Формирование листовой поверхности у сортов яблони на различных вставочных подвоях Текст научной статьи по специальности «Сельское хозяйство, лесное хозяйство, рыбное хозяйство»
Агрономия
ФОРМИРОВАНИЕ ЛИСТОВОИ ПОВЕРХНОСТИ У СОРТОВ ЯБЛОНИ НА РАЗЛИЧНЫХ ВСТАВОЧНЫХ ПОДВОЯХ
Н.Г. КРАСОВА,
доктор сельскохозяйственных наук, заведующая лабораторией сортоизучения яблони,
А.М. ГАЛАШЕВА,
кандидат сельскохозяйственных наук, старший научный сотрудник, ВНИИСПК Россельхозакадемии
Ключевые слова: яблоня, сорт, листовая поверхность, освещённость, плодовые образования.
302530, г. Орёл, п/о Жилина; тел. 8 (0862) 42-11-39; e-mail: [email protected]
Лист является основным органом растения, в котором синтезируются органические вещества, необходимые для роста, развития и плодоношения дерева. У молодых деревьев листья развиваются в основном на ростовых побегах; у плодоносящих
листья располагаются на побегах и плодовых образованиях. В начале вегетации листья развиваются на плодушках, причём очень быстро, достигая максимума к концу цветения. На побегах листовой полог увеличивается по мере их роста, кото-
рый продолжается более длительный период, иногда до середины лета.
Проникновение света в крону определяется размерами дерева, формой кроны, а также густотой листового полога и состоянием листового аппарата. Интенсивность освещённости уменьшается от периферии к середине кроны.
Так, установлено, что проникновение фотосинтетической радиации в крону дерева Мекинтош составило всего 15% от освещённости периферии крон. В других случаях интенсивность освещённости внутри кроны сильнорослого дерева сорта Делишес уменьшалась на 58% на расстоянии около 2 м от вершины дерева [6]. В кроне 4-метровых деревьев сорта Кокс Оранж Пепин освещённость на расстоянии 1 м снижалась на 34% [5]. От освещённости зависит размер листьев, плодов, их окраска и количество. Листья периферии кроны обладают большей фотосинтетической активностью.
В других случаях интенсивность освещённости внутри кроны сильнорослого дерева сорта Делишес уменьшалась на 58% на расстоянии около 2 м от вершины дерева [6]. В кроне 4-метровых деревьев сорта Кокс Оранж Пепин освещённость на расстоянии 1 м снижалась на 34% [5]. От освещённости зависит размер листьев, плодов, их окраска и количество. Листья периферии кроны обладают большей фотосинтетической активностью.
Процесс ассимиляции в глубине загущённой кроны резко ослабляется. Отмечено, что в крону карликовых деревьев проникает в 2,5-4 раза больше солнечной радиации, чем в крону сильнорослых деревьев [2].
Яблоня принадлежит к числу растений, которым не требуется большая интенсивность света. Оптимальной для яблони является освещённость в пределах 0,7-0,9 кал./ см2Чмин. При поступлении на листовую поверхность меньше 0,2 кал./ см2Чмин. продуктивный фотосинтез почти прекращается [1]. Для нормальной работы листьев освещение не должно опускаться ниже 30% от полного солнечного. Недостаток света препятствует образованию полноценного фотосинтетического аппарата листа, что ограничивает ассимиляцию энергии и углекислоты.
В данной работе приводятся результаты изучения величины листовой поверхности и освещённости кро-
Apple, variety, leafsurface, illumination, fruit formations.
Таблица 1
Интенсивность солнечной радиации сортов яблони на различных вставочных подвоях (кал./см2Чмин), 2004-2005 гг
Вставка, подбой (ЕЛ Сорт (А) На высоте 1 м На высоте 2 м Среднее
Г-1 34 (карлик) Болотовское 0,35 0,87 0,86
Орлик 0,84 0,92 0,88
Имрус 0,53 0,78 0,66
среднее 0,74 0,86 0,80
3-17-38 (карлик) Болотовское 0,37 0,89 0,88
Орлик 0,55 0,77 0,66
Имрус 0,57 0,85 0,71
среднее 0,66 0,84 0,75
3-3-72 (пол\карлик) Болотовское 0,63 0,86 0,75
Орлик 0,48 0,63 0,56
Имрус 0,43 0,74 0,58
среднее 0,51 0,74 0,63
Сильнорослый семенной Болотовское 0,67 0,78 0,73
Орлик 0,38 0,50 0,44
Имрус 0,59 0,64 0,62
среднее 0,55 0,64 0,60
НСР 05А=0,06; В=0 ,07; ¿®=0,13
Таблица 2
Интенсивность солнечной радиации и площадь листовой поверхности
кольчаток летних сортов на различных вставочных подвоях (кал. / см2Чмин), 2005 г.
/ см2Чмин), 2005 г.
Вставка, под вой (В) Сорт.А На высоте 1 м № высоте 2 м Среднее Среднее количество листьев на 1 плодовое образование Площадь одного листа, см Площадь веек листьев кольчатки, см
3-3-72 (полугарлик) Л4ланное 0,75 0£8 0.8Î 80 Î9j6 І36.8
№ннее алое D .60 0,76 0,68 9J6 30.9 296,6
ОрЛО ЕИМ 0,39 057 0.46 5J6 250 140,0
ГЬПИрОВИЗ ÛjSÎ о .es 0.65 65 ЗІ .7 т. 3
Мелба 058 0,77 0,68 — — —
Среднее 059 0,75 0.66 75 Î9j6 ІІ3.9
Сильнорослый семенном И4ланное 055 056 Ü .35 60 173 103,3
FbHHee алое 0,48 0,76 0,62 5J8 16,3 945
Орловім 0,42 0J60 0.51 5J8 21,7 125,9
ГЬПИрОЕИ 0,51 ÙjSÎ 0,57 5£i Ш 148,0
Мелба 0,4в 0J62 0,56 — — —
Среднее 0,46 OjSO 0,53 5,7 iU 113,1
НСР А 0,16 0,18
В 0,10 0,11
АБ ¡Щ 025
ны сортов яблони при выращивании на различных вставочных подвоях.
Методика исследований
Исследования проводились в садах ОПХ ВНИИСПК (посадки 19901993 годов). Изучение освещённости кроны проводилось по «Методике определения светового режима в кронах плодовых деревьев» [3], площади листьев — весовым методом [4].
Особенности закладки цветковых почек на кольчатках в зависимости от величины листовой поверхности изучались по вариантам:
■ кольчатки с плодами и с цветковой почкой;
■ кольчатки без плодов с цветковой почкой;
■ кольчатки с плодами без цветковой почки;
■ кольчатки без плодов и без цветковой почки.
Результаты исследований
Освещённость кроны плодового дерева является одним из факторов, обеспечивающих продуктивность фотосинтеза, и в значительной мере определяет количество плодов на ветке и закладку цветковых почек под урожай следующего года.
Освещённость кроны у яблони в немалой степени зависела от особенностей роста сорта и влияния на него вставочного подвоя. Карликовые вставочные подвои способствуют образованию слаборослой кроны (в 20летнем возрасте — высотой 3,0-3,2 м). Интенсивность солнечной радиации в различных зонах кроны таких деревьев составляла 0,80-0,75 кал./см2 ■ мин. в среднем по сортам Болотовс-кое, Имрус, Орлик, что существенно выше в сравнении с деревьями на по-лукарликовых вставках и на сильнорослом семенном подвое (табл. 1). Интенсивность солнечной радиации сортов летнего срока созревания на полукарликовых вставочных подвоях составила 0,66 кал./см2 ■ мин. — на 20% выше, чем на сильнорослом семенном подвое (0,53 кал./см2 ■ мин.). Особенно слабо освещалась крона сильнорослых деревьев на высоте 1 м — 0,39-0,51 кал./см2 ■ мин., что составило 41-54% от освещённости открытой площадки.
От состояния и величины листового аппарата дерева зависит формирование плодов и закладка цветковых почек под урожай следующего года. У летних сортов на полукарли-ковом вставочном подвое 3-3-72 при лучшей освещённости кроны листья на кольчатках были крупные; средняя площадь 1 листа составила от 25,0 см2 у сорта Орловим до 30,9 см2 у сорта Раннее алое при среднем количестве листьев у этих сортов от 5,6 до 9,6 (табл. 2).
У летних сортов на полукарли-ковом вставочном подвое 3-3-72 при лучшей освещённости кроны листья на кольчатках были крупные; средняя площадь 1 листа составила от 25,0 см2 у сорта Орловим до 30,9 см2 у сорта Раннее алое при среднем количестве листьев у этих сортов от 5,6 до 9,6 (табл. 2).
На сильнорослом семенном подвое при освещённости кроны 0,53 кал./см2 ■ мин. у сортов листья были мельче при меньшей общей листовой
Агрономия
Я
к
интенсивность солнечной радиации площадь листовой поверхности кольчатки
Желанное Раннее Папировка Орловим алое
Рисунок 1. Интенсивность солнечной радиации и площадь листовой поверхности кольчаток сорта Орловим на полукарликовом вставочном
подвое 3-3-72
Таблица 3
Облиственность кольчаток сортов яблони на полукарликовых вставочных подвоях
Оорт Вставочный подвой Варианты’ Среднее количество листьев на 1 плодовое образование Площадь одного листа, с» Площадь веек листьев колывтки, см
ГЬнять ИсЗЄЕЗ 3-4-98 1 60 14,9 1010
2 6.4 12.4 79.4
3 60 93 55,2
4 32 90 29,0
среднее 50 13,4 66,2
Первинка 3-4-98 1 7)5 16, 0 1210
2 50 18,0 104,4
3 60 30 43,0
4 52 11,1 57,7
среднее 62 13,3 82,9
№лрус 3-4-98 1 92 12,7 1170
2 60 11,0 75,0
3 62 90 61,4
4 3,4 7,3 25,0
среднее 6,4 10,2 69,6
Болотовское 3-4-98 1 32 14,5 1190
2 60 19,5 1170
3 60 11. 4 75.2
4 75.2
4 40 30 34,4
среднее 62 13,5 86,4
Орловский пионер 3-3-72 1 6.4 14.1 90.2
2 60 15,3 1010
3 30 11.2 40,3
4 30 70 25,2
среднее 5,1 11,9 64,2
Орлик 3-3-72 1 11.6 14,1 1630
2 70 11.0 86,0
3 5,4 85 46,0
4 32 50 18,0
среднее 70 90 78,4
Чистотел 3-3-72 1 90 14.7 141,1
2 70 18,3 128,1
3 30 15,0 1292
4 2.6 85 22,1
среднее 60 14,2 105,1
Синап орловский 3-4-98 1 11.0 17,1 188,1
2 62 16.0 99.2
3 50 17,0 95,2
4 30 90 28,0
среднее 6.5 14.8 1020
Среднее по сортам 1 30 14,8 1302
2 60 15,2 93,3
3 60 11.3 68,8
4 35 50 29,9
* Варианты: 1 — кольчатки с плодами и с цветковой почкой;
2 — кольчатки без плодов с цветковой почкой;
3 — кольчатки с плодами без цветковой почки;
4 — кольчатки без плодов и без цветковой почки.
Агрономия
поверхности кольчаток в сравнении с деревьями на полукарликовой вставке.
При слабой освещённости кроны сорта Орловим на полукарликовом вставочном подвое (0,46 кал./см2 ■ мин.) листовая поверхность кольчаток была наименьшей (140 см2) из изученных летних сортов (рис. 1).
Изучение закладки цветковых почек в зависимости от величины листовой поверхности проводили на восьми сортах яблони, выращенных на полукарликовых вставочных подвоях. Выявлена довольно чёткая зависимость закладки цветковых почек от величины листовой поверхности (табл. 3).
Кольчатки, несущие плоды, закладывали цветковую почку при наличии в среднем по сортам 8,8 листьев с общей площадью листовой поверхности 130 см2 (1-й вариант).
Наибольшее количество листьев на кольчатках с плодами и цветковыми почками отмечено у сортов
Орлик и Синап Орловский (11,0-11,6) при высокой общей листовой поверхности кольчаток (163,6-188,1 см2). Кольчатки с достаточной обли-ственностью у этих сортов в год плодоношения закладывают цветковые почки. У Орлика 49,5% и у Синапа Орловского 52,5% кольчаток плодоносят 2-3 года подряд.
Неплодоносящие кольчатки у изученных сортов закладывали цветковые почки (2-й вариант) при листьях с общей площадью 98,8 см2.
При слабой облиственности плодоносящие кольчатки, несущие лишь 6 листьев при среднем размере листа 11,3 см2 (3-й вариант), не закладывали цветковые почки под урожай следующего года. На кольчатках без плодов и не заложивших цветковую почку было самое меньшее количество листьев (3,5) небольшого размера (8,3 см2) общей площадью листьев на кольчатке 29,9 см2.
У сортов на карликовых вставочных подвоях общая тенденция обли-
ственности кольчаток с различной нагрузкой плодами и цветковыми почками сохраняется.
При слабой облиственности даже неплодоносящие кольчатки не закладывают цветковые почки как на карликовых, так и на полукарликовых вставочных подвоях (рис. 2, табл. 4).
Площадь листовой поверхности плодовых образований и их состояние оказывают решающее влияние на формирование урожая. При лучшей облиственности сортов на карликовых вставочных подвоях нагрузка урожая на единицу объёма кроны у них составила 19,5 кг/м3, на полукар-ликах — 15,0 кг/м3. Так, у сорта Имрус на карликовом вставочном подвое 317-38 площадь листьев одной кольчатки составляет 97,1 см2, на полу-карлике 3-4-98 — 69,6 см2. Нагрузка урожая на единицу объёма кроны у этого сорта 21,1 кг/м3 на карлике и 17,2 кг/м3 на полукарлике. Соответственно, выше была и нагрузка урожая на единицу проекции кроны: 24,4 кг/м2 на карлике и 18,8 кг/м2 на полукарлике; на единицу площади поперечного сечения штамба: 1,9 и 1,5 кг/см2.
Выводы
1. Световой режим плодового дерева определяется особенностями роста генотипа и влиянием на него подвоя или вставки. На карликовых вставочных подвоях сорта яблони освещались лучше в сравнении с деревьями на полукарликовой вставке и сильнорослом семенном подвое.
2. Состояние и величина листового аппарата в значительной мере определяет формирование плодов и закладку цветковых почек. Выявлена зависимость закладки цветковых почек от величины листовой поверхности.
3. На кольчатках с достаточной листовой поверхностью одновременно с плодоношением закладывалась цветковая почка при наличии 8-11 листьев при общей листовой поверхности не менее 118-158 см2 в зависимости от слаборослого вставочного подвоя.
4. При общей площади листьев на кольчатке менее 40-60 см2 даже неплодоносящие кольчатки не закладывали цветковые почки.
5. У сортов на слаборослых вставочных подвоях на кольчатках формируются более крупные листья в сравнении с сильнорослыми деревьями этих же сортов.
Таблица 4
Площадь листовой поверхности кольчаток сортов яблони на карликовом
(3-17-38) и полукарликовом (3-4-98) вставочных подвоях
Сорта Вста- вочный подвой Площадь всех листьев 1 кольщтки.ом Нагрузка урожая на единица
с плодами и с цветковой почсой без плодов с цветковой почкой с плодами бег цветковой почки без плодов и без цветковой гт:»-»:и об ъема кроны, кг>л площади проекции кроны ltr.fl : площади попереч- ного сечения штамба, кг/см
ktapyc карлик 1922 1160 42,1 382 21,1 24,4 1,9
п/к 1170 75,0 61,4 250 172 13,3 1.5
Чистотел карлик 1710 15Î0 1150 452 16,7 18,5 1.2
п/к 141,1 128.1 1292 22,1 11.1 17,5 2.7
Первинка карлик 1620 1430 73,0 322 — — —
rvV 1210 104.4 43,0 57,7 — — —
Орловский пионер карлик 107 0 1080 74,0 14,1 2D.0 22,1 1.5
п/к 90,2 1010 40,3 252 163 18,5 2,5
Среднее карлик 1530 1510 77,0 320 195 Î1,8 1.5
п/к 1175 102,1 69,7 325 150 18,3 2,2
1 Еэргант 2 Еарианг 3 нарншг 4 щшшг
мисунок z. закладка цветковых почек у сортов яолони в зависимости от величины листовой поверхности и наличия плодов
закладка цветковых почек у сортов яолони в зависимости от величины листовой поверхности и наличия плодов
Литература
1. Гриненко В. В., Попова В. Т. Оптимизация светового режима в саду // Садоводство. 1979. № 7. С. 31.
2. Завалко Л. Б. Физиологические особенности яблони на слаборослом подвое : автореф. дис. … канд. биол. наук. Краснодар, 1967. С. 23-24.
3. Лукьянов В. М., Денисов А. М. Методика определения светового режима в кронах плодовых деревьев // Сельскохозяйственная биология. 1968. № 4. С. 582-584.
4. Рубин С. С., Данилевская О. М. Определение площади листьев плодовых деревьев // Ботанический журнал. 1957. Т. 42.
№ 5. С. 34.
5. Jackson J. E. 1984. Effects of cropping on tree vigour. Acta Hort. 146 : 83-88.
6. Heinicke D. R. 1966. Characteristics of Mcintosh and Red Delicious apples … Proc. Amer. Soc. Hort. Sci. 89 : 10-13.
Методика оценки фотосинтезирующей поверхности кроны деревьев Текст научной статьи по специальности «Сельское хозяйство, лесное хозяйство, рыбное хозяйство»
УДК 581.1
методика оценки фотосинтезирующей поверхности кроны деревьев
Г.В. АНИСОЧКИН, ст. преподаватель МГУЛ(Г)
(1) ФГБОУ ВПО «Московский государственный университет леса» 141005, Московская обл., г. Мытищи-5, ул. 1-я Институтская, д. 1, МГУЛ
8 903 577 3488; 8 926 976 32 99
В статье приведены результаты разработанного автором способа определения площади листовой поверхности кроны дерев. Способ определения площади листовой поверхности крон включает определение массы кроны дерева, массы выборки, определение площади поверхности листовых пластин выборки. Площадь листовой поверхности кроны дерева в предлагаемом способе автор определяет соотношением совокупной площади листовых пластин и их массы к массе кроны дерева. Совокупную площадь листовых пластин выборки определяют как сумму площадей отсканированных поверхностей отдельных листовых пластин выборки. Площадь поверхности каждого листа выборки определяют при помощи компьютерной программы, которая вычисляет ее по соотношению черных и белых пикселей отсканированных листьев. Соотношение площади фотосинтезирующей поверхности кроны дерева к текущему приросту определяет, какая площадь фотосинтезирующей поверхности требуется данной породе для генерации единицы прироста для конкретных условий места произрастания. В зависимости от этого показателя можно определять и планировать плотность произрастания отдельных деревьев породы в насаждении в соответствии с возрастной динамикой их роста, осуществлять подбор пород, определяя целевое назначение создаваемых или реконструируемых насаждений. Также этот показатель, определяемый предлагаемым способом, позволит более достоверно планировать выбираемый запас в ходе рубок ухода, более точно корректировать оставляемую полноту в насаждении в соответствии с возрастом культивируемой породы. Способ определения площади листовой поверхности кроны дерева, включающий определение площади листовых пластин выборки при помощи компьютерной программы, позволяет определить площадь листовой поверхности кроны дерева с высокой достоверностью.
