медицинского университета Святого Августина в Сан-Диего, Калифорния. В этой лекции мы рассмотрим организацию двигательной системы, в частности, кортикоспинального тракта и других моторных путей.В конце этого раздела вы должны уметь: описать анатомию и функции спинного мозга, включая ядра и пластинки; обсудить кровоснабжение спинного мозга; определить место происхождения, перекрестия и уровни прекращения кортикоспинального тракта и других моторных путей; описать вегетативную нервную систему, подразделения, волокна, нейротрансмиттеры и регуляцию; дифференцировать верхние и нижние мотонейроны нервной системы; и описать распространенные нарушения походки в неврологии. Слайд 2: Первичная сенсорная и двигательная области Здесь, на слайде 2, показаны основные сенсорные и моторные области.Помните, что эти области расположены по обе стороны от центральной борозды, которая отделяет лобную долю от теменной доли. Первичная моторная кора красного цвета находится в прецентральной извилине, а первичная сенсорная кора — в постцентральной извилине. Есть несколько важных областей коры моторных ассоциаций, которые лежат непосредственно перед первичной моторной корой, включая дополнительную моторную область, выделенную зеленым цветом, и премоторную кору, отмеченную оранжевым цветом. Эти области участвуют в моторном планировании более высокого порядка и проецируются на первичную моторную кору.Точно так же соматосенсорная ассоциативная кора в теменной доле получает входные данные от первичной соматосенсорной коры и также важна для сенсорной обработки более высокого порядка. Слайд 3: Соматотопическая организация Функциональное картирование и исследования повреждений продемонстрировали, что первичная моторная и соматосенсорная кора — это то, что описывается как соматотопически организованная. Соматотопическая организация гарантирует, что информация из определенной области тела представлена в определенной области коры головного мозга.Волокна в пути расположены так, что информация из нижней части тела проходит по определенному пути или части центральной нервной системы, которая отличается от информации из верхних частей тела. Например, информация, относящаяся к нижней части тела, перемещается по волокнам, расположенным медиально в белом веществе спинного мозга, тогда как информация, относящаяся к верхней части тела, перемещается по волокнам, расположенным в боковых частях. Таким образом, считается, что этот путь имеет соматотопическую или пространственную ориентацию.Эта пространственная ориентация гарантирует, что точка происхождения сигнала сохраняется на всем пути до конечной точки завершения пути. Давайте посмотрим на это более внимательно. Слайд 4: Сенсорный и моторный гомункул Изображенные здесь корковые карты — сенсорный гомункул слева (гомункул означает «маленький человек» на латыни) и справа — моторный гомункул — были разработаны, чтобы помочь нам в широком понимании того, что области коры соответствуют областям в теле.Эти изображения используются, чтобы помочь нам в клинической локализации корковой функции. В результате получился гротескно изуродованный человек с непропорционально большими руками, губами и лицом по сравнению с остальным телом. Из-за мелкой моторики и чувствительных нервов, обнаруженных в этих конкретных частях тела, у гомункула они представлены как более крупные. Часть тела с меньшим количеством сенсорных и / или моторных связей с мозгом кажется меньше. Слайд 5: спинной мозг При рассмотрении анатомии спинного мозга спинной мозг делится на серое вещество, изображенное здесь слева, и окружающее белое вещество.Спинной мозг содержит центральное серое вещество в форме буквы «Н» или в форме бабочки, окруженное восходящими и нисходящими столбиками белого вещества, также называемыми канатиками. Сенсорные нейроны ганглиев задних корешков имеют раздвоенные аксоны. Одна ветвь передает сенсорную информацию с периферии, а другая несет информацию через филаменты спинных нервных корешков в дорсальную часть спинного мозга. В центральном веществе находится дорсальный задний рог, который участвует в основном в сенсорной обработке; промежуточная зона, содержащая интернейроны и некоторые специализированные ядра; и вентральный рог, или передний рог, который содержит двигательные нейроны.