Способ определения площади листовой поверхности крон включает определение массы кроны дерева, массы выборки, определение площади поверхности листовых пластин выборки. Площадь листовой поверхности кроны дерева в предлагаемом способе автор определяет соотношением совокупной площади листовых пластин и их массы к массе кроны дерева. Совокупную площадь листовых пластин выборки определяют как сумму площадей отсканированных поверхностей отдельных листовых пластин выборки. Площадь поверхности каждого листа выборки определяют при помощи компьютерной программы, которая вычисляет ее по соотношению черных и белых пикселей отсканированных листьев. Соотношение площади фотосинтезирующей поверхности кроны дерева к текущему приросту определяет, какая площадь фотосинтезирующей поверхности требуется данной породе для генерации единицы прироста для конкретных условий места произрастания. В зависимости от этого показателя можно определять и планировать плотность произрастания отдельных деревьев породы в насаждении в соответствии с возрастной динамикой их роста, осуществлять подбор пород, определяя целевое назначение создаваемых или реконструируемых насаждений. Также этот показатель, определяемый предлагаемым способом, позволит более достоверно планировать выбираемый запас в ходе рубок ухода, более точно корректировать оставляемую полноту в насаждении в соответствии с возрастом культивируемой породы. Способ определения площади листовой поверхности кроны дерева, включающий определение площади листовых пластин выборки при помощи компьютерной программы, позволяет определить площадь листовой поверхности кроны дерева с высокой достоверностью.
Ключевые слова: площадь листовой поверхности, выборка, дерево, лист, крона, модельное дерево, масса кроны дерева, площадь кроны дерева.
Леса России занимают 69 % ее суши. Они выполняют важную, планетарного значения природоохранную и экологическую функцию. В лесах России сосредоточено более 25 % хвойной древесины. Важным фактором являются природно-климатические условия России, обладающие огромным потенциалом, способствующим произрастанию высокопол-нотных насаждений. В настоящее время в России ведется интенсивное лесопользование, ряд насаждений пройден рубками неоднократно. Возобновления лесов в европейской части России идет очень медленно.
В лесах России сосредоточено более 25 % хвойной древесины. Важным фактором являются природно-климатические условия России, обладающие огромным потенциалом, способствующим произрастанию высокопол-нотных насаждений. В настоящее время в России ведется интенсивное лесопользование, ряд насаждений пройден рубками неоднократно. Возобновления лесов в европейской части России идет очень медленно.
Вблизи крупных промышленных центров леса рекреационного назначения находятся в плачевном состоянии из-за плохой экологии и сильной рекреационной нагрузки, вызванной вследствие интенсивного посещения населением. Отрицательная динамика развития наблюдается у некоторых насаждений, находящихся вдали от индустриальных центров. Так, например, известны проблемы Теллермановских дубрав, над решением которых вот уже не один год работают сотрудники
Института лесоведения РАН. В сложившейся ситуации важнейшей задачей лесоустройства и лесного хозяйства является планирование лесных территорий, подбор пород с большей устойчивостью в насаждениях рекреационного назначения и пород, обладающих наибольшим потенциалом роста в насаждениях эксплуатационного назначения. Для решения этих задач проведены исследования в области определения потенциала продуктивности отдельных пород.
Как нам известно, деревья, растущие на нашей планете, потребляя в процессе жизненного цикла углекислый газ, выделяют кислород, накапливают в себе углерод в виде древесины, который после завершения жизненного цикла дерева разлагается, распадаясь на углекислый газ и минеральные вещества, замыкая, таким образом, круговорот углерода в природе.
Факторы, влияющие на интенсивность роста определенной породы, весьма многочисленны: биологический, климатический, почвенный, агротехнический. Выразить чис-
ловым значением влияние каждого фактора на рост насаждения сложно, так как каждый фактор является функцией нескольких элементов. При решении этой задачи перечисленные факторы для каждой породы можно представить условно линейной связью с продуктивностью. По исследованиям Л.А. Иванова (1959 г.), обнаружена прямая связь между массой листьев (хвои) и течением жизненных процессов. Наибольший вес листьев (хвои), а следовательно, и наибольший расход влаги, интенсивность фотосинтеза, дыхания и других процессов наблюдается у всех древесных пород в возрасте кульминации текущего прироста. Возникает вопрос, все ли породы деревьев одинаково выполняют эту функцию в различных условиях произрастания? Вероятнее всего нет, потому как некоторые породы в определенных условиях произрастать вообще не способны, а если и произрастают, то дают минимальный прирост. Однозначно можно сказать, что некоторые породы обладают большей устойчивостью к неблагоприятным факторам, к таким, как подтопление или пересыхание, уплотнение почвы, загазованность воздуха и т. д. Какое количество древесины способна накопить та или иная порода в определенных условиях существования за время своей жизни в различные периоды интенсивности роста? Какое количество кислорода дерево выделяет в эти периоды?
Соотношением площади фотосинтези-рующей поверхности кроны дерева к текущему приросту определим некий коэффициент J. Иными словами, какая площадь фотосин-тезирующей поверхности необходима данной породе для генерации единицы прироста в определенных условиях. В зависимости от этого показателя находится плотность произрастания отдельных деревьев породы в насаждении, полнота и запас интересуемой породы. Также по предлагаемому показателю, оценивая соотношение количества выделяемого кислорода и прирост древесины, можно корректировать возраст рубки той или иной породы в конкретных условиях.
Сравнивая породы по коэффициенту У, можно определить потенциал или перспективность породы относительно сравни-
ваемой. выб.
выб.
кр.
м.
(1)
выб.
где Sкр — площадь листовой поверхности кро
ны дерева;
М — масса листьев кроны дерева;
Seыб. — сумма площадей листовых пластин листьев выборки;
М г — масса листьев выборки.
выб. А
Определение площади листовой поверхности кроны дерева может быть описано следующим образом. Массу листьев кроны дерева делим на партии. Массу каждой партии определяем взвешиванием при помощи маятникового безмена. Массу листьев кроны дерева устанавливаем сложением полученных масс партий. Отбираем из массы листьев кроны в выборку 100 листовых пластин. Определяем массу 100 листовых пластин взвешиванием на электронных весах. Во избежание погрешностей, которые могут возникнуть из-за транспи-рации, взвешивание массы кроны и выборки нужно производить одновременно. Каждую листовую пластину из выборки кладем на рабочую поверхность сканирующего устройства. Производим сканирование ее поверхности. Сканируемое изображение освещается белым светом, получаемым, как правило, от флуоресцентной лампы сканера. Отраженный свет через редуцирующую (уменьшающую) линзу попадает на фоточувствительный полупроводниковый элемент, называемый прибором с зарядовой связью. Каждый пиксель строки
сканирования изображения соответствует определенным значениям напряжения на приборе с зарядовой связью. Полученные значения напряжения преобразуют в цифровую форму либо через аналого-цифровой преобразователь (для полутоновых сканеров), либо через компаратор (для двухуровневых сканеров). Черные пиксели отображают площадь поверхности листовой пластины. Белые пиксели отображают площадь растра сканирования, не занятого площадью поверхности листовой пластины. Таким образом, последовательно сканируем каждую строку изображения и получаем растр изображения. Сохраняем полученный растр изображения в файл в графическом формате. Определяем площадь листовой пластины листа дерева с помощью компьютерной программы следующим образом:
Черные пиксели отображают площадь поверхности листовой пластины. Белые пиксели отображают площадь растра сканирования, не занятого площадью поверхности листовой пластины. Таким образом, последовательно сканируем каждую строку изображения и получаем растр изображения. Сохраняем полученный растр изображения в файл в графическом формате. Определяем площадь листовой пластины листа дерева с помощью компьютерной программы следующим образом:
1. Запускаем компьютерную программу, определяющую площадь.
2. На экране дисплея выбираем команду «1. Загрузить изображение».
3. Выбираем имя файла с растром изображения отсканированного листа.
4. Компьютерная программа считывает файл с растром изображения отсканированного листа для его последующей обработки.
5. На экране дисплея выбираем команду «2. Рассчитать % площади».
6. Компьютерная программа подсчитывает количество черных и белых пикселей отсканированного растра с изображением листовой пластины.
7. Компьютерная программа определяет площадь листовой пластины Х, вычисляя соотношения черных и белых пикселей отсканированного растра с изображением листовой пластины по формуле (2)
x=M°’Sf
(2)
где Л0 — количество черных пикселей;
N — количество белых пикселей;
Sf -площадь отсканированного растра.
8. Последовательно обрабатываем каждую из листовых пластин в выборке.
9. Компьютерная программа после обработки каждой листовой пластины в окне напротив надписи «Площадь в кв. см» показывает величину площади листовой пласти-
ны и сохраняет полученную величину в памяти компьютера.
10. Компьютерная программа суммирует площади листовых пластин в выборке.
В программе существует клавиша «3. Пересчитать площадь в кв. см», которую используют при изменении графического формата сканера или отсканированного изображения.
Описанный способ обеспечивает сокращение времени определения площади листовой пластины, облегчает в целом работу по определению площади листовой поверхности кроны дерева и повышает достоверность полученных результатов, в частности, при обработке выборок с небольшим количеством листьев.
Способ определения площади листовой поверхности кроны дерева, заключающийся в измерении и суммировании площадей листовых пластин выборки, определении массы листовых пластин выборки, массы листовых пластин кроны дерева, при помощи компьютерной программы позволяет определить площадь листовой поверхности кроны дерева с высокой достоверностью.
Отношение площади фотосинтези-рующей поверхности к текущему годовому приросту позволяет понять, какое пространство необходимо дереву для образования оптимального прироста в насаждении, и то же соотношение для лесов рекреационного назначения показывает, какая древесная порода способна выделить большее количество кислорода в атмосферу на единице площади, тем самым определяя ее экологичность.
Библиографический список
1. Анучин, Н.П. Лесоустройство / Н. П. Анучин. — М.: Сельхозиздат, 1962. — 586 с.
П. Анучин. — М.: Сельхозиздат, 1962. — 586 с.
2. Анучин, Н.П. Проблемы лесопользования / Н.П. Анучин. — М.: Лесная промышленность, 1986. — 264 с.
3. Общесоюзные нормативы для таксации лесов / В.В. Загреев, В.И. Сухих, А.З. Швиденко и др. — М.: Колос, 1992. — 495 с.
4. Мелехов, И.С. Лесоводство / И.С. Мелехов — М.: Агро-промиздат. 1989. — 302 с.
5. Моисеев, Н.А. Экономика лесного хозяйства: учебное пособие / Н.А. Моисеев, Г.М. Киселев, М.П. Пименов. — М.: МГУЛ, 2002. — 72 с.
6. Крючков, В.В. Предельные антропогенные нагрузки и состояние экосистем Севера / В.В. Крючков. — М.: Экология. — 1991. — № 3. — С. 28-40.
7. Рубцов, В.В. Адаптационные реакции дуба на дефолиацию / В.В. Рубцов, И.А. Уткина (отв. ред. А.С. Исаев). — М.: Ин-т лесоведения, 2008. — 302 с.
8. Таксация леса: теоретические основы вычислений: учебное пособие / Г.В. Матусевич, Л.В. Стоноженко, Н.Г. Иванов и др. — М.: МГУЛ, 2013. — 181 с.
9. Air Quality Guidelines — Ecological effects of air pollutants // World Health Organization; Regional Office for Europe; ICP/CEN 902/m 71(S). — 29 July, 1985.
10. Харин, О.А. Лесотаксационный справочник / О.А. Ха-рин. — М.: МЛТИ. 1991. — 155 с.
11. Таблицы и модели хода роста и продуктивности насаждений основных лесообразующих пород Северной Ев-
разии / А.З. Швиденко, Д.Г. Щепащенко, С. Нильссон, и др. — М.: 2008. — 886 с.
12. Энциклопедия лесного хозяйства: в 2-х томах. — Т.1. — ВНИИЛМ, 2004. — 416 с.
— Т.1. — ВНИИЛМ, 2004. — 416 с.
13. Биологическая продуктивность и бюджет углерода лиственничных лесов северо-востока России: монография / Д.Г. Щепащенко, А.З. Швиденко, В.С. Шалаев. — М.: МГУЛ, 2008. — 296 с.
14. Опыт агрегированной оценки основных показателей биопродуктивного процесса и углеродного бюджета наземных экосистем России. Запасы растительной органической массы / А.З. Швиденко, С Нильссон, В.С. Столбовой и др. — М.: Экология, 2000. — № 6. — С. 403-410.
TECHNIQUE OF ESTIMATION OF TREE CROWN PHOTOSYNTHESIZING SURFACE Anisochkin G.V., Senior Lecturer MSFU(1)
[email protected] (1) Moscow State Forest University (MFSU), 1st Institutskaya st., 1, 141005, Mytischi, Moscow reg., Russia, +7 903 577 3488; +7 926 976 32 99
The method of estimation of tree crown leaf surface is demonstrated in the article. The method includes the estimation of tree crown mass and sample mass, and the calculation of leaf surface of the sample. The author calculates the leaf surface using the ratio of the total leaf surface and leaf mass to the tree crown mass. The leaf surface, in its turn, is the sum of surfaces of separated leaves, which were taken from the sample and scanned. Special software was used to calculate the surface area of each leaf, using the proportion of black and white pixels in each scan. The ratio of tree crown photosynthesizing area to the current growth shows what part of photosynthesizing area is required for the tree species under analysis to generate the growth unit in certain conditions of the site. Using this ratio the researcher can calculate and plan the growth density of the separate trees of the species according to the age dynamics of their growth. He can also select certain tree species taking into account the purpose why the stands are created or reconstructed. The ratio is also suitable to calculate the wood volume which can be taken during improvement thinning and to regulate the stand density considering the age of the cultivated species. The method to calculate the tree crown leaf surface, which includes the computer calculation of leaf surface areas, allows to calculate the tree crown leaf surface with high preciseness.
The ratio is also suitable to calculate the wood volume which can be taken during improvement thinning and to regulate the stand density considering the age of the cultivated species. The method to calculate the tree crown leaf surface, which includes the computer calculation of leaf surface areas, allows to calculate the tree crown leaf surface with high preciseness.
Key words: leaf surface area, sample, tree, leaf, crown, model tree, tree crown mass, tree crown weight..
References
1. Anuchin N.P. Lesoustroystvo [Forest Inventory]. Moscow: 1962. 586 p.
2. Anuchin N.P. Problemy lesopol’zovaniya [Forest Use Problems]. Moscow: Lesnaya promyshlennost’, 1986, 264 p.
3. Zagreev V.V., Sukhikh V.I., Shvidenko A.Z., Gusev N.N., Moshkalev A.G. Obshchesoyuznye normativy dlya taksatsii lesov [Union Standards for Forest Taxation]. Moscow: Kolos, 1992, 495 p.
4. Melekhov I.S. Lesovodstvo [Forestry]. Moscow: Agropromizdat, 1989, 302 p.
5. Moiseev N.A., Kiselev G.M., Pimenov M.P. Ekonomika lesnogo khozyaystva. Uchebnoe posobie [Economy of Forestry. StudentsManual]. Moscow: MGUL, 2002, 72 p.
6. Kryuchkov, V.V. Predel’nyeantropogennye nagruzkiisostoyanie ekosistemSevera [Anthropogenic Load and Northern Ecosystems Conditions]. Moscow: Ekologiya, 1991, no 3, pp. 28-40.
7. Rubtsov V.V., Utkina I.A. Adaptatsionnye reaktsii duba na defoliatsiyu [Adaptive Reaction Oak On Defoliation]. Moskva, 2008, 302 p.
8. Matusevich GV Stonozhenko LV Ivanov NG and etc..; Taksatsiya lesa: teoreticheskie osnovy vychisliniy : uchebnoe posobie [Forest Taxation: the theoretical foundations of computing: Students Manual] The Ministry of Education and Science of the Russian Federation; MSFU. Moscow: MSFU, 2013, 181 p.
Moscow: MSFU, 2013, 181 p.
9. Air Quality Guidelines — Ecological effects of air pollutants [Ecological effects of air pollutants] World Health Organization ; Regional Office for Europe ; ICP/CEN 902/m 71(S). 29 July, 1985.
10. Kharin O.A. Lesotaksatsionnyy spravochnik [Forest Taxation Guide]. Moscow: MLTI. 1991. 155 p.
11. Shvidenko A.Z., Shchepashchenko D.G., Nil’sson S., Buluy Yu.I. Tablitsy i modeli khoda rosta i produktivnosti nasazhdeniy osnovnykh lesoobrazuyushchikh porod Severnoy Evrazii [Growth and Productivity Tables and Models for Northern Eurasia Main Forest Forming Species]. Moscow, 2008, 886 p.
12. Entsiklopediya lesnogo khozyaystva [Encyclopedia of Forestry]. T.1. VNIILM, 2004, 416 p.
13. Shchepashchenko D.G., Shvidenko A.Z., Shalaev V.S. Biologicheskaya produktivnost’ i byudzhet ugleroda listvennichnykh lesov Severo-Vostoka Rossii: monografiya [Biological Productivity and North-East Russian Broadleaf Forests Carbon Budget: Monograph]. Moscow: MGUL, 2008, 296 p.
14. Shvidenko A.Z., Nilson S., Ctolbovoy V.S. and etc Opyt agregirovannoy otsenki osnovnykh pokazateley bioproduktivnogo protsessa i uglerodnogo byudzheta nazemnykh ekosistem Rossii. Zapasy rastitel’noy organicheskoy massy [Bioproductive Process and Carbon Budget Main Indexes Aggregate Valuation Experience Vegetative Organic Biomass Reserves]. Moscow: Ekologiya, 2000, no. 6, pp. 403-410.
Способ определения площади листовой поверхности плодового дерева
Изобретение относится к плодоводству. Цель изобретения — повышение производительности труда и точности при определении площади листовой поверхности плодовой стенки и индекса ценоза. Определяют среднюю площадь листовой поверхности в единице объема, за который принимают один линейный метр объема плодовой стенки. Для этого метровую линейку помещают в плодовую стенку под разными углами к вертикали и среднюю площадь листовой поверхности в единице объема определяют подсчетом количества листьев, приходящихся на линейный метр, плотности размещения листьев и средней площади одного листа. Затем определяют высоту и ширину плодовой стенки и расстояние между рядами и рассчитывают площадь листовой поверхности и индекс ценоза соответственно на 1 м2 проекции кроны и 1 м2 ценоза по формулам AЪ = A @ .N3.H.10-4 и A = AЪ.D/B, где A @ — средняя площадь одного листа, см2 N — количество листьев, приходящееся на 1 линейный метр объема плодовой стенки, м-1 H — средняя высота плодовой стенки, м D — средняя ширина плодовой стенки, м B — расстояние между рядами, м.
Определяют среднюю площадь листовой поверхности в единице объема, за который принимают один линейный метр объема плодовой стенки. Для этого метровую линейку помещают в плодовую стенку под разными углами к вертикали и среднюю площадь листовой поверхности в единице объема определяют подсчетом количества листьев, приходящихся на линейный метр, плотности размещения листьев и средней площади одного листа. Затем определяют высоту и ширину плодовой стенки и расстояние между рядами и рассчитывают площадь листовой поверхности и индекс ценоза соответственно на 1 м2 проекции кроны и 1 м2 ценоза по формулам AЪ = A @ .N3.H.10-4 и A = AЪ.D/B, где A @ — средняя площадь одного листа, см2 N — количество листьев, приходящееся на 1 линейный метр объема плодовой стенки, м-1 H — средняя высота плодовой стенки, м D — средняя ширина плодовой стенки, м B — расстояние между рядами, м.