Моторные нейроны посылают свои аксоны из спинного мозга через нити вентральных нервных корешков. Серое вещество спинного мозга также можно разделить на ядра или, используя другую номенклатуру, интраламинарное, названное Брором Рекседом, с различными функциями, которые будут обсуждены позже. Белое вещество спинного мозга состоит из задних дорсальных столбов, боковых столбов и передних вентральных столбов. Слайд 6: спинной мозг Спинной мозг по своей длине различается по размеру и форме.Белое вещество, состоящее из продольных трактов, является самым толстым на уровне шейки матки здесь, вверху, где большинство восходящих волокон уже вошли в спинной мозг, а большинство нисходящих волокон еще не закончились на своих мишенях, в то время как крестцовый канатик в основном состоит из серого вещества. Кроме того, спинной мозг имеет два увеличения: шейное и пояснично-крестцовое, которые дают начало нервным сплетениям рук и ног, соответственно. В спинном мозге больше серого вещества на шейном и пояснично-крестцовом уровнях, чем на грудном, особенно в вентральных рогах.В грудном канатике представлен латеральный рог, который содержит столбцы промежуточно-боковых клеток. Промежуточно-боковые клеточные столбы существуют на позвоночных уровнях от T1 до L2 и опосредуют всю симпатическую иннервацию тела. Слайд 7: Кровоснабжение спинного мозга Кровоснабжение спинного мозга происходит от ветвей позвоночных артерий и спинномозговых корешковых артерий. Эти позвоночные артерии дают начало передней спинной артерии, которая проходит вдоль вентральной поверхности спинного мозга.Кроме того, две задние спинномозговые артерии отходят от позвоночных или задних нижних мозжечковых артерий и снабжают дорсальную поверхность спинного мозга. Передняя и задняя спинномозговые артерии имеют различное выступание на разных уровнях и образуют спинномозговое артериальное сплетение, окружающее спинной мозг. В позвоночный канал по всей его длине входит 31 сегментарная артериальная ветвь. Большинство ветвей отходят от аорты и снабжают мозговые оболочки. Только от 6 до 10 из них достигают спинного мозга в виде корешковых артерий, вновь возникающих на разных уровнях, в зависимости от человека.Большая корешковая артерия Адамкевича, обычно выступающая артерия, отходящая с левой стороны в любом месте от T5 до L3, но обычно видимая между T9 и T12, обеспечивает основное кровоснабжение поясничного и крестцового отдела спинного мозга. Срединно-грудной отдел, приблизительно от Т4 до Т8, расположен между снабжением поясничной и позвоночной артерии и является уязвимой зоной относительно пониженной перфузии. Эта область очень восприимчива к инфаркту во время торакальной хирургии или других состояний, вызывающих снижение аортального давления.Передняя спинномозговая артерия кровоснабжает примерно две трети спинного мозга, включая передние рога и передние боковые столбики белого вещества. Задние спинномозговые артерии кровоснабжают задний спинной столб и часть задних рогов. Слайд 8: Двигательные системы Когда мы думаем о сложности движений, необходимых для спортивных достижений, игры на музыкальном инструменте или других умелых движений, мы можем понять, что двигательные системы включают сложные сети или нейронные цепи, которые общаются через множество иерархических петель обратной связи.Краткое изложение моторных систем представлено здесь, показаны только основные моторные петли и опущены важные сенсорные входы. Помните, что изображенные мозжечок и базальные ганглии участвуют в этих петлях обратной связи и проецируются обратно в кору головного мозга через таламус. Сами они не проецируются на нижние двигательные нейроны. Ранее мы обсуждали, что в самой коре головного мозга есть многочисленные моторные цепи для управления моторикой, и они проецируются на базальные ганглии, изображенные здесь синим цветом.Базальные ганглии представляют собой массивное серое вещество, которое находится в каждом полушарии глубоко до дна боковых желудочков. Они встроены в белое вещество, которое проецируется через таламус обратно в кору. Базальные ганглии участвуют в подсознательном контроле тонуса скелетных мышц и координации усвоенных моделей движений. В нормальных условиях эти ядра не инициируют определенных движений, но как только движение началось, они будут обеспечивать общий рисунок и ритм, особенно для туловища и проксимальных мышц конечностей.