Г Г1К1З
: !IfI, hilIl(ГИ I> КИХ
С f Г11 Г,ЛИГ
1672986 А1 ссflс А 01 G 7/00
ГГ!СУДЛС С1Г1Г IIIII.!И КОМИ1Г Т
Г10 ИЗОЬf f- Т F 11ИfIM И O Т КЛСЯТИЯ1Л
ПРИ СКС1Т СССР
ОПИСАНИЕ ИЗОБРЕТЕНИЯ о 4 м О
I QQ
1о
1р
К АВТОРСКОМУ СВИДЕТЕЛЬСТВУ (21) 4727988/15 (22) 07 08.89 (46) 30 08.91.Бюл. N 32 (71) Научно-производственное обьединение
«Алмалы» (72) В.B,Ìóñòàôèíà (53) 581.45 (088 8) (56) Томилин В.Ф. и Лукьянов В.М. Быстрое определение площади листьев у яблони. Вестник сельскохозяйственной науки, 1972, N
2. с. 107 109. (54) СГ1ОСОБ ОПРЕДF Г1Е НИЯ Г1ЛОЩАДИ
ЛИСТОВОИ ПОВЕРХНОСТИ ПЛОДОВОГО
ДЕРЕВА (57) Изобретение относится к плодоводству, Цель изобретения — повышение производительности труда и точнос1и при определении площади листовои поверхности плодовси гтенки и индекса ценоза. Определяют среднюю площадь листовой поверхности в единице обьема, за который принимают один линеиный метр обьема
Изобретение относится к сельскому хозяиству, в частности к плодоводству для оценки Тотосинтетической деятельности и продуктивности плодовых рас.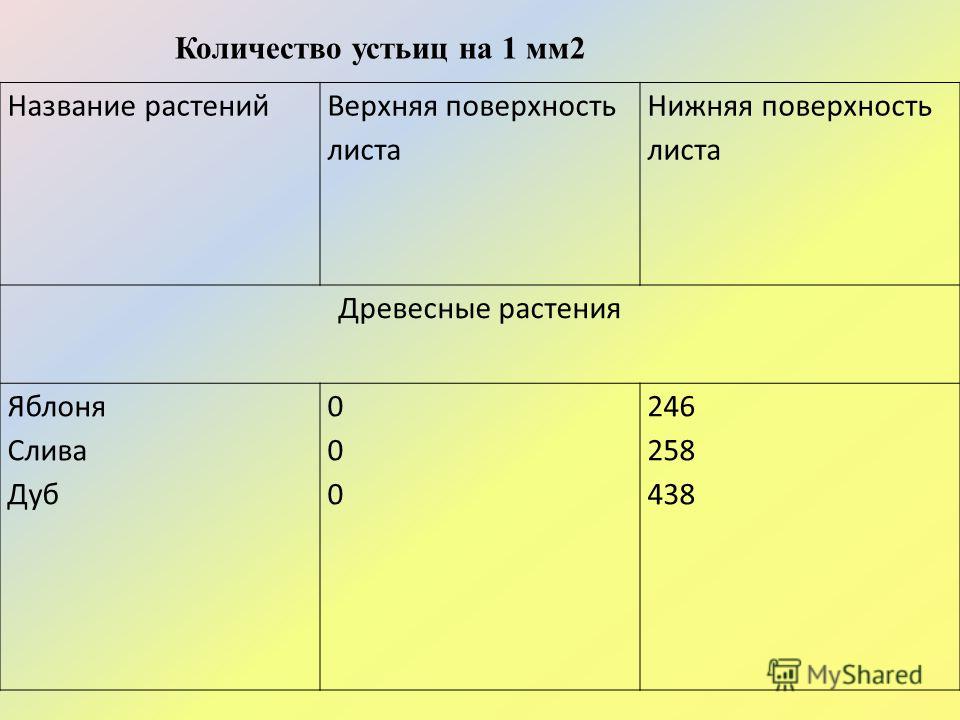 гений.
гений.
Цель изобретения повышение производительности труда и точности при определении площади листовой поверхности плодовои =генки и индекса ценоэа.
При Осуществлении способа определяют среднюю площадь листовой поверхности в единице обьема, за который принимают один линейный метр обьема плодовой стенки Для этого метровую линейку помещают в плодовую стенку под разными углами к вертикали, и среднюю плодовой стенки. Для этого метровую линейку помещают в плодовую стенку под разными углами к вертикали и среднюю площадь листовои поверхности в единице обьема определяют подсчетом количества листьев, приходящихся на линейный метр, плотности размещения листьев и средней площади одного листа. Затем определяют высоту и ширину плодовой стенки и расстояние между рядами и рассчитывают площадь листовой поверхности и индекс ценоза соответс.венно на 1 м проекции кроны и 1 м ценоза по формулам А = Агт х
2 1
N х Н х 10 и А — А к 0/B, где Arz средняя
3 4 1 площадь одного листа, см . N количество
2, листьев, приходящееся на 1 линейный метр обьема плодовой сгенки, м; H — средняя
-1, высота плодовой стенки, м; О — средняя ширина плодовой стенки. м. 8 расстояние между рядами, м площадь листовой поверхности в единице обьема определяют подсчетом количества листьев приходящихся на линейный метр, плотности размещения листьев и средней площади одного листа. Затем определяют высоту и ширину плодовой стенки и расстояние между рядами и рассчитывают площадь листовой поверхности и индекс ценоэа соответственно на 1 м проекции
2 кроны и 1 м ценоза по формулам
А = QgxN хНх10 . (1)
А=А х—
8 (2) где Агт — средняя площадь одного листа, см г
1672986 крайней мере в 3 раза сократить время на определение листовой поверхности плодоBOVI CTP.Hbl, Составитель Г,Бростюк
Редактор Л.Павлова Техред M.Ìoðãåí1àë Корректор О.Кундрик
Заказ 2866 Тираж 379 Подписное
ВНИИГ1И Государственного комитета по изобретениям и открытиям при ГKII1 СС:(P
113035, Москва, Ж 35, Рвугрская наб.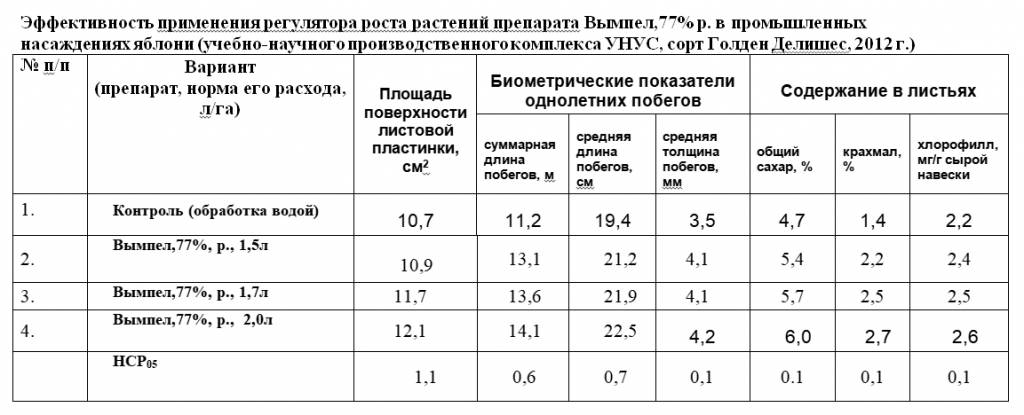 . 4/5
. 4/5
Производственно издательский комбинат «патент» r Ужгород уп Г в вринв 1г11
N — количество листьев, приходящееся на 1 линейный метр объема плодовой стенки, м
Н вЂ” средняя высота плодовой стенки, м;
0 -средняя ширина плодовой стенки, м;
 — расстояние между рядами, м.
Пример, Деревья яблони сорта Голден Далишес в возрасте 5 лет при схеме посадки 0,9 х 3,5 образовали плодовую стену. На участке плодовой стены длиной 1 м определяли весовым методом среднюю площадь случайным образом выбранных листьев А = 28,6 см . Деревянную линейку г помещали в плодовую стену под различными углами к вертикали и каждый раз подсчитывали количество листьев, приходящееся на 1 м линейки, Линейку помещали в плодовую стену так, чтобы охватить все пространство плодовой стены на протяжении 1 м длины. Количество подсчетов может варьировать в зависимости от равномерности расположения листьев, до 20 подсчетов.
Конкретные значения: 9, 10, 13, 3, 5, 10, 11, 13,7, 8,5,7. 13,9,9, 11, 10,9,8. Среднее значение 9,1 м . Далее измеряли среднюю
-1 длину и ширину плодовой стены в данном месте: Н — 1,7 м; D =- 1.3 м. Площадь листовой поверхности плодовой стены на 1 м
2 проекции кроны вычисляли по фоом ле (1):
А =- 28,6 х (9,1) х 7 х 10 =- 3,66 м /м, а на
1 м ценоза — по формуле (2): A =- 3,66 х 1,3
:3,5=1,36м /м .
Предлагаемый способ позволяет упростить операции, повысить точность и по
5 Формула изобретения
Способ определения площади листовой поверхности плодового дерева, включающий определение средней площади листовой поверхности в единице объема дерева с
10 последующим пересчетом на всю листовую поверхность, отличающийся тем, что. с целью повышения производительности труда и точности при определении площади листовой поверхности плодовой стенки и
15 индекса ценоэа, среднюю площадь листовой поверхности определяют на одном линейном метре объема плодовой стенки путем определения средней площади одного листа и плотности расположения листьев, 20 а расчет площади листовой поверхности плодовой стенки и индекс ценоза определяют на 1 м проекции кроны и на 1 м ценоза
2 по формулам соответственно
А — Агг xN хНх10
25 1D
А =Ах—
В где Аа — средняя площадь одного листа, см;
N — количество листьев. приходящихся на 1 линейный метр объема плодовой стенки,м
приходящихся на 1 линейный метр объема плодовой стенки,м
-1, Н средняя высота плодовой стенки. м;
0 — средняя ширина плодовой стенки, м:
 — расстояние между рядами, м.
Регулирование водного режима дерново-подзолистой почвы в условиях плодового питомника и характер распределения влаги в почвенном профиле при различных режимах увлажнения | Романовская
1. Ахмедов А.Д., Темерев А.А., Галиуллина Е.Ю. Надежность систем капельного орошения // Известия нижневолжского агроуниверситетского комплекса: наука и профессиональное образование. 2010. № 3 (19). С. 84–89.
2. Борисова А.А. Вопросы ускоренного производства и повышения выхода привитых саженцев яблони в питомниках средней зоны РСФСР: Автореф. дис. … канд. с.-х. наук. М., 1981. 25 с.
3. Бородычев В.В. Современные технологии капельного орошения овощных культур. Коломна: ФГНУ ВНИИ “Радуга”, 2010. 241 с.
4. Бородычев В.В., Дубенок Н.Н., Лытов М.Н., Белик О.А. Особенности водного режима почвы при капельном орошении сельскохозяйственных культур // Достижения науки и техники АПК. 2009. № 4. С. 22–25.
5. Бородычев В.В., Криволуцкая Н.В., Криволуцкий А.А., Стрижакова Е.А. Продуктивность яблоневого сада интенсивного типа на капельном орошении // Известия нижневолжского агроуниверситетского комплекса: наука и профессиональное образование. 2012. № 3 (27). С. 8–14.
6. Бурмистрова А.Ю. Регулирование водного режима почвы при капельном орошении плодовых питомников в Нечерноземной зоне: Дис. … канд. с.-х. наук. М., 2013. 229 с.
229 с.
7. Григоров М.С., Овчинников А.С., Боровой Е.П., Ахмедов А.Д. Современные перспективные водосберегающие способы полива в Нижнем Поволжье. Волгоград: “Нива”, 2010. 244 с.
8. Грязев В.А. Выращивание саженцев для высокопродуктивных садов. Ставрополь, 1999. 208 с.
9. Дашков В.Н., Радюк. И.И., Дегтеров Д.В., Снежко Э.К. Анализ способов орошения садовых культур в условиях республики Беларусь // Агропанорама. 2010. № 1. С. 11–16.
10. Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: ИД Альянс, 2011. 352 с.
11. Елсайед А.М. Обоснование режимов капельного орошения земляники на дерново-подзолистых почвах: Дис. … к. с.-х. н. М., 2011. 192 с.
12. Иволгин В.С. Оптимизация по критериям ресурсосбережения производственных процессов в плодово-ягодных питомниках: Автореф. дис. … канд. техн. наук. М., 2002. 19 с.
13. Каширская О.В. Совершенствование технологии выращивания посадочного материала для интенсивных садов яблони в ЦЧР: Автореф. дис. … канд. с.-х. наук. М., 2011. 22 с.
14. Колесников В.А. Корневая система плодовых и ягодных культур и методы ее изучения. М., 1962. 191 с.
15. Марков Ю.А. Программа и методика исследований по орошению плодовых и ягодных культур. Мичуринск: ВНИИС им. И.В. Мичурина, 1985. 117 с.
16. Мережко И.М. Качество посадочного материала и продуктивность плодовых насаждений. Киев: “Урожай”, 1991. 152 с.
Мережко И.М. Качество посадочного материала и продуктивность плодовых насаждений. Киев: “Урожай”, 1991. 152 с.
17. Программно-методические указания по агротехническим опытам с плодовыми и ягодными культурами / под ред. Н.Д. Спиваковского. Мичуринск, 1956. 184 с.
18. Методика постановки опытов с плодовыми, ягодными и цветочно-декоративными растениями / под ред. В.А. Комиссарова. М.: Просвещение, 1982. 240 с.
19. Плешаков В.Н. Методика полевого опыта в условиях орошения. Волгоград: ВНИИОЗ, 1983. 148 с.
20. Поспишилова Л. Передвижение воды в почве при дождевании и капельном способе полива: Автореф. дис. … канд. биол. наук. М., 1989. 25 с.
21. Потапов В.А. Программа и методика исследований по вопросам почвенной агротехники в интенсивном садоводстве (методические рекомендации). Мичуринск: ВНИИС им И.В. Мичурина, 1976. 100 с.
22. Рожков В.А., Кузнецова И.В., Рахматуллоев Х.Р. Методы изучения корневых систем растений в поле и лаборатории. М.: Почв. ин-т им. В.В. Докучаева, Моск. гос. ун-т леса, 2004. 40 с.
23. Рожков В.А., Бондарев А.Г., Кузнецова И.В., Рахматуллоев Х.Р. Физические и водно-физические свойства почв. М.: Почв. ин-т им. В.В. Докучаева, Моск. гос. ун-т леса, 2002. 73 c.
24. Рожнов С.И. Разработка технологии капельного орошения саженцев яблони в условиях Нижнего Поволжья: Дис. … канд. с.-х. наук. М., 2004. 204 с.
25. Сабиров М.К., Раззаков М.Д. Влияние капельного орошения на рост и урожайность садов (обзор). Ташкент, 1990. 24 с.
Сабиров М.К., Раззаков М.Д. Влияние капельного орошения на рост и урожайность садов (обзор). Ташкент, 1990. 24 с.
26. Сергиенко А.В. Капельное орошение молодого яблоневого сада на слаборослых подвоях: Дисс. … канд. с.-х. наук. Волгоград, 2008. 226 с.
27. Сычев В.Г. Кузнецова Е.И. и др. Орошение и применение удобрений в Нечерноземной зоне РФ. Монография. М.: ЦИНАО, 2004. 276 с.
28. Технология орошения садов, ягодников и питомников (рекомендации). М.: Агропромиздат, 1987. 61 с.
29. Трунов Ю.В. Минеральное питание клоновых подвоев и саженцев яблони: научное издание. Мичуринск: Изд. Мич. ГАУ, 2004. 175 с.
30. Трунов Ю.В. Минеральное питание и удобрение яблони. Воронеж, Кварта, 2010. 400 с.
31. Шугай П.Ю. Режим капельного орошения школки плодовых культур при повышенных температурах окружающей среды: Дисс. … канд. техн. наук. Краснодар, 2005. 173 с.
32. Assaf R., Levin I., Bravdo B. The effect of soil water distribution in drip irrigation of apple trees on their growth and yield // Proc. International Round-table Conference on microirrigation. Budapest, 1986. Vol. 1, pp. 19–25.
33. Assouline S. The Effects of Microdrip and Conventional Drip Irrigation on Water Distribution and Uptake // Soil Science Society of America Journal. 2002. Vol. 66. No. 5. pp. 1630–1636.
34. Zebe J.A., Serna-Perez A. Partial rootzone drying to save water while growing apples in a semi-arid region // Irrigation and Drainage. Managing water for sustainable agriculture. 2012. Vol. 61. No. 2. pp. 251–259.
Zebe J.A., Serna-Perez A. Partial rootzone drying to save water while growing apples in a semi-arid region // Irrigation and Drainage. Managing water for sustainable agriculture. 2012. Vol. 61. No. 2. pp. 251–259.
Анализ изученности кроны и насаждений яблони
Достижения биологии, физиологии сельскохозяйственных растений, комплексный подход к исследованию продуктивности естественных и искусственных фитоценозов обусловили в последние 60 лет очень существенный рост урожайности посевов и насаждений с/х культур.
Появилось много сортов и гибридов, которые в соответствующих условиях дают значительный прирост урожайности.
Плодоводство
В плодоводстве получили широкое распространение слаборослые подвои, малообъемные формировки крон и загущенные насаждения.
Такие типы садов объективно обеспечивают более эффективное использование основного нерегулируемого в полевых условиях ресурса – прихода фотосинтетически активной радиации солнца.
Проводятся интенсивные испытания разных систем формирования и обрезки кроны, технологий возделывания разных плодовых пород и сортов в различных климатических и почвенных условиях.
Организован мониторинг плодовых насаждений в разных почвенно-климатических зонах Украины, исследуются адаптации садов к неблагоприятным факторам среды обитания.
При подборе сортов для выращивания в конкретных условиях целесообразно знать их потенциальную продуктивность в данных условиях.
При этом в комплексе традиционных параметров, которые используются плодоводами при сравнительном изучении сортов (сила роста, габитус кроны, характер плодоношения, устойчивость к неблагоприятным факторам).
Целесообразно ввести показатели хозяйственной продуктивности фотосинтеза и фотосинтетический потенциал. Эти показатели дают возможность анализировать, насколько полно может исследуемый сорт использовать потенциал почвенного плодородия, тепловые и световые ресурсы местности и, на этой основе, проводить более рациональные исследования и подбор сортов.
В данной работе сделана первая попытка провести такой анализ трех сортов яблони в условиях предгорной зоны Крыма.
Обзор литературы
К.А. Тимирязев
В результате исследований роли света в фотосинтезе установил «космическую роль» зеленых растений на Земле.
Растения поглощают поступающий из космоса на Землю солнечный свет, трансформируют световую энергию в химическую энергию продуктов фотосинтеза – органических веществ, которые являются пищей для всех животных и человека.