Области головного мозга, включая ассоциативные области коры, такие как дополнительная моторная область, премоторная кора и теменная ассоциативная кора, имеют решающее значение для планирования и формулирования двигательной активности. Поражение этих областей ассоциативной коры может привести к тому, что мы называем апраксией, при которой наблюдается дефицит моторного планирования и исполнения более высокого порядка, несмотря на нормальную силу. Петли обратной связи мозжечка помогают программировать и настраивать движения, контролируемые на сознательном и подсознательном уровнях.Мозжечок уточняет выученные паттерны движений, сравнивая моторные команды с проприоцептивной информацией, и выполняет корректировки, необходимые для плавности движений. Повреждение этих областей может вызвать то, что мы называем атаксией или нарушением порядка, нарушением координации мышц. Слайд 9: нейроны верхнего и нижнего двигателя Напомним, что соматические моторные пути включают по крайней мере два мотонейрона: верхний мотонейрон, тело клетки которого находится в обрабатывающем центре центральной нервной системы, и нижний мотонейрон, изображенный здесь синим цветом, тело клетки которого лежит в ядре мозга. стволовые, поскольку они относятся к черепным нервам или в спинном мозге, поскольку они относятся к периферическим нервам.Верхний двигательный нейрон синапсирует с нижним двигательным нейроном и иннервирует одну двигательную единицу в скелетной мышце. Активность верхнего мотонейрона может способствовать или ингибировать нижний мотонейрон. Активация нижнего мотонейрона вызывает сокращение иннервируемой мышцы. Только аксон нижнего мотонейрона выходит за пределы ЦНС. Разрушение или повреждение нижнего мотонейрона устраняет произвольный и рефлекторный контроль над иннервируемой моторной единицей. Слайд 10: Первичные нисходящие (моторные) пути В этой таблице описаны основные нисходящие двигательные пути по расположению верхних двигательных нейронов, месту назначения, стороне пересечения или перекреста и их роли в двигательной функции.Нисходящие двигательные пути можно разделить на боковые и медиальные двигательные системы в зависимости от их расположения в спинном мозге. Две боковые двигательные системы — это боковой кортикоспинальный тракт и руброспинальный тракт, которые контролируют движения конечностей. Боковой кортикоспинальный тракт особенно важен для быстрого ловкого движения отдельных пальцев или суставов. Большинство этих путей пересекаются от места своего происхождения и спускаются в контралатеральный спинной мозг, чтобы контролировать контралатеральные конечности.Вот почему левая часть мозга контролирует правую часть тела. Четыре медиальные двигательные системы — это передний кортикоспинальный тракт, вестибулоспинальный тракт, ретикулоспинальный тракт и тектоспинальный тракт. Эти пути контролируют проксимальные осевые мышцы и мышцы пояса, участвующие в постуральном тонусе, балансе, ориентировочных движениях головы и шеи и автоматических движениях, связанных с походкой. Медиальные двигательные системы спускаются ипсилатерально или двусторонне. Некоторые опускаются только к нескольким верхним шейным сегментам, как мы увидим на следующих слайдах.Медиальные двигательные системы имеют тенденцию оканчиваться на интернейронах, которые выступают по обе стороны от спинного мозга, контролируя движения, которые затрагивают несколько двусторонних сегментов позвоночника. Из-за этого односторонние поражения медиальной двигательной системы не вызывают явных нарушений. Руброспинальный тракт у человека невелик, и его функция изучена недостаточно. Считается, что он играет роль в моторной функции после повреждения кортикоспинального тракта, а также в сгибательной или декортикальной позе верхних конечностей, что наблюдается при поражениях выше уровня красных ядер. Слайд 11: Соматотопическая организация проекций медиальной и латеральной двигательной системы на клетки переднего рога Когда мы смотрим на соматотопную организацию спинного мозга, здесь мы видим медиальную и боковую двигательные системы, проецирующиеся на клетки переднего рога. Наши боковые моторные системы — кортикоспинальный и руброспинальный тракты, выделенные красным цветом, — проецируются на латеральные клетки переднего рога, в то время как медиальные моторные системы — передние кортикоспинальные, вестибулоспинальные, ретикулоспинальные и тектоспинальные, отмеченные синим цветом, — проецируются на медиальные клетки переднего рога.