Как следствие это функционального открытия, К.А. Тимирязев поставил задачу более эффективного использования поступающей на Землю от Солнца световой энергии.
«…луч солнца, упавший не на зеленый лист возделываемого поля, а на камень или голую землю – это кусок хлеба, безвозвратно потерянный для человечества.»
Опыт целенаправленного выращивания человеком растений для получения урожая плодов накапливался несколько тысячелетий, но только в последние два столетия создаются научные основы этой деятельности.
И.В. Мичурин
В плодоводстве устанавливается значение сорта и подвоя, и получают практическую значимость тезисы И.В. Мичурина «Сорт решает успех дела», «Подвой – это фундамент плодового дерева».
Проводятся исследования биологии плодовых деревьев и устанавливаются основные закономерности естественного формирования и развития надземной части, корневых систем.
Разрабатываются основы интенсификации производства за счет введения и использования слаборослых подвоев, ухода за корневыми системами через орошение, удобрение и комплексов рациональных приемов ее обработки и содержания в междурядьях сада.
Ранее мы писали про Хранение семян
Разрабатываются системы естественно-улучшенных формировок.. Особенно важна в интенсификации плодоводства роль использования искусственных формировок крон в сочетании с использованием слаборослых подвоев и сортов с комплексом ценных свойств.
При этом в качестве теоретической основы применения в саду искусственных формировок малообъемных крон используется тезис: обеспечить достаточно высокую освещенность листьев внутри кроны при загущенном расположении деревьев в ряду, т.е. создавать «крону-ряд».
Такой подход к конструировании сада основан на сформулированном К.А. Тимирязевым тезисе о необходимости рационального использования человеком солнечного света.
Системный подход
Исторически сложилось так, ч то только в 50-х годах прошлого столетия ученые-растениеводы начали применять системный подход к исследованию посевов и насаждений как продуктивных фотосинтезирующих систем.
Были разработаны приборы и методики исследований фотосинтеза и светового режима агрофитоценозов, установлены принципы оценки энергетической эффективности посевов и насаждений.
Принята и осуществлена международная биологическая программа исследований фотосинтеза на разных уровнях, в том числе – уровне агрофитоценозов, проводятся исследования по новой международной программе под эгидой ООН.
Рекомендуем прочитать: Строение семян
В результате этих исследований были установлены основные критерии оценки агрофитоценозов как био-физиологических систем:
- индекса облиствленности (индекса листовой поверхности) посевов разных культур,
- фотосинтетического потенциала агрофитоценозов,
- характера распределения фотосинтетически активной радиации в зависимости от структуры посевов основных полевых культур.
Установлены механизмы влияния режимов водного и минерального питания на урожайность агрофитоценозов через изменения фотосинтетического потенциала, интенсивности фотосинтеза, его продуктивность и хозяйственный коэффициент.
Концепция
Разработана концепция создания комплексов последовательных агротехнических мероприятий и приемов по формированию агрофитоценозов с конкретными характеристиками продукционной фотосинтезирующей системы.
Которые практически гарантируют получение запланированного количества и качества урожая. На основе этой концепции осуществляется программирование урожайности.
Которое предусматривает мониторинг состояния агроценоза как фотосинтезирующей системы и своевременную коррекцию процессов роста и формирования урожая через изменение водного и минерального питания или воздействия непосредственно на растение.
Большая часть таких исследований и разработок проведена для посевов полевых культур и овощных растений в условиях теплиц.
Плодовые сады как агрофитоценозы своей структурой и биологическими особенностями плодоношения очень сильно отличаются от полевых агрофитоценозов.
Ещё по теме: Размножение луковицами
По этой причине нельзя механически применять результаты исследований фитоценотической деятельности и урожая в посевах как рекомендации для критериев формирования структуры сада как фотосинтезирующей системы.
Для исследования динамики формирования фотосинтетического потенциала и продуктивности фотосинтеза плодовых деревьев нельзя использовать методические подходы, используемые для этих целей в полевых агроценозах, т.к. они основаны на удалении с поля растений учетных площадок.
По этой причине насаждения плодовых деревьев с разными конструкциями, размерами крон, системами их формирования и обрезки, схемами размещения в саду исследованы как продукционные фотосинтезирующие системы недостаточною
Если учесть большое разнообразие биологических особенностей роста, габитуса кроны и плодоношения разных плодовых пород и сортов.
Исследования
Около пятидесяти лет в Крыму проводились исследования характера распространения суммарной и фотосинтетически активной радиации в кронах яблони, активности и хозяйственной продуктивности фотосинтеза листьев в разных зонах кроны.
Этими исследованиями в общем подтверждены данные, полученные в подобных исследованиях, проведенных в других климатических зонах.
Установлено, что продуктивность фотосинтеза различна у разных сортов яблони и прямо зависит от среднедневной интенсивности ФАР и интенсивности фотосинтеза в соответствующей зоне кроны.
В результате исследований светового режима кроны в зависимости интенсивности фотосинтеза от освещенности листьев получили теоретическое обоснование практические данные о более высокой урожайности загущенных садов с плоскими и малообъемными кронами.
Во всех встреченных в доступной литературе рекомендациях по формированию крон в качестве основного положения указывается, что зона достаточного освещения листьев простирается до 1,5 м в глубину кроны от ее периферийных листьев.
В работе отмечается, что при очень сильном загущении кроны зона недостаточного освещения листьев может находиться уже на расстоянии 0,5 м от периферийных листьев. В этой же работе отмечено, что плотность листового полога зависит от формировки кроны, ее обрезки и свойств сорта.
Из этого следует важный : рекомендация создавать «плодовую стену» или «крону-ряд» шириной до 2,5 – 3,0 метров не всегда оправдана, нужно учитывать особенности роста, характера ветвления и плодоношения конкретного сорта.
А.А.Ничипорович
Работами А.А.Ничипоровича и других исследователей агрофитоценозов как продукционных фотосинтезирующих систем установлено.
Что урожайность агрофитоценоза зависит прямо пропорционально от хозяйственной продуктивности фотосинтеза и фотосинтетического потенциала: Урожайность (сухого в-ва) = ПФ хоз Х ФП.
Хозяйственная продуктивность фотосинтеза яблони – это суточный прирост сухого вещества яблок в расчете на 1 м2 листьев. ПФ хоз прямо зависит от обеспеченности растений водой и освещенности листьев, условий минерального питания растений.
На хозяйственный коэффициент фотосинтеза яблони в саду может очень сильно влиять обрезка кроны, а также биологические особенности сорта и подвоя.
Учитывая разную адаптированность сортов яблони к температурному и водному режиму атмосферы в период вегетации, можно предположить.
Что средняя за день интенсивность фотосинтеза и общее накопление (прирост) сухого вещества биомассы в расчете на 1м2 листьев за сутки у разных сортов также могут быть разными в одинаковых условиях.
В доступной литературе нет сведений о чистой продуктивности фотосинтеза яблони, а сведения о хозяйственной продуктивности фотосинтеза встречаются очень редко.
Это обусловлено большими методическими трудностями в отличие от полевых культур, имеющих относительно небольшие размеры растений, у плодоносящих деревьев яблони практически невозможно провести прямые учеты годичного прироста биомассы всего дерева.
Для таких растений применимы методы расчета прироста биомассы по учетам объемов древесины и массы листьев и плодов.
Фотосинтез
Значительно проще проводить расчеты хозяйственной продуктивности фотосинтеза через учеты массы урожая с одного дерева и площади листьев в его кроне.
Однако такой подход к решению задачи возможен практически только в садах с малообъемными кронами, т.к. площадь листьев в кроне всего дерева определить очень трудно.
Хозяйственную продуктивность фотосинтеза определяют кольцеванием плодоносных веточек (кольчаток¸копьец, плодовых прутиков). При этом отток продуктов фотосинтеза из листьев, расположенных на такой веточке, прерывают удалением неширокого кольца коры.
В результате такой операции практически все продукты фотосинтеза листьев, расположенных выше снятого кольца коры, расходуются на рост плода.
Прирост сухого вещества плода за время опыта учитывается по разнице массы в начале (в день окольцевания веточки) и в конце (при созревании плода) определения.
Для определения содержания сухого вещества в плоде в начале опыта подбирают точно таких же размеров плод этого же сорта. По данным этот метод можно использовать и для определения чистой продуктивности фотосинтеза яблони.
Кольцевание
Для этого, кроме прироста сухого вещества плода, нужно учесть также прирост сухого вещества окольцованной веточки.
При кольцевании веточки с плодами нужно учитывать, что о суммарной площади листьев, которые будут питать плод на окольцованной ветке, может зависеть продуктивность их фотосинтеза.
Необходимо также помнить, что в разных по освещенности листьев зонах кроны продуктивность фотосинтеза может различаться очень сильно. Эти сведения говорят о несовершенстве метода, т.к. он требует учета большого количества сопутствующих факторов.
Несмотря на методические трудности, полная оценка потенциальной продуктивности сорта в конкретных условиях выращивания невозможна без таких исследований.
Это заключение основывается на недостаточном обосновании практических рекомендаций расчета удельной облиствленности завязей при прореживании плодов в кронах яблони на разных подвоях, суммарной площади листьев для получения запланированного урожая с 1 дерева и 1 га сада.
Читайте также: Размножение черенками
Например, проф. А.П.Драгавцев приводит сведения о том, что при нагрузке яблонь урожаем 1,0 – 1,5 кг плодов на 1 м2 листьев в кроне обеспечивает регулярное плодоношение, а 2,0 кг и более на 1 м2 листьев вызывает периодичность плодоношения.
Мержаниан Ю.А. приводят результаты расчетов удельной продуктивности листьев в разных зонах кроны яблони. Он изменяется от 1,0 – 0,3 кг внутри кроны до 0,8 – 1,3 кг яблок на 1 м2 листьев в других зонах кроны.
При этом автор отмечает, что при устойчивой обеспеченности растений водой более продуктивны листья деревьев на слаборослом и среднерослом подвое по сравнению с сильнорослыми.
Но при недостатке орошения и засушливом лете у деревьев на слаборослом подвое удельная продуктивность листьев снижается сильнее, чем на сильнорослых.
Автор не приводит сведений продуктивности листьев разных сортов яблони, а этот вопрос имеет очень большое значение для подбора сортов в конкретных климатических условиях. В работе Н.И.Копылова отмечаются различия продуктивности фотосинтеза трех сортов яблони.
Местные условия
Учитывая разную адаптированность сортов к местным условиям, исследование хозяйственной продуктивности фотосинтеза как показателя адаптированности сорта может иметь большое значение.
Посевы однолетних полевых культур, насаждения плодовых деревьев – все агрофитоценозы – создаются для получения урожая, чтобы удовлетворить потребности для питания с/х животных и человека, решать экономические задачи.
Урожай (хозяйственная часть биомассы растений) формируется из продуктов фотосинтеза и интегральным показателем эффективности фотосинтеза является его хозяйственная продуктивность.
В процессе фотосинтеза происходит трансформация световой энергии солнечного излучения в химическую энергию органических соединений, которые синтезируются из СО2 и Н2О.
Фотосинтез зависит от многих внешних условий и внутренних факторов растений в агроценозе, но, в конечном итоге, все эти условия и факторы не изменяют главного условия – если световая энергия в достаточной мощности не поглощается листьями, то фотосинтез невозможен.
Поэтому, с точки зрения физики, агрофитоценозы нужно рассматривать как «ловушки» световой энергии, поступающей на Землю от Солнца и «устройства», которое эту «пойманную», т.е. поглощенную хлоропластами энергию «консервируют» и накапливают в биомассе.
Такой подход к оценке агрофитоценозов дает основной критерий – энергетическую эффективность посева, насаждения.
Этот критерий показывает, какая часть фотосинтетически активной радиации солнца, поступающей на всю площадь агрофитоценоза, использована растениями и накопилась в виде химической энергии в урожае с этой площади.
Эффективность
Очевидно, что чем выше урожайность, тем выше и энергетическая эффективность. Чтобы повысит энергетическую эффективность агрофитоценоза, необходимо обеспечить максимально возможное в конкретном агроценозе поглощение ФАР в течение вегетации.
Основной путь решения этой задачи – обеспечить достаточную облиствленность агрофитоценоза за весть этот период.
Как уже отмечалось, эффективность фотосинтетической деятельности листьев в плодовом саду как агрофитоценозе характеризуется хозяйственной продуктивностью фотосинтеза.
Хозяйственная продуктивность показывает, сколько граммов сухого вещества плодов накапливается за каждые сутки периода формирования урожая одним м2 листьев.
Урожайность, т.е. масса всех плодов на 1 га сада, формируется на этой площади не одни сутки, а в течение всего периода от цветения до созревания плодов, и не 1 м2 листьев, а всей площадью листьев, которая «работает» на 1 га сада в течение периода формирования урожая.
Показатель, который дает возможность оценить агрофитоценоз как фотосинтезирующую систему, способную обеспечить формирование определенной урожайности при известной продуктивности фотосинтеза, называют фотосинтетическим потенциалом.
Величину фотосинтетического потенциала 1 га фитоценоза математически можно выразить как произведение среднесуточной за время формирования урожая суммарной площади листьев на 1 га (S ср (м2)) и количества суток в периоде формирования урожая (n сут): ФП = S ср, м2 Х n сут.
Урожай
Установлено, что для получения удовлетворительных урожаев при средних значениях продуктивности фотосинтеза фотосинтетический потенциал полевых культур должен быть не менее 1 млн м2 * суток.
Для садовых агроценозов в доступной литературе сведения о фотосинтетическом потенциале садов разного типа не найдено. Учитывая принципиально важное значение фотосинтетического потенциала сада в его урожайности, целесообразно дать краткий анализ возможностей увеличения ЫАР сада.
ФП рассчитывается как произведение двух сомножителей: средней площади листьев и количества суток формирования.
Поскольку длительность периода формирования урожая является сортовым признаком и практически очень мало зависит от агротехники, основным и практическим единственным способом увеличить ФП сада является принятие мер по увеличению суммарной площади листьев в саду.
Облиствленность
Т.е. повышение облиствленности сада. Облиствленность сада можно охарактеризовать несколькими показателями: суммарной площадью листьев в кроне одного дерева или 1 га сада.
А также индексом облиствленности (или индексом листовой поверхности – ИЛП), т.е. соотношением суммарной площади листьев с площадью поверхности почвы, над которой эти листья расположены.
Для полевых культур сплошного сева ИЛП считается оптимальным при значениях 5 – 6, т.е. на 1 га посева размещается 5 – 6 га суммарной площади листьев. Для насаждений плодовых деревьев часто используются рекомендации придерживаться таких же величин ИЛП.
Однако структура садового агроценоза принципиально отличается от структуры посева полевых культур. В саду любой конструкции значительная часть поверхности почвы (междурядья сада) остается незанятой деревьями.
Поэтому ИЛП сада целесообразно рассчитывать как соотношение суммарной площади листьев кроны и площади проекции этой кроны на поверхность почвы. При таком расчете ИЛП сада должен быть значительно более высоким, чем ИЛП поля.
При этом очень важно, чтобы крона не была загущена. Структура кроны зависит от сортовых особенностей дерева, но может быть создана и поддерживаться в заданных параметрах обрезкой и формированием.
Ранее мы писали: Стратификация семян
Динамика нарастания площади листьев в начале вегетации зависит от характера ветвления (у сортов с преобладанием кольчаток облиствленность нарастает быстрее) и обрезки (укорачивание годичных приростов задерживает нарастание облствленности кроны).
В имеющейся литературе очень мало сведений о сравнительных характеристиках формирования облиствленности и фотосинтетическом потенциале разных сортов в насаждениях различной конструкции крон плодовых деревьев, поэтому исследования в этом направлении и могут быть актуальными.
Анализ изученности кроны и насаждений яблони как продукционной фотосинтезирующей системы показывает, что в существующих садах при урожайности 30 – 40 т плодов с 1 га потенциал сада как физиологической системы реализуется далеко не полностью.
Часто этот потенциал садов в конкретных условиях просто неизвестен. По этой причине целесообразно проводить исследования основных показателей сорта, от которых зависит урожайность сада как результат функционирования его как продукционной фотосинтезирующей системы.
Зная эти показатели, можно судить а потенциале урожайности этого сорта в конкретных условиях и принимать целенаправленные меры по реализации этого потенциала через корректировку в системе мер по созданию сада и ухода за ним.
ВЛИЯНИЕ ЛИСТОВЫХ ПОДКОРМОК НА ПРОДУКТИВНОСТЬ И КАЧЕСТВО ПЛОДОВ ЯБЛОНИ НА ПОДВОЕ СК4 | Попова
1. Ryndin A., Belous O., AbilfazovaY. [et al.]. The regulation of the functional state of subtropical crops with micronutrients, Potravinarstvo. 2017;11(1):175-182. DOI: https://dx.doi.org/10.5219/669
2. Сергеева Н.Н. Оптимизация минерального питания плодовых культур на юге России. В сборнике: Садоводство и виноградарство 21 века, Краснодар, 1999, 204-205.
3. Сергеева Н.Н., Киселева Г.К., Караваева А.В. Влияние некорневых подкормок на содержание фотосинтетических пигментов в листьях яблони, Плодоводство и виноградарство юга России. 2019,55(1):82-94. DOI: 10.3079/2219-5335-2019-1-55-82-94.
4. Popova V.P., Yaroshenko O.V., Sergeeva N.N. The Effect of foliar feeding on physiological condition of apple trees and chemical content of fruits, Potravinarstvo. 2018;12(1):634-643. DOI:https://doi.org/10.5219/928.
5. Дорошенко Т.Н., Чумаков С.С., Максимцов Д.В. Особенности некорневого питания плодовых растений при действии температурных стресс-факторов весенне-летнего периода, Плодоводство и ягодоводство России, 2012;30:22–30.
Дорошенко Т.Н., Чумаков С.С., Максимцов Д.В. Особенности некорневого питания плодовых растений при действии температурных стресс-факторов весенне-летнего периода, Плодоводство и ягодоводство России, 2012;30:22–30.
6. Дорошенко Т. Н., Чумаков С.С., Максимцов Д.В. [и др.]. Влияние некорневых подкормок на процессы жизнедеятельности плодовых растений при действии температурных стресс-факторов. Виды и уровни воздействия стресс-факторов среды на устойчивость агроэкосистем в условиях изменения климата, Краснодар, 2012. URL: http://www.kubansad.ru/ru/ www.kubansad.
7. Ненько Н.И., Сергеева Н.Н., Караваева А.В. Влияние листовых подкормок на адаптацию растений яблони к стрессовым факторам летнего периода. В сборнике: Последние научные достижения, 2012:14-18.
8. Трунов Ю.В., Цуканова Е.Н. Ткачёв О.А [и др.]. Активизация адаптационных механизмов растений яблони под влиянием специальных удобрений, Плодоводство и виноградарство юга России, 2011;12(6). URL: http://journal.kubansad.ru/archive/13/
9. Buntsevich L., Sergeeva N. Morphophysiological effects of various foliar nutrition regimes in apple in the south of Russia, Universal Jornal of Plant Scence. 2014;2(3):63-68.URL: http://www.hrpub.org/.
10. Ненько Н.И., Сергеева Н.Н., Караваева А.В. Исследование адаптивных реакций сортов яблони на фоне листовых обработок специальными удобрениями и регуляторами роста, Плодоводство и виноградарство Юга России 2015;35(05). URL: http://journal.kubansad.ru/pdf/15/05/07.pdf.
11. Попова В.П., Ярошенко О.В., Сергеева Н.Н., Ненько Н. И. Рациональная система применения специальных удобрений на яблоне, Научные труды Северо-Кавказского федерального научного центра садоводства, виноградарства, виноделия. 2018;17:51-56. DOI: 10.30679/2587-9847-2018-17-51-56.
И. Рациональная система применения специальных удобрений на яблоне, Научные труды Северо-Кавказского федерального научного центра садоводства, виноградарства, виноделия. 2018;17:51-56. DOI: 10.30679/2587-9847-2018-17-51-56.
12. Nenko N., Sergeeva N., Kiseleva G., Karavaeva A. Effect of growth regulators and mineral foliar application apple on stability fruit cenosis, News of science and education. 2015;5(29):53-61. URL: https://elibrary.ru/item.asp?id=24556639.
13. Методические указания по проведению исследований в длительных опытах с удобрениями. Ч. III. М.: ВАСХНИЛ, 1976, 135 с.
14. Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур. Орёл: ВНИИСПК, 1999. 608 с.
15. Гавриленко В.Ф., Ладыгина М.Е., Хандобина Л.М. Большой практикум по физиологии растений. М.: Высшая школа, 1975. 380 с.
16. Волков Ф.А. Методика исследований в садоводстве. М.: ВСТИСП, 2005. 150 с.
Влияние листовых подкормок на продуктивность и качество плодов яблони в условиях Краснодарского края | Попова
1. Ryndin A., Belous O., Abilfazova Y. et al. The regulation of the functional state of subtropical crops with micronutrients, Potravinarstvo. 2017;11(1):175-182. DOI: https://dx.doi.org/10.5219/669
2. Сергеева Н. Н. Оптимизация минерального питания плодовых культур на юге России. В сборнике: Садоводство и виноградарство 21 века, Краснодар, 1999, 204-205.
3. Сергеева Н. Н., Киселева Г. К., Караваева А. В. Влияние некорневых подкормок на содержание фотосинтетических пигментов в листьях яблони, Плодоводство и виноградарство юга России. 2019;55(1):82-94. DOI: 10.3079/2219-5335-2019-1-55-82-94.
4. Srinivasa Rao N. K., Laxman R. H., Shivashankara K. S. Physiological and Morphological Responses of Horticultural Crops to Abiotic Stresses. Abiotic Stress Physiology of Horticultural Crops. 2016:3-17. DOI:https://doi.org/10.1007/978-81-322-2725-0_1
5. Abilphazova Y., Belous O. Adaptability of grades and hybrids of tangerine in a subtropical zone of Russia. Potravinarstvo. 2015;9(1):299-303. DOI: https://doi.org/10.5219/485.
6. Kuzin A. I., Trunov Yu. V., Solovyev A. V. Effect of fertigation on yield and fruit quality of apple (Malus domesticaBorkh.) in high-density orchards on chernozems in Central Russia. Acta Horticulturae. 2018;1217:343-350. DOI:https://doi.org/10.17660/ActaHortic.2018.1217.43
7. Popova V. P., Yaroshenko O. V., Sergeeva N. N. The Effect of foliar feeding on physiological condition of apple trees and chemical content of fruits, Potravinarstvo. 2018;12(1):634-643. DOI:https://doi.org/10.5219/928.
8. Дорошенко Т. Н., Чумаков С. С., Максимцов Д. В. Особенности некорневого питания плодовых растений при действии температурных стресс-факторов весенне-летнего периода, Плодоводство и ягодоводство России, 2012;30:22-30.
9. Дорошенко Т. Н., Чумаков С. С., Максимцов Д. В. и др. Влияние некорневых подкормок на процессы жизнедеятельности плодовых растений при действии температурных стресс-факторов. Виды и уровни воздействия стресс-факторов среды на устойчивость агроэкосистем в условиях изменения климата, Краснодар, 2012. URL: http://www.kubansad.ru/ru/ www.kubansad. (ссылка активна на 25.05.2019).
и др. Влияние некорневых подкормок на процессы жизнедеятельности плодовых растений при действии температурных стресс-факторов. Виды и уровни воздействия стресс-факторов среды на устойчивость агроэкосистем в условиях изменения климата, Краснодар, 2012. URL: http://www.kubansad.ru/ru/ www.kubansad. (ссылка активна на 25.05.2019).
10. Ненько Н. И., Сергеева Н. Н., Караваева А. В. Влияние листовых подкормок на адаптацию растений яблони к стрессовым факторам летнего периода. Последние научные достижения, 2012:14-18.
11. Трунов Ю. В., Цуканова Е. Н., Ткачёв О. А. и др. Активизация адаптационных механизмов растений яблони под влиянием специальных удобрений, Плодоводство и виноградарство юга России. 2011;12(6):78-89. URL: http://journal.kubansad.ru/archive/13/ (ссылка активна на 25.05.2019).
12. Buntsevich L., Sergeeva N. Morphophysiological effects of various foliar nutrition regimes in apple in the south of Russia, Universal Journal of Plant Science. 2014;2(3):63-68. URL: http://www.hrpub.org/. (link activeon 05/25/2019)
13. Ненько Н. И., Сергеева Н. Н., Караваева А. В. Исследование адаптивных реакций сортов яблони на фоне листовых обработок специальными удобрениями и регуляторами роста, Плодоводство и виноградарство Юга России. 2015;35(05):83-94. URL: http://journal.kubansad.ru/pdf/15/05/07.pdf. (ссылка активна на 25.05.2019)
14. Попова В. П., Ярошенко О. В., Сергеева Н. Н., Ненько Н. И. Рациональная система применения специальных удобрений на яблоне, Научные труды Северо-Кавказского федерального научного центра садоводства, виноградарства, виноделия. 2018;17:51-56. DOI: 10.30679/2587-9847-2018-17-51-56.
15. Nenko N., Sergeeva N., Kiseleva G., Karavaeva A. Effect of growth regulators and mineral foliar application apple on stability of fruit cenosis, News of science and education. 2015;5(29):53-61.
16. Методические указания по проведению исследований в длительных опытах с удобрениями. Ч. III. М.: ВАСХНИЛ, 1976, 135 с.
17. Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур. Орёл: ВНИИСПК, 1999, 608 с.
18. Гавриленко В. Ф., Ладыгина М. Е., Хандобина Л. М. Большой практикум по физиологии растений. М.: Высшая школа, 1975. 380 с.
19. Волков Ф. А. Методика исследований в садоводстве. М.: ВСТИСП, 2005, 150 с.
Индекс площади листьев под кроной яблони с использованием полусферической фотографии в засушливых регионах
Основные моменты
- •
Оценка LAI методом логарифмического эллипса была близка к прямой LAI.
- •
Оценка LAI по полусферическим изображениям с использованием метода логарифмического эллипса была точной даже без калибровки индекса слипания.
- •
Полусферическая фотография может обеспечить надежные параметры купола для модели эвапотранспирации.
Abstract
Полусферические изображения яблоневого сада были получены с помощью объектива «рыбий глаз» в 2010 году, а затем соответственно проанализированы методами Bonhom, 2000, 2000G, сфера, эллипс, кольца LAI 2000-3, кольца LAI 2000-4 и Миллера, чтобы получить индекс площади листьев (LAI), индекс площади растений (PAI), долю промежутков, угол наклона листьев и индекс комкования в засушливом регионе северо-запада Китая. Результаты показывают, что во время DOY (день года) 202–237 (на стадии расцвета) оценка LAI методом логарифмического эллипса и PAI методом колец LAI 2000-4 составляла 1.96 и 1,95 м 2 м −2 соответственно, что близко к прямому LAI (1.85 м 2 м −2 ). Однако прямой LAI был выше, чем оцененный LAI по методу эллипса Hemiview (1,59 м 2 м -2 ) и ниже, чем оцененный PAI по методу анализа фракции зазора в Photoshop (2,67 м 2 м — 2 ). Таким образом, методы логарифмического эллипса и колец LAI 2000-4 были использованы для оценки LAI (PAI) по полусферическим изображениям.И оцененный LAI методами логарифмического эллипса и колец LAI 2000-4 использовался для анализа сезонных колебаний параметров растительного покрова. LAI быстро увеличивался на стадии распускания листьев и затем поддерживался на высоком уровне от стадии роста до сбора урожая. Доля зазора купола на южной стороне была аналогична таковой на северной стороне. Угол наклона листа по горизонтали имел тенденцию получать больше излучения по мере развития яблони. Кроме того, оценка LAI из полусферических изображений с использованием методов логарифмического эллипса и колец LAI 2000-4 была точной даже без калибровки индекса слипания.Таким образом, оцененные параметры полога из полусферических изображений с использованием методов логарифмического эллипса и колец LAI 2000-4 могут быть использованы для моделирования эвапотранспирации.
Результаты показывают, что во время DOY (день года) 202–237 (на стадии расцвета) оценка LAI методом логарифмического эллипса и PAI методом колец LAI 2000-4 составляла 1.96 и 1,95 м 2 м −2 соответственно, что близко к прямому LAI (1.85 м 2 м −2 ). Однако прямой LAI был выше, чем оцененный LAI по методу эллипса Hemiview (1,59 м 2 м -2 ) и ниже, чем оцененный PAI по методу анализа фракции зазора в Photoshop (2,67 м 2 м — 2 ). Таким образом, методы логарифмического эллипса и колец LAI 2000-4 были использованы для оценки LAI (PAI) по полусферическим изображениям.И оцененный LAI методами логарифмического эллипса и колец LAI 2000-4 использовался для анализа сезонных колебаний параметров растительного покрова. LAI быстро увеличивался на стадии распускания листьев и затем поддерживался на высоком уровне от стадии роста до сбора урожая. Доля зазора купола на южной стороне была аналогична таковой на северной стороне. Угол наклона листа по горизонтали имел тенденцию получать больше излучения по мере развития яблони. Кроме того, оценка LAI из полусферических изображений с использованием методов логарифмического эллипса и колец LAI 2000-4 была точной даже без калибровки индекса слипания.Таким образом, оцененные параметры полога из полусферических изображений с использованием методов логарифмического эллипса и колец LAI 2000-4 могут быть использованы для моделирования эвапотранспирации.
Ключевые слова
Программное обеспечение для анализа
Яблоневый сад
Структура навеса
Доля зазора
Полусферическое изображение
Индекс площади листа
Рекомендуемые статьи Цитирующие статьи (0)
Полный текст
Рекомендуемые статьи
Цитирующие статьи
Площадь фотосинтетической поверхности яблонь
- Кшиштоф Токарц
- Мацей Кочурек
 org/Person» itemprop=»author»> Ян Пиларски
org/Person» itemprop=»author»> Ян ПиларскиДоклад конференции
5
Упоминания- 1.5к
Загрузки
Часть
Продвинутые темы науки и техники в Китае
Книжная серия (ATSTC)
Реферат
Развитие площадей поверхности стебля и листьев было изучено сравнительно в течение годового цикла, и были рассчитаны доли этих площадей в общей площади деревьев. В зависимости от сорта общая площадь поверхности деревьев: всех ветвей, сучьев и стволов деревьев после зимней обрезки составляла от 27 дм 2 до 118 дм 2 .В начале вегетационного периода доля листьев во вновь развившихся стеблях составляла 90–92%, а доля этих новых стеблей составляла 8–10%. В более поздний период вегетации листовая площадь составляла 83–89%, а площадь новых стеблей — 11–17%, и различий между сортами не было. К концу вегетационного периода общая площадь листьев на дереве составляла от 110–120 дм 2 до 296 дм 2 , а площадь вновь выросших стеблей — от 42 дм 2 до 168 дм 2 .
В течение вегетационного периода доля площади стебля уменьшалась, а доля листьев увеличивалась, и с августа площадь листьев превышала площадь стеблей у большинства изученных сортов. В заключительной части вегетационного периода листовая площадь превышала площадь стеблей более чем на 20–45%.
Ключевые слова
Фотосинтетическая площадь поверхности Стебли Лист
Это предварительный просмотр содержимого подписки,
войдите в систему
, чтобы проверить доступ.
Предварительный просмотр
Невозможно отобразить предварительный просмотр. Скачать превью PDF.
Ссылки
Aschan G, Pfanz H, Vodnik D, Batic F (2005) Фотосинтетические характеристики вегетативных и репродуктивных структур зеленого морозника (Helleborus Viridis L.
 Agg.) Photosynthetica 43: 55–64
Agg.) Photosynthetica 43: 55–64CrossRefGoogle Scholar
Бенцинг Д.Х., Фридман В.Е., Петерсон Дж., Ренфроу А. (1983) Бесплодие, пеламентные корни и преобладание орхидных в эпифитном биотопе.Являюсь. J. Bot. 70: 121–133
CrossRefGoogle Scholar
Bosc A, Grandcourt DA, Loustau D (2003) Изменчивость поддерживающего дыхания стебля и ветвей у соснового дерева Pinus Pinaster. Tree Physiol. 23: 227–236
PubMedCrossRefGoogle Scholar
Брейер Л., Экхардт К., Фреде Х.Г. (2003) Значения параметров растений для моделей в умеренном климате. Ecol Model 169: 237–293
CrossRefGoogle Scholar
Бланке М.М., Ленц Ф. (1989) Фотосинтез фруктов — обзор.Plant Cell Environ. 12: 31–46
CrossRefGoogle Scholar
Комсток Дж. П., Элерингер Дж. Р. (1988) Сезонные закономерности развития растительного покрова и накопления углерода у девятнадцати видов теплых пустынных кустарников. Oecologia 75: 327–335
CrossRefGoogle Scholar
Чен Дж. М., Рич П. М., Гауэр С. Т., Норман Дж. М., Пламмер С. (1997) Индекс площади листьев бореальных лесов: теория, методы и измерения. J Geophys Res 102: 29429–29443
CrossRefGoogle Scholar
Дима Э., Манетас Й., Псарас Г.К. (2006) Структура распределения хлорофилла во внутренних тканях стебля: данные эпифлуоресцентной микроскопии и измерения отражательной способности у 20 видов древесных растений.Деревья 20: 515–521
CrossRefGoogle Scholar
Достер Р.Х., Джеймс Д.А. (1998) Размер домашнего ареала и место обитания дятлов с красной кокеткой в горах Уашита в Арканзасе. Wilson Bull 110: 110–117
Google Scholar
Foote KC, Schaedle M (1976) Суточные и сезонные закономерности фотосинтеза и дыхания стеблями Populus Tremuloides Michx. Plant Physiol. 58: 651–655
PubMedCrossRefGoogle Scholar
Hew CS, NG Y, Wong SC, Yeoh HH, Ho KK (1984) Фиксация диоксида углерода в воздушных корнях орхидей.Physiol. Растение. 60: 154–158
CrossRefGoogle Scholar
Kaim RF (2004) Ослабление осадков лесными пологами. Кандидат наук. Диссертация, Университет штата Орегон: Corvallis, OR, pp. 150
Google Scholar
Kitaya Y, Yabuki K, Kiyota M, Tani A, Hirano T., Aiga I (2002) Газообмен и концентрация кислорода в пневматофорах и опорных корнях Четыре вида мангровых зарослей. Деревья 155–158
Google Scholar
Харук В.И., Миддлтон Е.М., Спенсер С.Л., Рок Б.Н., Уильямс Д.Л. (1995) Фотосинтез коры осины и его значение для дистанционного зондирования и оценки углеродного бюджета в бореальной экосистеме.Загрязнение воды, воздуха и почвы. 82: 483–497
CrossRefGoogle Scholar
Kocurek M (2007) Udział Pędów Roślin Zielnych: Reynutria Japinica i Helianthus Tuberosus w Fotosyntetycznym Wiązaniu CO
2
. (Участие стеблей травянистых растений: Reynutria Japonica и Helianthus Tuberosus в фотосинтетической ассимиляции CO
2
. Докторская диссертация, Институт физиологии растений PAS, Краков, Польша
Google Scholar
Lange OL, Zuber M (1977) Frerea Indica, стеблевое суккулентное растение САМ с листопадными C3-листьями.Oecologia 31: 67–72
CrossRefGoogle Scholar
Larcher W, Lutz C, Nagelle M, Bodner M (1988) Фотосинтетическое функционирование и ультраструктура хлоропластов в стволовых тканях Fagus Sylvatica. J. Plant Physiol. 132: 731–737
CrossRefGoogle Scholar
Nilsen ET (1995) Степень, закономерности и роль фотосинтеза ствола в углеродной экономике растений. В: Gartner B (ред.), Стебли растений — физиология и функциональная морфология. Academic Press: San Diego, pp. 223–240
Google Scholar
Нобель PS, Hartsock TL (1986) Leaf and Stem CO
2
Поглощение в трех подсемействах кактусов Physiol.80: 913–917
PubMedCrossRefGoogle Scholar
Пфанц Х., Ашан Г. (2001) Существование фотосинтеза коры и стебля и его значение для общего увеличения выбросов углерода. Эко-физиологический и экологический подход. Progress in Botany Vol. 62: 477–510
CrossRefGoogle Scholar
Pfanz H, Aschan G, Langenfeld-Heyser R, Wittman C, Loose M (2002) Экология и экофизиология кортикального слоя стволов деревьев и фотосинтеза древесины. Naturwissenchaften 89: 147–162
CrossRefGoogle Scholar
Pilarski J (1984) Содержание хлорофильных пигментов в коре побегов и листьях Syringa Vulgaris L.Bul. Pol. Акад. Sci. Биол. Sci. 32: 415–423
Google Scholar
Пиларски Дж. (1990) Фотохимическая активность изолированных хлоропластов коры и листьев сирени (Syringa Vulgaris L.). Photosynthetica 24: 186–189
Google Scholar
Пиларски Дж. (1997) Связь между солнечным облучением, температурой и фотохимической активностью хлоропластов, выделенных из коры и листьев сирени Syringa Vulgaris L.). Pol. J. Environ. Stud. 6: 53–57
Google Scholar
Пиларски Дж. (1999) Градиент фотосинтетических пигментов в коре и листьях сирени (Syringa Vulgaris L.) Acta Physiol. Растение. 21: 365–373
CrossRefGoogle Scholar
Пиларски Дж. (2002) Суточные и сезонные изменения интенсивности фотосинтеза в стебле сирени (Syringa Vulgaris L.). Acta Physiol. Растение. 24: 29–36
CrossRefGoogle Scholar
Пиларски Дж., Токарз К. (2005) Сравнение оптических свойств фруктовых деревьев: черешни, вишни, груши, сливы и грецкого ореха. Folia Horticult. 17: 89–101
Google Scholar
Schaedle M (1975) Tree Photosynthetsis.Annu Rev растительный физиол. 26: 101–115
CrossRefGoogle Scholar
Scurlock JMO, Asner GP, Gower ST (2001) Всемирные исторические оценки и библиография индекса площади листа, 1932–2000. Технический меморандум ORNL TM-2001/268, Национальная лаборатория Ок-Ридж, Ок-Ридж, Теннесси, США
Google Scholar
Смит С.Д., Осмонд С.Б. (1987) Фотосинтез стебля в эфемерных эфемерах пустыни, Eriogonum Inflatum: морфология, проводимость устья и вода -Эффективность использования в полевых условиях.Oecologia 72: 533–541
CrossRefGoogle Scholar
Solhaug KA, Haugen J (1998) Сезонное изменение фотоингибирования фотосинтеза в коре Populus Tremula L. Photosynthetica 35: 411–417
CrossRefGoogle Scholar
- Kilar Tokarski,
Pilar Tokarski. (2005) Оптические свойства и содержание фотосинтетических пигментов в стеблях и листьях яблони. Acta Physiol. Растение. 27: 183–191
CrossRefGoogle Scholar
Tokarz K (2007) Fotosyntetyczna Rola Pędów Jabłoni (Malus Domestica) и Ich Udział w Bilansie Wiazania CO
2
.Фотосинтетическая роль стеблей яблони (Malus Domestica) и их участие в фотосинтетической ассимиляции CO
2
. Диссертация, Институт физиологии растений PAS, Краков, Польша
Google Scholar
Weiskittel AR, Maguire AM (2006) Площадь поверхности ветвей и ее вертикальное распределение в прибрежной пихте Дугласа. Деревья 20: 657–667
CrossRefGoogle Scholar
Wittmann C, Pfanz H, Loreto F, Centritto M, Pietrini F, Alessio G (2006) Stem CO
2
Высвобождение при освещении: кортикальный фотосинтез, фотодыхание или ингибирование Митохондриальное дыхание.Растение, клетка и окружающая среда. 29: 1149–1158
CrossRefGoogle Scholar
Вайс Д., Шенфельд М., Халеви А. (1988) Фотосинтетическая активность в венчике петунии. Plant Physiol 87: 666–670
PubMedCrossRefGoogle Scholar
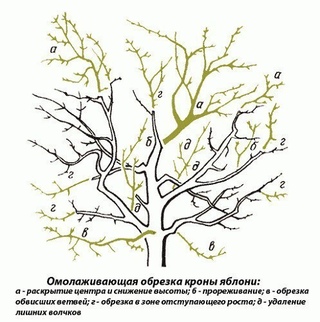 Agg.) Photosynthetica 43: 55–64
Agg.) Photosynthetica 43: 55–64Информация об авторских правах
© Zhejiang University Press, Ханчжоу и Springer-Verlag Berlin Heidelberg 2013
Авторы и аффилированные лица
- 1.Institute of Plant Physiology 2. Институт биологии Гуманитарно-научный университет Яна Кохановского Кельце Польша
Использование для защиты от парши яблони
Зависимость площади поверхности и количества листьев яблони от суммы температур: использование для защиты от парши яблони
| ||||||||||||
URL www.actahort.org Хостинг KU Leuven
© ISHS
(PDF) Цифровая фотография обложки для оценки индекса площади листа (LAI) яблонь с использованием переменного коэффициента ослабления света
Sensors 2015, 15 2871
4. TangF, H .; Dubayah, R .; Swatantran, A .; Хофтон, М.; Шелдон, С .; Clark, D.B .; Blair, B. Получение
вертикальных профилей LAI над тропическими дождевыми лесами с использованием лидара формы волны в Ла-Сельва, Коста-Рика.