Боковые клетки переднего рога контролируют дистальные мышцы конечности, а медиальные клетки переднего рога контролируют проксимальную мускулатуру туловища. Слайд 12: Кортикоспинальный тракт Теперь посмотрим на нисходящие двигательные тракты. Кортикоспинальный тракт — это клинически важный нисходящий моторный путь, который состоит из латеральной и передней части. Этот путь иногда называют пирамидной системой из-за его связи с мозговыми пирамидами.Кортикоспинальный тракт обеспечивает произвольный контроль над скелетными мышцами. Эта система начинается в пирамидных клетках первичной моторной коры, и аксоны этих верхних мотонейронов спускаются в ствол головного мозга и спинной мозг в синапсы на нижних моторных нейронах, изображенных красным, которые контролируют скелетные мышцы. Слайд 13: Кортикоспинальный тракт Если посмотреть на путь кортикоспинального тракта, более половины волокон берут начало в первичной моторной коре, области 4 Бродмана, прецентральной извилины.Остальные возникают из премоторной и дополнительной моторной области 6 или из областей 3, 1, 2, 5 и 7 теменной доли. Нейроны первичной моторной коры, участвующие в кортикоспинальном тракте, расположены в основном в корковом слое 5. Слой 5 пирамидный. синапсы проекции клеток непосредственно на двигательные нейроны в вентральном или переднем роге спинного мозга, а также на спинномозговые интернейроны. Около 3% кортикоспинальных нейронов представляют собой гигантские пирамидные клетки, называемые клетками Беца, которые являются крупнейшими нейронами нервной системы человека.Аксоны из коры входят в верхние части белого вещества головного мозга или в лучистую корону и спускаются к внутренней капсуле. Помните, что помимо кортикоспинального тракта белое вещество головного мозга передает двунаправленную информацию между различными областями коры, а также между корой и глубокими структурами, такими как базальные ганглии, таламус и ствол мозга. Эти пути белого вещества образуют веерообразную структуру, когда входят во внутреннюю капсулу, которая конденсируется до все меньшего и меньшего количества волокон по мере того, как создаются связи с различными подкорковыми структурами.Давайте продолжим следить за трактатом на следующих нескольких слайдах. Слайд 14: внутренняя капсула Опять же, кортикоспинальный тракт начинается в моторной коре и спускается во внутреннюю капсулу. Внутренняя капсула или выступающие волокна связывают кору головного мозга с промежуточным мозгом, стволом головного мозга, мозжечком и спинным мозгом. Все проекционные волокна должны проходить через промежуточный мозг, где аксоны, восходящие к сенсорным областям коры, проходят между аксонами, спускающимися из моторных областей коры.Внутренняя капсула лучше всего видна в этом горизонтальном срезе мозга, в котором правая и левая внутренние капсулы выглядят как наконечники стрел, здесь красного цвета, или две буквы «V», остриями обращенные друг к другу. Обратите внимание, что таламус и хвостатое ядро всегда расположены медиальнее внутренней капсулы, в то время как бледный шар и скорлупа всегда расположены латеральнее внутренней капсулы. Слайд 15: внутренняя капсула Вот схематическое изображение горизонтального вида, показывающего три части внутренней капсулы: переднюю конечность вверху, заднюю конечность внизу и колено.Обратите внимание, что передняя конечность внутренней капсулы отделяет головку хвостатого отростка от бледного шара и скорлупы, а задняя конечность отделяет таламус от бледного шара и скорлупы. Колено, или «колено» на латыни, представляет собой переход между передней и задней конечностями на уровне отверстия Монро. Кортикоспинальный тракт находится в задней части внутренней капсулы, и вы можете видеть, что соматотопическая карта сохранилась, поэтому волокна на лице, буква «F», расположены ближе к переднему краю, а волокна руки (буква «А») и ноги. , буква «L» все больше и больше кзади.Волокна, проходящие из коры головного мозга в ствол головного мозга, включая двигательные волокна лица, называются кортикобульбарными, а не кортикоспинальными, поскольку они проходят из коры головного мозга в ствол головного мозга или луковицу. Несмотря на такое соматотопическое расположение, волокна внутренней капсулы достаточно компактны, поэтому поражения на этом уровне обычно вызывают слабость всего контралатерального тела — лица, руки и ноги. Слайд 16: Кортикоспинальный тракт По мере того, как мы продолжаем спускаться от внутренней капсулы, кортикоспинальные тракты встречаются с церебральными ножками или ножками головного мозга.Белое вещество располагается в брюшной части ножек головного мозга и называется основанием ножек. Волокна кортикоспинального тракта спускаются по вентральным мостам. Они образуют костномозговые пирамиды. Опять же, по этой причине кортикоспинальный тракт иногда называют пирамидным трактом. Переход от продолговатого мозга к спинному мозгу называется шейно-медуллярным переходом и происходит на уровне большого затылочного отверстия. В этот момент около 85% волокон пирамидного тракта пересекаются в пирамидальном перекресте и входят в латеральные столбцы белого вещества спинного мозга, формирующие латеральный кортикоспинальный тракт.Соматотопическое представление присутствует в латеральном кортикоспинальном тракте, при этом волокна, контролирующие верхнюю конечность, расположены медиальнее тех, которые контролируют нижнюю конечность. Наконец, аксоны в латеральном кортикоспинальном тракте входят в центральное серое вещество спинного мозга, чтобы синапсировать с клетками переднего моторного рога. Оставшиеся примерно 15% кортикоспинальных волокон продолжаются в спинной мозг на ипсилатеральной стороне, не пересекаясь, и входят в передние столбы белого вещества, образуя передний кортикоспинальный тракт. Слайд 17: Нисходящие автомобильные тропы Опять же, здесь мы можем видеть боковой путь кортикоспинального тракта, а также руброспинальный тракт, которые отвечают за противоположную моторную функцию. Обратите внимание: справа у человека руброспинальный тракт небольшой и заканчивается шейным канатиком. Считается, что он отвечает за выполнение функций после повреждения кортикоспинального тракта, а также может играть роль в сгибательной или декортикальной позе в верхних конечностях с поражениями выше уровня красного ядра, в котором сохраняется руброспинальный тракт. Слайд 18: Нисходящие автомобильные тропы На этом слайде слева мы видим передний кортикоспинальный тракт, заканчивающийся шейным и верхним грудным канатиками, и он контролирует двустороннюю подмышечную впадину и поясную мышцу, а справа — вестибулоспинальный тракт с медиальными волокнами, заканчивающимися шейным и шейным отделами. верхний грудной спинной мозг и боковые части, которые до сих пор считаются частью медиальной системы, оканчиваются всей спинной мозговой системой.Вестибулоспинальные тракты отвечают за положение головы и шеи (медиальные волокна) и равновесие (боковые волокна). Слайд 19: Нисходящие автомобильные тропы Последние два нисходящих моторных пути — это тектоспинальный тракт, заканчивающийся шейным канатиком, и ретикулоспинальный тракт, заканчивающийся вдоль всего спинного мозга. Тектоспинальный тракт отвечает за координацию движений головы и глаз, что плохо понимается людьми.Ретикулоспинальный тракт отвечает за автоматическую осанку и движения, связанные с походкой. Слайд 20: Вегетативная нервная система (ВНС) В отличие от соматических моторных путей, описанных на предыдущих слайдах, вегетативная нервная система обычно контролирует более автоматические и висцеральные функции организма. Вегетативные эфференты анатомически уникальны. Существует периферический синапс, расположенный в ганглии между центральной нервной системой и эффекторной железой, или гладкой мышцей.В вегетативную нервную систему поступают сенсорные (афферентные) сигналы как в центре, так и на периферии. Однако сама вегетативная нервная система состоит только из эфферентных путей. Слайд 21: Подразделения ANS Вегетативная нервная система имеет два основных отдела: симпатический, также называемый грудопоясничным, который возникает от грудного уровня 1 до поясничного уровня 2 или 3. Симпатический отдел изображен здесь на левой стороне слайда и участвует в основном в «борьбе или функции полета, такие как увеличение частоты сердечных сокращений и артериального давления, расширение бронхов и увеличение размера зрачка.