Remote Sens. Environ. 2012, 124, 242–250.
5. Tang, H .; Dubayah, R .; Бролли, М .; Ganguly, S .; Zhang, G. Крупномасштабное извлечение индекса площади листьев
и вертикального профиля листвы с космического лидара сигналов (GLAS / ICESat). Remote Sens. Environ.
2014, 154, 8–18.
6. Вильялобос, Ф.J .; Оргаз, Ф .; Матеос, Л. Неразрушающее измерение площади листьев оливковых деревьев
(Olea europaea L.) с использованием метода инверсии промежутков. Agric. Для. Meteorol. 1995, 73, 29–42.
7. Jonckheere, I .; Fleck, S .; Nackaerts, K .; Муйс, Б .; Coppin, P .; Weiss, M .; Барет, Ф.Д.Р. Обзор
методов определения индекса площади листа на месте: Часть I. Теории, датчики и полусферическая фотография
. Agric. Для. Meteorol. 2004, 121, 19–35.
8. Ле Дантек, В.; Dufrene, E .; Saugier, B. Межгодовые и пространственные вариации максимальной площади листа
Индекс лиственных древостоев умеренного пояса. Для. Ecol. Manag. 2000, 134, 71–81.
9. Mencuccini, M .; Грейс, Дж. Климат влияет на соотношение площади листьев и площади заболони у сосны обыкновенной.
Tree Physiol. 1995, 15, 1–10.
10. Bréda, N.J.J. Наземные измерения индекса листовой поверхности: обзор методов, инструментов и текущих споров
. J. Exp. Бот. 2003, 54, 2403–2417.
11. Chen, J.M .; Цихлар, Дж. Теория анализа размера щелей в растительном покрове для улучшения оптических измерений индекса площади листа
. Прил. Опт. 1995, 34, 6211–6222.
12. Lang, A.R.G .; Макмертри, Р. Общая площадь листьев одиночных деревьев Eucalyptus grandis оценивается на основе
коэффициентов пропускания солнечного луча. Agric. Для. Meteorol. 1992, 58, 79–92.
13. Lang, A.R.G .; Юэцинь, X. Оценка индекса площади листа по пропусканию прямого солнечного света в
прерывистых пологах.Agric. Для. Meteorol. 1986, 37, 229–243.
14. Welles, J.M .; Коэн, С. Измерение структуры купола с помощью анализа фракции зазора с использованием коммерческого оборудования
. J. Exp. Бот. 1996, 47, 1335–1342.
15. Frazer, G.W .; Fournier, R.A .; Trofymow, J.A .; Холл, Р.Дж. Сравнение цифровой и пленочной фотографии «рыбий глаз»
для анализа структуры лесного полога и пропускания света в промежутках. Agric. Для. Meteorol.
2001, 109, 249–263.
16.Macfarlane, C .; Hoffman, M .; Eamus, D .; Kerp, N .; Higginson, S .; McMurtrie, R .; Адамс, М.
Оценка индекса площади листьев в эвкалиптовом лесу с использованием цифровой фотографии. Agric. Для. Meteorol.
2007, 143, 176–188.
17. Macfarlane, C .; Arndt, S.K .; Ливсли, С.Дж .; Эдгар, A.C .; White, D.A .; Adams, M.A .; Eamus, D.
Оценка индекса площади листьев в эвкалиптовом лесу с вертикальной листвой, с использованием укрытия и полнокадровой фотографии
«рыбий глаз». Для. Ecol.Manag. 2007, 242, 756–763.
18. Macfarlane, C .; Grigg, A .; Евангелиста, К. Оценка площади лесной листвы с использованием укрытия и полного кадра
Фотография «рыбий глаз»: мышление внутри круга. Agric. Для. Meteorol. 2007, 146, 1–12.
19. Fuentes, S .; Poblete-Echeverría, C .; Ортега-Фариас, С .; Tyerman, S .; de Bei, R. Автоматизированная оценка
индекса площади листьев по кронам виноградной лозы с использованием фото обложек, видео и компьютерных методов анализа
.Aust. J. Grape Wine Res. 2014, 20, 465–473.
20. Lindroth, A .; Пертту К. Простой расчет коэффициента исчезновения древостоев. Agric. Meteorol.
1981, 25, 97–110.
Сравнение плодовых и неплодоносящих деревьев в JSTOR
Abstract
Сезонные изменения фотосинтеза яблонь (Malus domestica Borkh.) Отслеживались для изучения влияния взаимодействий источник-поглотитель на фотосинтез и фотодыхание.Повышенная скорость фотосинтеза наблюдалась в течение двух периодов вегетации и коррелировала с процессом плодоношения. Первый период повышенной скорости фотосинтеза приходился на период цветения, когда шпорцевые листья на цветущих побегах демонстрировали до 25% более высокую скорость фотосинтеза, чем вегетативные шпорцевые листья на основе площади листа. Скорость ассимиляции CO2 была также выше у плодовых деревьев, чем у неплодоносящих деревьев в период быстрого роста плодов с июля по сентябрь. Фотодыхание, темное дыхание, сопротивление листьев и транспирация не показали сезонных изменений, которые коррелируют с наличием или отсутствием плодов.Эти данные представляют собой первое всестороннее исследование влияния цветения / образования плодов на фотосинтез и фотодыхание у многолетних растений.
Journal Information
Международный журнал «Физиология растений», основанный в 1926 году, посвящен физиологии, биохимии, клеточной и молекулярной биологии, генетике, биофизике и экологической биологии растений. Физиология растений — один из старейших и наиболее уважаемых журналов по науке о растениях.
Информация об издателе
Oxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные труды по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Морфометрия выявляет сложные и унаследованные формы листьев яблони
Реферат
Яблоко ( Malus spp .) — широко выращиваемая и ценная плодовая культура. Форма и размер листьев важны для цветения яблони, а также могут быть ранними индикаторами других ценных с точки зрения сельского хозяйства признаков. Мы исследовали 9000 листьев 869 уникальных образцов яблони, используя линейные измерения и комплексные морфометрические методы. Мы выявили аллометрические вариации отношения длины к ширине между образцами и видами яблони.Аллометрическая вариация была обусловлена вариацией ширины листовой пластинки, а не длины. Соотношение сторон сильно коррелировало с первичной осью морфометрической изменчивости (PC1), количественно определяемой с использованием эллиптических дескрипторов Фурье (EFD) и постоянной гомологии (PH). В то время как основным источником вариаций было соотношение сторон, последующие ПК соответствовали сложным вариациям формы, которые не фиксировались линейными измерениями. После связывания морфометрической информации с более чем 122 000 общегеномных SNP, мы обнаружили высокие значения наследуемости в узком смысле даже на более поздних ПК, что указывает на то, что комплексные морфометрические параметры могут фиксировать сложные наследуемые фенотипы.Таким образом, такие методы, как EFDs и PH, улавливают наследственные биологические вариации, которые можно было бы упустить с помощью одних только линейных измерений, и которые потенциально могут быть использованы для отбора скрытого фенотипа, обнаруживаемого только с использованием комплексных морфометрии.
Введение
Яблоки ( Malus spp .) — одна из наиболее широко выращиваемых фруктовых культур в мире, занимающая третье место в мире по объему производства более 84 миллионов тонн в 2014 году (1). Форма и размер листьев яблони играют важную роль в росте и развитии дерева и в конечном итоге влияют на характеристики плода.Листья яблони, как правило, простые, эллиптически-яйцевидные. Предыдущие исследования яблони использовали линейные измерения, такие как длина и ширина, для количественной оценки формы листа (2, 3). Соотношение длины к ширине является основным источником вариаций формы листьев. Различное соотношение сторон приводит к непропорциональному увеличению или уменьшению длины по сравнению с шириной или аллометрической изменчивостью листьев (4, 5). Хотя линейные измерения, такие как длина и ширина листа, полезны, они не могут полностью уловить разнообразие форм листа.Неспособность всесторонне измерить форму листа также ограничивает нашу способность распознавать общий генетический вклад.
Эллиптические дескрипторы Фурье (EFD) — ценный, хорошо известный инструмент для количественной оценки контура формы. Анализ EFD сначала преобразует контур в цепной код, метод сжатия данных без потерь, который кодирует форму с помощью цепочки чисел, в которой каждое число указывает пошаговые движения для восстановления пикселей, составляющих форму. Затем к цепному коду применяется разложение Фурье, определяющее форму как гармонический ряд.EFD широко использовались для количественной оценки формы листьев у различных видов, таких как виноград (6), томат (7) и Passiflora (8). В предыдущей работе для оценки формы плодов яблони использовались EFD (9), но этот метод еще не применялся к листьям яблони. Недавно разработанный морфометрический метод, постоянная гомология (PH), обеспечивает еще один метод оценки формы листа. PH, как и EFD, нормализован к разнице в размере, но также может быть инвариантным относительно ориентации. PH обрабатывает пиксели контура как двумерное облако точек перед применением оценки плотности соседей к каждому пикселю.Ряд ядер кольцевого зазора с увеличивающимся радиусом используется для выделения и сглаживания контурных плотностей. Количество соединенных компонентов записывается как функция плотности для каждого кольца, в результате чего получается кривая (сокращенная версия постоянного штрих-кода), которая количественно определяет форму как топологию. Подход PH на основе топологии также может быть применен к зубчатой и корневой архитектуре, что позволяет использовать аналогичную структуру в различных структурах предприятия (10, 11).
Всестороннее измерение формы листа с использованием таких подходов, как EFDs и PH, важно, поскольку особенности формы могут быть связаны с важными для сельского хозяйства признаками.У яблони листья присутствуют во время длительной ювенильной фазы, но плоды появляются только на зрелых деревьях, и, таким образом, признаки листьев могут способствовать раннему отбору без необходимости использования генетических маркеров. У яблони обычно требуется 5 лет для достижения значительного плодоношения, и любая способность выбросить деревья, не обладающие интересующим признаком на ранней стадии развития, чрезвычайно ценна (12). Уже есть несколько случаев, когда уникальные характеристики листьев служат ранним маркером других генетических различий яблони.Например, ген, лежащий в основе красного цвета мякоти плода, может приводить к накоплению антоцианов в листьях, вызывая красную листву (13, 14), в то время как столбчатая архитектура дерева может сопровождаться увеличением количества листьев, площади, веса на единицу площади и длины. отношение к ширине (15). PH листьев также был предложен в качестве раннего индикатора слабокислых плодов (16).
Помимо того, что они служат ранними маркерами для других признаков, форма и размер листьев могут влиять на количество света, получаемого деревом, а освещенность имеет решающее значение для цветения яблони.Проникновение света приводит к более высокому уровню цветения, в то время как повреждение листьев или дефолиация могут уменьшить цветение (17). Прореживание яблонь до определенного соотношения листьев и плодов — обычная практика для достижения оптимального цвета и размера плодов. Напротив, деревья с меньшим количеством плодов могут увеличить вегетативный рост и, следовательно, площадь листьев (18). В предыдущей работе несколько характеристик листьев, таких как площадь и периметр, коррелировали с размером плодов яблони (19). Ясно, что существует важная взаимосвязь между листьями и фруктами, и всесторонняя количественная оценка вариации формы листьев является решающим компонентом для понимания этой взаимосвязи у яблока.
Листья являются основными фотосинтетическими органами яблони, но генетическая основа, лежащая в основе их формы и размера, остается неизвестной. У хлопка один локус контролирует основные формы листа (20), но в большинстве случаев форма листа, по-видимому, контролируется множеством локусов с малым эффектом (5, 21). Существует ограниченное количество примеров геномного анализа формы листа у яблони, однако предыдущее исследование по картированию двух родительских сцеплений обнаружило два предполагающих локуса количественных признаков для размера листа (2). Предыдущая работа также измерила несколько характеристик листьев, таких как площадь, периметр и округлость, у 158 образцов яблони.Исследование связало эти измерения с 901 однонуклеотидным полиморфизмом (SNP), но не обнаружило значимых взаимосвязей генотип-фенотип (19). До сих пор не предпринимались попытки оценить генетическую наследственность комплексных морфометрических фенотипов листьев, таких как описанные с использованием EFD и PH. Поэтому остается неясным, в какой степени эти методы улавливают биологически значимые наследственные вариации.
Чтобы полностью понять генетическую основу формы листа, важно включать как линейные, так и морфометрические оценки формы.Снижение затрат на секвенирование и доступ к большой и разнообразной коллекции зародышевой плазмы позволили нам проанализировать около 9000 листьев из более чем 800 уникальных образцов, которые мы связали с более чем 122000 общегеномных SNP. Мы представляем первый всесторонний анализ формы листьев у яблони, показывающий, что как образцы, так и виды демонстрируют аллометрические вариации из-за различий в ширине листовой пластинки. Хотя основная ось вариации яблока с использованием EFD и PH обусловлена этой аллометрической вариацией, мы находим высокие значения наследуемости в узком смысле даже для более поздних основных компонентов, что указывает на то, что всесторонние оценки формы отражают наследственную вариацию, которую можно было бы упустить только с помощью линейных оценок. .
Результаты
Вариации формы листьев яблони
Мы исследовали 24 фенотипа, связанных с формой и размером яблони, включая длину, ширину, площадь поверхности, сухой вес, массу листьев на единицу площади, вариацию внутри дерева и общую форму, оцененные с помощью ПК. полученные из данных EFD (эллиптический дескриптор Фурье) и PH (постоянная гомология) (см. материалы и методы и рис. 1-2). Предоставляются размер выборки и распределение каждого фенотипа, а также исходные данные (рисунок S1; таблица S1).
Рис. 1. Визуализация метода постоянной гомологии для ядра кольцевого пространства 7.
Двоичные изображения были преобразованы в двумерное облако точек (a), которое затем нормализовалось с использованием оценщика плотности по Гауссу (b). Для каждого листа использовали 16 ядер кольцевого пространства. Ядро кольца 7, обозначенное фиолетовым цветом (c), используется в качестве примера для этой визуализации. Оценщик плотности умножается на кольцо 7 (d). Функцию также можно визуализировать на виде сбоку (e, f). Когда плоскость движется сверху вниз, количество связанных компонентов записывается вдоль кривой (g).Ниже (g) представлены пять визуализаций кривых, которые представлены красными вертикальными пунктирными линиями на (g).
Рис. 2. Вклад гармоник эллиптического дескриптора Фурье в форму листа.
Изображенные формы листьев являются средними формами листьев для всех 915 деревьев. Гармоники с 1 по 15 представлены на оси x, и каждая гармоника умножается на коэффициент усиления на оси y, чтобы визуализировать их вклад в среднюю форму листа. Коэффициент усиления 0 указывает на удаление гармоники; коэффициент 1 дает нормальную форму; а значения выше 1 преувеличивают эффекты, чтобы лучше визуализировать вклад гармоники в окончательную форму.
Чтобы визуализировать основные оси морфометрической изменчивости, мы выбрали репрезентативный лист из образцов с минимальными и максимальными значениями вдоль первых 5 ПК для EFD и PH (рис. 3a). Образцы с экстремальными значениями по ПК1 для обоих методов аналогичны. Фактически, «Binet Rouge» имеет самое низкое значение вдоль PC1 для EFD и PH, при этом ось ясно показывает уменьшение отношения длины к ширине (аспекту). Ядра кольцевого пространства, наиболее сильно влияющие на PH PC1 (Рисунок S2), являются дополнительным доказательством того, что этот PC улавливает вариации в соотношении сторон.Изменения в форме листьев, фиксируемые ПК более высокого порядка, более сложны и загадочны, и поэтому не фиксируются с помощью одних только линейных измерений. Кроме того, в то время как первичная ось вариации (PC1) с использованием EFDs и PH может объяснять сходные аспекты морфологии листа, морфопространства, полученные в результате этих двух методов, различаются (Рис. 3b).
Рис. 3. Примеры формы листа на ПК, полученные из EFD и PH.
Двоичные изображения листьев образцов с минимальными и максимальными значениями вдоль ПК 1–5 для оценок EFD и PH.PC были рассчитаны с использованием значений, рассчитанных как среднее значение для 8-10 листьев, но отображается только один репрезентативный лист. ПК были скорректированы по REML в зависимости от положения деревьев в саду. Имя доступа также указано (а). Визуализация PC1 и PC2 для данных EFD и PH. Присоединение с минимальным и максимальным значениями по PC1 и PC2 указано (b).