Другой главный отдел, парасимпатический отдел, также называемый краниосакральным отделом, напротив, возникает из ядер черепных нервов и от крестцовых уровней 2-4, и он участвует в функциях «отдыха и переваривания», таких как увеличение желудочной секреции и перистальтики. , замедляя частоту сердечных сокращений и уменьшая размер зрачка. Кишечная нервная система считается третьим отделом вегетативной нервной системы, состоящим из нервного сплетения, расположенного в стенках кишечника, которое участвует в контроле перистальтики и желудочно-кишечных секреций.Преганглионарные нейроны симпатического отдела образуют цепочку, называемую симпатическим стволом, изображенную красным цветом. Симпатический ствол проходит от шейного до крестцового уровней с каждой стороны спинного мозга. Симпатический ствол позволяет симпатическим эфферентам, выходящим только на грудопоясничном уровне, достигать и других частей тела. Парасимпатические преганглионарные волокна возникают из парасимпатических ядер черепных нервов и из парасимпатических ядер крестцового нерва и расположены в сером веществе S2, 3 и 4. Слайд 22: Волокна автономного двигателя Здесь мы можем увидеть соматическую моторную систему розовым цветом по сравнению с вегетативными симпатическими висцеральными моторными эффектами синим цветом. Преганглионарные нейроны симпатического отдела расположены в промежуточно-латеральном столбце клеток в пластинке 7 спинного мозга на уровнях от T1 до L2 или 3. При симпатической стимуляции эфферентные двигательные команды будут перемещаться к симпатическим ганглиям, которые иннервируют постганглионарные волокна в гладкие мышцы. , железы и органы. Слайд 23: Нейротрансмиттеры ANS Если смотреть на нейротрансмиттеры вегетативной нервной системы, синаптическая нейротрансмиссия в холинергических преганглионарных нейронах как в симпатических, так и в парасимпатических ганглиях опосредуется нейротрансмиттером ацетилхолина на никотиновых рецепторах. Симпатическая и парасимпатическая системы различаются постганглионарными нейротрансмиттерами. В симпатическом пост-ганглии здесь, вверху, нейроны выделяют преимущественно норэпинефрин в конечные органы — нашу ткань-мишень — и активируют адренергические альфа- или бета-рецепторы.Парасимпатические постганглионарные нейроны, здесь внизу, выделяют преимущественно ацетилхолин и активируют мускариновые холинергические рецепторы в конечных органах. Норадренергические или адренергические подтипы альфа 1, альфа 2, бета 1, бета 2 и бета 3, а холинергические мускариновые 1, мускариновые 2 и мускариновые 3 подтипы опосредуют различные действия этих нейромедиаторов на конечные органы. Одно заметное исключение из правила симпатической / ацетилхолиновой парасимпатии норадреналина — постганглионарные нейротрансмиттеры в потовых железах, которые иннервируются симпатическими постганглионарными нейронами, выделяющими ацетилхолин. Слайд 24: Постановление ANS Если смотреть на вегетативную нервную систему и ее регуляцию, симпатический и парасимпатический отток контролируется как прямо, так и косвенно высшими центрами, включая гипоталамус, ядра ствола мозга, такие как солитарное ядро, миндалину лимбической системы и несколько других областей лимбической кора. Вегетативные ответы также регулируются афферентной сенсорной информацией, включая сигналы от внутренних рецепторов, таких как хеморецепторы, осморецепторы, терморецепторы и барорецепторы.Эта регуляция регулирует и координирует деятельность вегетативных центров, расположенных в мосту и мозговом веществе, которые регулируют частоту сердечных сокращений, кровяное давление, дыхание и пищеварение. Следует также отметить, что каждая парасимпатическая и симпатическая функции имеют уникальное расположение в гипоталамусе. Слайд 25: Верхний двигательный нейрон против нижнего двигательного нейрона Идея соотношения верхнего мотонейрона и нижнего мотонейрона очень полезна с клинической точки зрения.Специфические признаки связаны с верхним двигательным нейроном и нижним двигательным нейроном, которые помогают локализовать поражения. Помните, что верхний мотонейрон кортикоспинального тракта проецируется из коры головного мозга в нижний мотонейрон, расположенный в переднем роге спинного мозга. Нижние двигательные нейроны, в свою очередь, проецируются через периферические нервы в скелетные мышцы. Идентичная концепция применима к кортикобульбарному тракту и моторным ядрам черепных нервов. Признаки поражения нижних мотонейронов включают мышечную слабость, атрофию, фасцикуляции и гипорефлексию.