Затем мы исследовали корреляцию между всеми измеренными признаками (Таблица S3). Оценивая корреляцию ПК в результате классической морфометрической техники, такой как EFD, с новым морфометрическим подходом, основанным на топологии, таким как PH, мы показываем, насколько дополняют друг друга методы (Рисунок 4; Рисунок S3).Хотя существует очень значимая корреляция между PC1 для обоих методов (R 2 = 0,949, p <1 × 10 −15 ), более поздние PC часто не имеют значительной корреляции, наиболее заметным исключением являются EFD PC2 и PH PC3. (R 2 = 0,432, p <1 × 10 −15 ), хотя несколько других ПК также показывают слабые корреляции. Таким образом, в то время как основная ось вариации (PC1) согласована и сильно коррелирована между методами, каждый метод фиксирует отдельные аспекты морфологии листа в последующих PC.
Рисунок 4. Корреляция между фенотипами листьев.
Значения над диагональю окрашены в соответствии с коэффициентом корреляции Пирсона, а значения под диагональю означают p-значения с поправкой Бонферрони. Рамка, обведенная пунктирными линиями, включает сравнения только между фенотипами, полученными с помощью всестороннего морфометрического анализа.
Многие фенотипы листьев сильно коррелируют друг с другом (рис. 4). В частности, соотношение сторон сильно коррелирует с PH PC1 (r = -0.878, p <1 × 10 −15 ), EFD PC1 (r = −0,855, p <1 × 10 −15 ) и малая ось (ширина листовой пластинки) (r = −0,734, p <1 × 10 −15 ). Корреляция между малой осью листа и площадью поверхности (r = 0,939, p <1 × 10 −15 ) выше, чем корреляция между большой осью (длиной лезвия) и площадью поверхности (r = 0,810, p < 1 × 10 −15 ). Как и ожидалось, площадь поверхности листа также сильно коррелирует со средней сухой массой листа (r = 0.934, p <1 × 10 −15 ), что указывает на то, что более крупные листья тяжелее.
Аллометрия в листьях яблони
Высокая корреляция между соотношением сторон и PC1 как для методов EFD, так и для PH указывает на то, что отношение длины к ширине является основным источником вариаций в форме листьев яблони. Если существует аллометрическая связь между малой и большой осью, и, таким образом, длина и ширина листа не увеличиваются с одинаковой скоростью, ожидается наклон, значительно отличающийся от 1.Мы обнаружили, что наклон между двумя измерениями значительно больше 1 (95% ДИ = 1,506-1,678, R 2 = 0,343, p <1 × 10 −15 ), что указывает на то, что малая ось увеличивается при увеличении скорость, чем большая ось. Хотя нет значимой корреляции между большой осью (длиной лопасти) и EFD PC1 (R 2 = 0,001, p = 1) или PH PC1 (R 2 = 0,002, p = 1), существует значимая корреляция. для малой оси (ширина лопасти) и EFD PC1 (R 2 = 0.541, p <1 × 10 −15 ) и PH PC1 (R 2 = 0,573, p <1 × 10 −15 ) (рисунок 5). Поскольку PC1 объясняет 80,23% вариации формы листа для EFD и 62,20% для PH, очевидно, что ширина листовой пластинки, а не длина, является основным источником вариации формы листа у яблони. Фактически, соотношение сторон, рассчитанное как отношение большой оси к малой оси, еще сильнее коррелирует с EFD и PH PC1, с R 2 0,732 для EFD PC1 (p <1 × 10 −15 ) и 2 рэнд в размере 0.771 для PH PC1 (p <1 × 10 −15 ). Учитывая значительную корреляцию между EFD PC1 и PH PC1 (таблица S3), неудивительно, что соотношение сторон сильно коррелирует с обоими.
Рис. 5. Корреляция между первичной осью вариации (PC1), полученной с использованием значений EFD и PH и мер формы листа.
EFD PC1 нанесен на большую ось (длина листовой пластинки) (a), малую ось (ширина листовой пластинки) (b) и соотношение сторон (отношение длины к ширине лезвия) (c). PH PC1 отображается в сравнении с теми же показателями на панелях d-f.Отклонения в процентах, объясненные PC1 до корректировки REML, показаны в скобках. Все p-значения скорректированы по Бонферонни на основе количества сравнений на рисунке 4. Также показана линия регрессии из линейной модели с заштрихованным 95% доверительным интервалом.
Помимо различий между образцами, мы исследовали различия в форме и размере листьев между видами, сравнивая Malus domestica , домашнюю яблоню, с ее первичным видом-предшественником, Malus sieversii (Таблица S4).PCA полногеномных данных SNP выявляет первичную ось генетической изменчивости, которая разделяет M. domestica и M. sieversii , хотя разделение неполное (Рисунок 6a). Большая ось (p = 0,975) листьев не различается между видами (рис. 6b). Однако малая ось (p = 4 × 10 -4 ) листьев M. domestica значительно больше, чем M. sieversii (рис. 6c), а соотношение сторон (p = 0,023) значительно меньше (рис. 6г).Таким образом, существует аллометрическая вариация как внутри (Рисунок 5), так и между (Рисунок 6) видов Malus .
Рис. 6. Генетическое и фенотипическое сравнение одомашненной яблони и ее дикого предка.
ПК 1 и 2 были получены из 75 973 полногеномных SNP, и образцы помечены как M. domestica (фиолетовый), M. sieversii (зеленый) или неизвестно (серый). листьев M. domestica не отличаются от листьев M. sieversii вдоль большой оси (b), но у них есть большая малая ось (c) и соотношение сторон (d).Приведенные P-значения скорректированы по Бонферрони на основе множественных сравнений (Таблица S4). Маркировка видов основана на классификации USDA.
Генетическая основа формы листьев яблони
GWAS 24 фенотипов листьев, исследованных в этом исследовании, не дало значительных результатов. Мы идентифицировали 70 значимых SNP, представляющих 5 фенотипов, которые представлены в таблице S5. Мы исследовали регионы, окружающие важные SNP для генов-кандидатов, с помощью инструмента GBrowse (Таблица S6) (22). Мы провели поиск в пределах окна +/- 5000 п.н., которое должно улавливать любые связанные причинные вариации с учетом быстрого распада LD, наблюдаемого в разнообразной коллекции яблок, которая в значительной степени воспроизводится в исследуемой здесь зародышевой плазме (23).Однако никаких сильных генов-кандидатов выявлено не было.
В то время как GWAS исследует геном на предмет наличия одиночных локусов с большим эффектом, геномное предсказание оценивает нашу способность предсказывать фенотип, используя данные маркеров для всего генома. Мы дополнили наш GWAS геномным предсказанием и наблюдали точность предсказания (r) в диапазоне от -0,10 до 0,52 (таблица S7; рисунок S5a). Соотношение сторон является основным источником вариаций формы листа (рис. 5c), и это также измерение листа, которое имело наивысшую точность геномного предсказания (0.52). Другие фенотипы сильно коррелировали с соотношением сторон, такие как ширина листа (0,51), малая ось (0,49), EFD PC1 (0,48) и PH PC1 (0,47), все имели относительно высокую точность прогнозов. PH PC3 (0,51) также был одним из наиболее хорошо предсказанных с использованием генетических данных.
Аналогичным образом, оценки наследуемости в узком смысле (h 2 ), рассчитанные с использованием GCTA (24), варьировались от 0 до 0,75, при этом самая высокая наследуемость наблюдалась для соотношения сторон (0,75), за которым следует ширина листа (0,71), EFD PC1 ( 0,71), малая ось (0.69) и PH PC1 (0,65) (рисунок 7; таблица S8). Оценки наследственности сильно коррелировали с точностью предсказания генома (рисунок S5b, R 2 = 0,936, p <1 × 10 -15 ), что неудивительно, учитывая, что оба метода предполагают предсказание фенотипа на основе данных SNP для всего генома. Ни один из фенотипов, измеряющих дисперсию в отобранных 8-10 листьях, не имел оценок наследуемости, значительно отличавшихся от 0.
Рисунок 7. Узкая наследуемость (h 2 ) фенотипов листьев.Значения
представляют собой аддитивную генетическую дисперсию (V g ), деленную на фенотипическую дисперсию (V p ) со стандартной ошибкой, рассчитанной с использованием GCTA (24). Пунктирные красные линии находятся при h 2 = 0, что указывает на то, что ни одна из фенотипических вариаций не объясняется генетическими данными. Доля общей фенотипической дисперсии, объясняемой каждым ПК, указана в скобках.
Хотя основным компонентом вариации формы листа, обнаруживаемой EFD и PH, является соотношение сторон, мы также были заинтересованы в том, чтобы определить, извлекают ли ПК более высокого порядка, улавливающие вариации, не всегда видимые глазом, информацию, имеющую биологическое значение.Используя геномное предсказание и оценки наследуемости, мы нашли доказательства генетической основы этих «скрытых фенотипов», которые невозможно измерить с помощью линейных методов. Например, наследуемость фенотипов, таких как PH PC6 (0,48), PH PC9 (0,35), PH PC10 (0,33) и EFD PC9 (0,33), аналогична традиционно измеряемым фенотипам, таким как длина листа (0,44) и масса листа на площадь. (0,40). Хотя более высокие ПК могут иметь относительно высокие значения наследуемости, после определенного момента значения (+/- стандартная ошибка) перекрываются с 0, что указывает на то, что они не наследуются.Пороговое значение для морфометрических PC с наследственной генетической основой составляет приблизительно PC17. Эти результаты предполагают, что, используя морфометрические методы, которые позволяют всесторонне измерять форму, мы описываем биологически значимые наследственные фенотипы, которые можно было бы упустить с помощью простых измерений, таких как длина, ширина и площадь листа.
Обсуждение
Форма и размер листьев играют решающую роль в росте и развитии яблонь, включая плоды. Чтобы выяснить генетическую основу этой вариации, мы количественно определили форму листьев яблони, используя традиционные линейные измерения и комплексные морфометрические методы.Наша работа предлагает первое сравнение между новой методикой, основанной на топологии, PH и EFD, которые, как мы считаем, дополняют друг друга, но отличаются друг от друга. Для обоих методов PC1 сильно коррелировал с соотношением сторон, что свидетельствует о том, что первичная ось изменения формы яблочного листа может быть зафиксирована с помощью линейных измерений. Малая ось или ширина листовой пластинки также сильно коррелировала с PC1, в то время как большая ось — нет. Таким образом, изменение соотношения сторон происходит из-за изменения ширины листовой пластинки, а не длины.Площадь поверхности листа также больше коррелировала с малой осью, чем с большой осью. Следовательно, изменение ширины листа важно как для размера, так и для формы яблоневых листьев, как и в предыдущей работе с помидорами (25).
Ширина листовой пластинки является источником различий не только между образцами яблони, но и между M. domestica и M. sieversii . Наличие одинаковых аллометрических отношений внутри и между видами предполагает, что генетические локусы, контролирующие внутривидовые вариации формы листа в пределах M.Domestica может быть тем же самым, что и те, которые контролируют дивергенцию формы листьев, наблюдаемую между домашним яблоком и его диким предком. Например, у птиц, хотя PC1 и PC2 формы клюва объясняют большую часть различий между 2000 видами, они также неизменно связаны с различиями между более высокими таксонами (обладающими> 20 видами) (26). Наши результаты показывают, что увеличение размера листа с момента одомашнивания не было общим увеличением размера листа, а, в частности, увеличением ширины лезвия, приводящим к более крупным листьям с уменьшенным соотношением длины к ширине.
Наша работа свидетельствует о том, что аллометрия является основным источником морфометрических изменений в листьях яблони. Эти результаты согласуются с исследованиями других видов, таких как томат, где соотношение длины к ширине было основным источником вариации формы (> 40%) (5). Точно так же работа с видами Passiflora и Vitis , выполненная с использованием двух независимых морфометрических методов, определила аллометрические вариации как основной источник вариаций в PC1, который объясняет по крайней мере 40% вариаций формы листьев (8, 27).Таким образом, линейные измерения — в частности, соотношение сторон — являются важным источником информации при описании формы листа. Однако линейных измерений недостаточно для охвата всего спектра разнообразия. В нашем исследовании PC1 составляет 62,20% или 80,23% вариации, в зависимости от используемой техники. Просто оценивая листья яблони с помощью линейных измерений, в некоторых случаях мы упускаем почти 40% вариации. В то время как PC1 сильно коррелирует с соотношением сторон, более поздние ПК представляют собой ортогональные вариации, которые, вероятно, могут быть зафиксированы только с помощью морфометрических методов, таких как EFD и PH.Поэтому для полной количественной оценки вариаций формы листьев необходимы комплексные морфометрические методы.
Чтобы выявить генетический вклад в форму листа, мы объединили линейные и комплексные морфометрические оценки формы с данными SNP для всего генома. Есть примеры простой генетической основы формы листа, такие как Arabidopsis thaliana , где ANGUSTIFOLIA и ROTUNDIFOLIA3 независимо контролируют ширину и длину листа (28). В ячмене уровни транскрипта BFL1 ограничивают ширину листа, при этом сверхэкспрессия приводит к более узким листьям и потере функции BFL1 , что приводит к уменьшению отношения длины к ширине (29).Используя GWAS, мы не обнаружили устойчивых ассоциаций с фенотипами формы, наблюдали низкое соотношение значимых SNP к количеству исследованных фенотипов и обнаружили, что значимые SNP были редко распределены по множеству хромосом. Кроме того, небольшое количество значимых SNP, вероятно, являются ложными ассоциациями из-за плохой коррекции загадочного родства, о чем свидетельствуют графики QQ (рис. S4). Эти наблюдения предполагают, что форма листа, скорее всего, полигенная и контролируется большим количеством локусов небольшого эффекта, например, у томатов и кукурузы (5, 21).Для сравнения, GWAS по фенотипам плодов яблони, таким как цвет и плотность, выявил сильные ассоциации, являющиеся результатом небольшого количества локусов большого эффекта (23). Однако возможно, что в настоящем исследовании были упущены локусы с большим эффектом либо из-за плохой сборки референсного генома, либо из-за недостаточной плотности маркеров. Улучшения в сборке генома и увеличение количества маркеров помогут в дальнейшем раскрыть генетическую архитектуру вариаций формы листьев яблони.
Наконец, мы исследовали степень, в которой форма листа передается по наследству и может быть предсказана с использованием данных SNP для всего генома.Мы обнаружили, что точность геномного предсказания основных осей вариации формы листа аналогична ранее опубликованным оценкам ширины (0,48) и длины (0,47) плода, что указывает на то, что форма листа так же наследуется, как и форма плода (23). В сочетании с несколькими значимыми результатами GWAS высокие оценки наследственности в узком смысле подтверждают полигенную основу формы листа. Соотношение сторон было определено как основной источник вариаций формы листьев у яблони и имело самые высокие геномные прогнозы и оценки наследственности, что указывает на наличие генетической наследственной основы для аллометрических вариаций яблони.Хотя мы не обнаружили генетическую основу вариации формы листьев в образце, мы намеренно взяли образцы листьев, представляющих среднее значение для дерева, и это могло снизить силу. Кроме того, хотя первые 5 ПК как для EFD, так и для PH объясняют большую часть вариаций формы листьев яблони, большинство ПК от 1 до 14 имеют оценки наследуемости выше 0,20 и могут по-прежнему представлять важные отличия в форме листьев от экологических, эволюционных или экологических. аграрная перспектива. Таким образом, хотя наша способность обнаруживать основные оси вариации формы листа с использованием данных всего генома является ожидаемой, наше наблюдение, что ПК более высокого уровня также наследуются, подтверждает, что эти комплексные морфометрические методы улавливают биологически значимые вариации, которые могут быть упущены только при линейных измерениях. .
Выводы
Из нашей работы ясно, что вариации формы и размера листьев яблони находятся под генетическим контролем. Кроме того, высокий прогноз генома и оценки наследуемости для ПК с более высокой морфометрией показывают, что такие методы, как EFD и PH, улавливают наследственные биологические вариации, которые будут упущены, если исследователи ограничат оценки формы листьев линейными измерениями. На основе этих результатов можно будет выполнить геномный отбор по фенотипу, который может быть обнаружен только с помощью морфометрии.Если ПК более высокого порядка коррелировал с признаком, который было трудно или дорого измерить, оценка формы листа потенциально могла бы использоваться в качестве прокси для этого фенотипа, так же, как красный цвет листа может использоваться для выбора цвета мякоти красных плодов в яблоки (13,14). Кроме того, лучшее понимание изменений формы и размера листьев у яблони может в конечном итоге иметь важные последствия для управления растительным покровом, где воздействие света имеет решающее значение для цветения (17). В конечном итоге, благодаря первому углубленному изучению формы листьев у яблони, мы обнаружили аллометрию между образцами и видами, а также доказательства того, что сложные и наследуемые фенотипы могут быть обнаружены с помощью комплексных морфометрических методов.
Материалы и методы
Сбор образцов
Яблони в Кентвилле, Новая Шотландия, Канада, весной 2012 г. были посажены на подвои M.9. Осенью деревья были выкорчеваны и хранились в холодильнике до весны 2013 г., когда деревья были посажены в неполной блочной конструкции (см. «Ограниченное максимальное правдоподобие (REML)» ниже). С 24 августа по 16 сентября 2015 года были собраны листья более 900 деревьев. С каждого дерева было собрано по 10 листьев. Листья сплющивали и помещали так, чтобы не касаться, а затем сканировали с помощью сканеров цветных изображений Canon CanoScan (LiDE 220).Затем листья сушили в течение 48 часов при 65 ° C и взвешивали, чтобы оценить общий сухой вес (г) для каждого дерева.
Морфометрический анализ
Сканы листьев были преобразованы в отдельное двоичное изображение для каждого листа с использованием настраиваемых макросов ImageJ, которые включали функцию «make binary» (30). Для каждого листа был создан новый файл изображения, названный в честь идентификатора дерева. Изображения были преобразованы в файлы RGB .bmp, а анализ цепного кода был выполнен с помощью SHAPE (31). Цепной код использовался для вычисления нормализованных эллиптических дескрипторов Фурье (EFD) в SHAPE.Нормализованные EFD считывались в Momocs v1.1.5 (32) в R (33), где гармоники B и C были удалены, чтобы устранить асимметричные вариации в форме листьев.
Бинарные изображения листьев также были проанализированы с использованием постоянной гомологии (PH) (10). Чтобы численно оценить форму листьев с помощью PH, мы извлекли контур листа с помощью двумерного облака точек (рис. 1а). После центрирования и нормализации контура по размеру его центроида мы использовали оценщик плотности по Гауссу (рис. 1b), который присваивает высокие значения (красный) пикселям с большим количеством соседних пикселей, а низкие значения (синий) — пикселям с меньшим количеством соседних пикселей.Мы умножили оценку плотности на ядро кольца или кольцо (рис. 1c), которое подчеркивает форму кольца в центре тяжести и, таким образом, инвариантно к ориентации (рис. 1d). Полученную функцию также можно визуализировать сбоку (рис. 1e, f). Когда мы перемещали плоскость сверху вниз, мы записывали количество связанных компонентов над плоскостью, образуя кривую. С каждым новым компонентом это значение увеличивалось, и каждый раз, когда компоненты объединялись, оно уменьшалось (рис. 1g).