Фасцикуляции — это аномальные мышечные подергивания, вызванные спонтанной активностью групп мышечных клеток. Примером доброкачественной фасцикуляции, не связанной с повреждением двигательных нейронов, является подергивание век, которое часто возникает после периодов усталости, избытка кофеина и напряжения глаз, например, при чтении в течение длительного времени. Признаки поражения верхних мотонейронов включают мышечную слабость и сочетание повышенного тонуса и гиперрефлексии, иногда называемое спастичностью. Спастичность — это непроизвольное повышение тонуса, зависящее от скорости, которое приводит к сопротивлению мышц движению. Слайд 26: Термины, обычно используемые для описания слабых мест Слабость — одно из важнейших функциональных последствий поражения как верхних, так и нижних мотонейронов. Используются различные термины, которые в клинической практике могут использоваться взаимозаменяемо для описания как тяжести, так и распределения слабости. Парез определяется как слабость или частичный паралич. Примером может быть гемипарез, и клинически вы увидите слабость на одной стороне тела.Плегия определяется как отсутствие движения. Примером может быть гемиплегия, и клинически вы не увидите движения на одной стороне тела. Паралич также определяется как отсутствие движения. Примером может служить паралич руки без движения руки. Паралич — менее точный термин для обозначения слабости или отсутствия движения. Примером может служить паралич лицевого нерва со слабостью или параличом лицевых мышц. Термины, обозначающие местоположение, включают полу-, что означает половину или одну сторону тела; пара — означает обе ноги; моно — означает одну конечность; ди- означает, что обе стороны тела поражены одинаково; и quadri- или tetra- означающие, что задействованы четыре конечности. Слайд 27: Локализация распространенных нарушений походки Нарушения походки могут быть вызваны нарушением функции практически любой части нервной системы, а также некоторыми ортопедическими заболеваниями. Таким образом, тщательное обследование походки является одним из наиболее чувствительных тестов тонкой неврологической дисфункции. Характерные нарушения походки можно увидеть при поражениях определенных систем. Спастическая походка может быть односторонним или двусторонним поражением кортикоспинального тракта и проявляется в виде циркумдукции жестких ног, иногда порезания ног и ходьбы на носках, уменьшения размаха рук, неустойчивого падения в одну сторону, и может проявляться в кортикальном, подкорковом и стволе головного мозга. инфаркты, поражающие верхние пути двигательных нейронов; церебральный паралич; дегенеративные состояния; рассеянный склероз; и травмы спинного мозга.Атаксическая походка может быть локализована на черве мозжечка или других срединных структурах мозжечка, проявляясь в походке с широкой базой, неустойчивостью, шатанием из стороны в сторону, падением в сторону худшей стороны патологии. Тонкий дефицит может быть обнаружен с помощью тандемной пробы Ромберга или прогулки в нетрезвом виде. Головокружительная походка локализуется в вестибулярных ядрах, вестибулярном нерве или полукружных каналах. Похоже на атаксическую походку, широкую и неустойчивую, пациенты будут раскачиваться и падать, когда их просят встать, сложив ноги вместе с закрытыми глазами, что называется признаком Ромберга.Фронтальная походка включает поражения, локализованные в лобных долях или во фронтальном подкорковом белом веществе, и проявляется в виде медленных, шаркающих нейро- или широкомасштабных «магнитных» — ступни едва отрываются от пола — а затем устойчивых, иногда напоминающих паркинсоническую походку. Паркинсоническая походка может быть локализована в черной субстанции или других областях базальных ганглиев и проявляется как медленная, шаркающая и узкая. У клиента будут трудности с началом движения, и он часто будет наклоняться вперед с меньшим размахом руки и поворачиванием единым блоком.Они неустойчивы при ретропульсии, которая при толчке делает несколько шагов назад, чтобы восстановить равновесие. Слайд 28: Локализация распространенных нарушений походки Дискинетическая походка часто может быть локализована в субталамическом ядре или других областях базальных ганглиев. |