Для каждого листа мы рассчитали 16 кривых, соответствующих 16 расширяющимся кольцам.Для вычислительных целей каждая кривая делится на 500 чисел, в результате чего форма каждого листа представлена 8000 (16 * 500) значениями.
Только листья, для которых были успешно рассчитаны как EFD, так и PH, были включены в последующий анализ. Кроме того, были включены только деревья с 8-10 листьями, поскольку листья иногда удалялись из-за разрывов, складок или отсутствия черешка, что не позволяло точно определить форму. Окончательный набор данных состоял из 915 деревьев с 8-10 листьями, включая 869 уникальных образцов и 8 995 листьев.
значений EFD и PH были усреднены по листьям отдельного дерева. Вклад гармоник с 1 по 15 EFD в среднюю форму листа на всех деревьях был визуализирован с помощью функции «hcontrib» в пакете Momocs R (рис. 2). Чтобы обеспечить различение образцов на основе формы листьев, был проведен анализ главных компонентов (PCA) с использованием функции Momocs ‘PCA’ (32) для EFD и функции prcomp в R для значений PH, которые центрируются, но не масштабируются. данные. Полученные значения ПК были скорректированы с использованием REstricted Maximum Likelihood (см. Ниже).Впоследствии мы определили присоединение с минимальным и максимальным значением для каждого из первых 5 ПК.
В дополнение к оценке контура листа с помощью EFD и PH, мы использовали еще несколько показателей для описания листьев. Используя ImageJ, мы автоматизировали измерение площади поверхности листа ( 2 см), длины (см) листа и ширины (см) листа, а также основных (длина лезвия) и второстепенных (ширина лезвия) осей листа. наиболее подходящий эллипс — без черешка — в периодических процессах (30).На протяжении всей рукописи мы используем «большой», когда говорим о длине листовой пластинки, и «малый», когда ссылаемся на ширину листовой пластинки. Мы также рассчитали соотношение сторон листа, разделив большую ось на меньшую. Кроме того, масса листьев на площадь была рассчитана для 780 деревьев, у которых мы располагали данными площади поверхности для всех 10 листьев, путем расчета отношения сухой массы к площади поверхности (г / см 2 ).
Хотя линейные фенотипы были рассчитаны как среднее значение для конкретного дерева, мы также оценили дисперсию внутри дерева по соотношению сторон, длине, ширине, большой и малой оси и площади поверхности.Дисперсия была рассчитана как коэффициент вариации с использованием функции «cv» в растровом пакете (34) в R для оценки изменчивости в пределах дерева в размере листа, которая обозначена как «var» в этой рукописи.
REstricted Maximum Likelihood (REML) корректировка фенотипических данных
Фруктовый сад, отобранный в этом исследовании, представляет собой неполный блочный дизайн с 1 из 3 стандартов на сетку. Стандарты или «контрольные деревья» — «Honeycrisp», «SweeTango» и «Ambrosia» — воспроизводятся по всей сети.Листья этих деревьев были взяты несколько раз по всему саду, что позволило нам скорректировать позиционные эффекты. Каждый фенотип был скорректирован с использованием модели REstricted Maximum Likelihood (REML), которая дала одно скорректированное значение для каждого образца, даже если было измерено несколько деревьев. Влияние сетки строк (rGrid), сетки столбцов (cGrid) и эффектов rGrid × cGrid было скорректировано для использования следующей модели REML:
Мы аппроксимируем линейную модель смешанных эффектов через REML, используя функцию «lmer» в пакете lme4 в R (35), а затем вычислили средние значения наименьших квадратов с помощью функции «lsmeans» в пакете lsmeans R (36).
Таким образом, хотя исходные данные о фенотипе были собраны для 915 деревьев, после корректировки REML осталось одно значение для каждого уникального образца, что дало 869 образцов. REML-корректировка применялась непосредственно ко всем оценкам размера, веса и дисперсии. Для PH и EFD мы применили REML после PCA, и, таким образом, процентный вклад для каждого ПК был рассчитан с использованием нескорректированных значений. Скорректированные данные для всех 24 фенотипов включены в Таблицу S1.
Феномный анализ
Корреляцию между фенотипами листьев рассчитывали с использованием корреляции Пирсона, а значения p были скорректированы по Бонферрони для множественных сравнений.Полученная тепловая карта была визуализирована с помощью функции geom_tile в ggplot2 в R (37). Затем мы исследовали листья для аллометрии, используя функцию SMA в пакете smartr R (38), чтобы оценить, отличается ли наклон между малой и большой осью с логарифмическим преобразованием от 1.
Образцы были помечены как Malus . Икс. domestica Борх. или Malus sieversii Лебедь. на основе информации, предоставленной веб-сайтом информационной сети по ресурсам зародышевой плазмы Министерства сельского хозяйства США (USDA) (http: // www.ars-grin.gov/) (Таблица S2). Мы использовали U-критерий Манна-Уитни, чтобы проверить, различались ли какие-либо фенотипы между видами, и скорректировали все p-значения с поправкой Бонферрони для множественных сравнений.
Геномные анализы
ДНК экстрагировали с использованием коммерческих наборов для экстракции. Библиотеки генотипирования путем секвенирования (GBS) получали с использованием рестрикционных ферментов ApeKI и PstI-EcoT221I в соответствии с Elshire, et al., . (39). Библиотеки GBS секвенировали с использованием технологии Illumina Hi-Seq 2000. Чтения, которые не прошли «фильтр целомудрия» Illumina, были удалены из необработанных файлов fastq.Остальные чтения были согласованы с Malus x. domestica v1.0 псевдогаплотипическая эталонная последовательность (40) с использованием инструмента выравнивания Burrows-Wheeler v0.7.12 (41) и конвейера Tassel версии 5 (42). Параметры Tassel включали minKmerL 30, mnQS 20, mxKmerNum 50000000 и batchSize 20. Длина kmerlength была установлена на 82 для ApeKI и 89 для PstI-EcoT22I на основе максимального размера штрих-кода. MinMAF для DiscoverySNPCallerPluginV2 был установлен на 0,01. Были использованы все остальные параметры по умолчанию.Небиаллельные сайты и индели удаляли с помощью VCFtools v.0.1.14 (43). Затем VCF для обоих ферментов были объединены с использованием специального сценария perl, предпочтительно сохраняя SNP, вызываемые PstI-EcoT22I, на перекрывающихся сайтах, поскольку эти сайты, как правило, были в более высоком покрытии.
Отсутствующие данные были рассчитаны с использованием LinkImputeR v0.9 (Money et al., Submitted, доступно: http://www.cultivatingdiversity.org/software.html) с глобальными пороговыми значениями 0,01 для частоты минорных аллелей (MAF) и 0,70 для отсутствие.Мы исследовали глубины от 3 до 8 и выбрали случай для вменения с максимальным отсутствием позиции / выборки 0,70, минимальной глубиной 5 и точностью вменения 94,9%. VCF был преобразован в таблицу генотипов с помощью PLINK v1.07 (44, 45).
Из 869 образцов, оцененных в этом исследовании, 816 имели геномные данные после вменения и фильтрации и были включены в последующий анализ. Полученная таблица генотипов состояла из 816 образцов и 197 565 SNP. Впоследствии был применен фильтр 0,05 MAF с помощью PLINK, после чего осталось 128 132 SNP.SNP с более чем 90% гетерозиготных генотипов были удалены. Окончательная таблица генотипов состояла из 816 образцов и 122596 SNP.
Для выполнения PCA SNP были отсечены на предмет неравновесия по сцеплению (LD) с помощью PLINK. Мы рассмотрели окно из 10 SNP, удалив один SNP из пары, если R 2 > 0,5, затем сдвинули окно на 3 SNP и повторили (команда PLINK: indep-pairwise 10 3 0,5). В результате получился набор из 75 973 SNP для 816 образцов. PCA выполняли на LD-обрезанных SNP всего генома с использованием «prcomp» в R с данными, которые были центрированы, но не масштабированы.Первые 2 геномных ПК были визуализированы с помощью ggplot2 в R (37).
Мы провели полногеномное ассоциативное исследование (GWAS) с использованием смешанной линейной модели в Tassel (версия 5) для каждого фенотипа с поправкой на родство между людьми с использованием матрицы родства, а также первых 3 ПК для структуры популяции (46, 47). Порог значимости был рассчитан с помощью simpleM (48, 49), который оценивает количество ПК, необходимых для объяснения 0,995 дисперсии, или количества независимых SNP.Предполагаемое значение Meff, использованное для расчета порога значимости, составило 91 667 SNP.
Мы провели поиск в регионах, окружающих любые важные SNP GWAS, используя базу данных генома для Rosaceae GBrowse, инструмент для Malus x. domestica v1.0 p геном (22). Мы использовали окно размером +/- 5000 п.н. (10 т.п.н.), окружающее значимый SNP, для проверки генов, а после идентификации мы использовали базовый инструмент поиска локального выравнивания (BLAST) от NCBI для поиска последовательности мРНК и представили результат. с максимальным баллом (50).
Геномное предсказание было выполнено с использованием функции «x.val» в R-пакете PopVar (51). Была выбрана модель rrBLUP, и 5-кратная (nFold = 5) перекрестная проверка была повторена 3 раза (nFold.reps = 3) без дополнительной фильтрации (min.maf = 0) из набора из 122596 SNP, используемых для GWAS. Были использованы все остальные параметры по умолчанию. В дополнение к выполнению геномного прогнозирования по основным 24 фенотипам, исследованным в этом исследовании, мы выполнили геномное прогнозирование на всех 40 ПК для EFD и на первых 40 ПК для значений PH.Мы также использовали функцию «rnorm» в R для генерации 1000 случайных фенотипов со средним значением 0 и стандартным отклонением 1 и выполнили геномное предсказание с использованием этих случайных фенотипов, чтобы получить диапазон точности геномного предсказания, который можно ожидать в случайном порядке. Наконец, мы использовали анализ комплексных признаков на уровне всего генома (GCTA) v.1.26.0, который оценивает генетические отношения между людьми на основе общегеномных SNP и использует эту информацию для расчета дисперсии, объясняемой этими SNP. Отношение аддитивной генетической изменчивости к фенотипической дисперсии используется для расчета наследуемости в узком смысле (h 2 ) или наследуемости SNP признака (24).Мы использовали GCTA для оценки наследуемости для каждого фенотипа, включая первые 40 ПК для EFD и PH. Мы также оценили корреляцию между точностью предсказания генома (r) и наследуемостью в узком смысле (h 2 ), используя корреляцию Пирсона.
Благодарности
Эта статья была написана отчасти благодаря финансированию программы Canada Research Chairs, Национального совета по научным и инженерным исследованиям Канады и Genome Canada. З.М. также была частично поддержана стипендией Киллама для получения докторской степени от Университета Далхаузи.
Мы хотели бы поблагодарить Гэвина Дугласа и Шерри Филлмор за их помощь в настройке статистического дизайна фруктового сада и конвейера вызовов SNP. Мы также благодарим всех бывших и нынешних членов Myles Lab за их работу по уходу за яблоневым садом.
Моделирование переноса ингибирующих и активирующих сигналов и их комбинированного воздействия на индукцию цветков: приложение к яблони
Обзор модели
Модель (SigFlow) была разработана на Python и использует библиотеки из платформы OpenAlea 30 .Модель работает на трехмерных древовидных архитектурах, закодированных в Multiscale Tree Graphs 16 (MTG) с тремя масштабами (дерево, сегменты ствола и метамеры) и дополненной трехмерными координатами органов 31 . Сегменты — это части стебля между двумя точками ветвления или одной точкой ветвления и концом оси. Метамеры представлены для сегментов стебля, соответствующих только самым последним побегам, и состоят из одного листа, одного междоузлия и соцветия, если оно есть (дополнительный рис.S1). Плоды и листья создают сигналы, перемещающиеся внутри конструкции в зависимости от расстояния с параметром «затухание», который можно настраивать для имитации различных распределений сигналов, то есть однородности внутри конструкции или местного источника питания. Эти сигналы достигают SAM и с вероятностью определяют их судьбу в зависимости от количества сигналов. ФИ моделируется на ЗРК, расположенном в крайнем положении однолетних побегов. В своей текущей версии модель работает на статических древовидных структурах, что соответствует нашей цели моделирования, поскольку FI обычно возникает после окончания вегетативного роста побегов у взрослых яблонь 32 .
Запрещение и активация количества сигналов и перенос
Уравнения для переноса сигналов (ингибирования и активации) между однолетними побегами и SAM были адаптированы из предыдущих исследований, посвященных распределению углерода между источниками и поглотителями 20,29 . Эти формализмы рассматривают распределение углерода как зависящее от расстояний между источниками и поглотителями и от значений прочности поглотителей в зависимости от типа и возраста органа. В нашем случае SAM рассматриваются как органы-мишени для ингибирования и активации сигнала с аналогичной способностью накапливать ингибирующий или активирующий сигнал независимо от SAM.Однолетние побеги считаются источниками тормозящих и активирующих сигналов, исходящих от фруктов и листьев соответственно. Количество сигналов, исходящих от каждого однолетнего побега, рассчитывается как сумма сигналов, исходящих от каждого отдельного плода и листа в данном побеге. Поскольку ранее предложенное уравнение 29 учитывало относительные значения силы погружения, распределение сигналов от каждого годового выстрела было переписано с учетом аналогичных способностей SAM по накоплению сигналов следующим образом:
$$ {\ text {q}} _ {{ {\ text {ij}}}} = \ frac {{{\ text {Q}} _ {i} \ times \ left ({\ frac {1} {{1 + {\ text {d}} _ {{ {\ text {ij}}}}}}} \ right) ^ {r}}} {{\ mathop \ sum \ nolimits_ {k = 1} ^ {n} {\ text {Q}} _ {{\ text {i}}} \ times \ left ({\ frac {1} {{1 + {\ text {d}} _ {{{{\ text {ik}}}}}}}} \ right) ^ {r}} } $$
(1)
, где n — количество SAM в дереве, q ij — количество сигнала, экспортируемого однолетними побегами (запрещающий или активирующий сигнал) i в SAM j , Q i количество сигнала, производимого за год i , d ij расстояние по топологическому пути между i и j и r параметр затухания, модулирующий эффект расстояния . {N} \ left ({{\ text {q}} _ {{{\ text {ij}}}}} \ right) $$
(2)
с N числом однолетних побегов.В следующем примере Q + F и Q — F используются для количества активирующего и запрещающего сигнала соответственно.
Расстояния между SAM и побегами рассчитываются на основе топологического положения органа и трехмерных координат в MTG. Расстояние d ij между побегами и SAM ( i , j ) в древовидной структуре вычисляется по топологическому пути как сумма евклидовых расстояний между (i) основанием и барицентром каждый годовой побег, (ii) плюс расстояния между последовательными основаниями компонентов растения и SAM (iii) плюс расстояние между основанием годового побега, несущего SAM, и его концом 29 .{-} — t_ {-}} \ right)}} {{v_ {-}}}} \ right)}} $$
(3)
, где \ (t_ {+} {\ text {и}} t_ {-} \) являются параметрами, называемыми «переходными» значениями (рис. 1b), указывающими количество сигнала ( Q F + или Q F — ) для которых SAM имеет 50% шанс срабатывания или запрета и v + и v — параметры, называемые «фактором формы» учет вариаций наклона функции.Когда v + или v — близки к 0, вероятность быстро меняется от 0 до 1, когда значения Q F + или Q F — превышает или отстает от t + или t — , тогда как переходы более прогрессивны, когда v + или v — значения увеличиваются ( Инжир.1в). Эти параметры используются для представления некоторой неопределенности в судьбе SAM (индуцированной цветком или нет) для данного значения ингибирующих и активирующих сигналов.
Рисунок 1
Представление уравнений переноса сигналов и влияния на индукцию цветков. ( a ) Взаимосвязь между расстоянием до меристем и количеством сигнала (активирующим и подавляющим) для различных значений «затухания» сигнала ( r — , r + ), ( b ) взаимосвязь между количество сигнала (активирующего и подавляющего) и пропорции индукции цветков для различных « переходных » значений ( t — , t + ), ( c ) соотношение между количеством сигнала (активирующим и подавляющим) пропорции цветочной индукции для различных значений «формы» ( v — , v + ).{+} _ {j}. $$
(5)
Оценка поведения модели на простых структурах
Модель была применена к простым древовидным структурам для анализа согласованности модели и ее чувствительности к значениям параметров. Первая структура состояла из ветви, состоящей из шести равноудаленных друг от друга побегов, несущих или не несущих листья или плоды. Здесь мы стремились проверить влияние параметра «затухание» сигнала ( r — , r + ) на количество сигнала, достигающего SAM, расположенных на разном расстоянии от источников сигнала.
Вторая структура состояла из двух полных ветвей с различным пространственным расположением плодов и листьев. В первом случае распределение плодов и листьев на двух ветвях было одинаковым, половина побегов приносила либо плоды, либо листья. Во втором случае все плоды или листья располагались только на одной ветке, а другая была полностью опорожнена или опущена. Здесь мы стремились протестировать широкий диапазон «затухания» сигнала ( r — , r + ) и параметра «переход» ( t — , t + ), чтобы оценить взаимосвязанные эффекты, создаваемые моделью, между древовидной архитектурой, транспортировкой сигналов и чувствительностью SAM к количеству сигналов.
Наконец, мы проанализировали формализмы модели («ограничивающий фактор», «мультипликативный формализм»; уравнения 4 и 5), используемые для интеграции комбинированных эффектов подавления и активации сигналов на SAM FI. Мы использовали структуру, состоящую из четырех контрастных ветвей, то есть листовой и плодоносящей ветви, лиственной ветви без плодов, плодовой ветви без листьев и дефолиированной и неплодоносящей ветви. Эти два формализма использовались с широким диапазоном «переходных» значений ( t — , t + ) для изменения чувствительности SAM к обоим сигналам.
Описание экспериментального набора данных
Данные, использованные для построения трехмерных представлений деревьев, а также для калибровки и проверки, были взяты из экспериментов, проведенных в 2017 году на 10-летних яблоневых деревьях (сорт Golden Delicious), расположенном в на экспериментальной станции SudExpé в Марсильярге, на юге Франции (43 ° 66 ′ с.ш., 4 ° 18 ′ в.д.). В этом эксперименте, описанном в Belhassine et al. 10 , листья и / или плоды были удалены в разных частях дерева (дополнительный рис.S2). На деревьях, находящихся в состоянии ВКЛЮЧЕНО (высокая плодовая нагрузка) или ВЫКЛЮЧЕНО (нагрузка урожая близка к 0), плоды или листья были удалены на половине побегов и половине ветвей деревьев, обученных как «солаксе» (один основной вертикальный ствол) и на одной стороне деревьев Y-образной формой (два основных ствола). Дополнительный набор деревьев, не подвергнутых удалению плодов или листьев, но демонстрирующих естественную изменчивость нагрузки на урожай, наблюдался в течение трех лет (2015–2016–2017).
Доля SAM FI на деревьях оценивалась в период полного цветения весной после начала обработки, как отношение общего количества репродуктивных почек к общему количеству растущих почек в различных частях деревьев (листовых, нелистовых или плодоносящие, неплодоносящие).Нагрузка на урожай деревьев оценивалась как количество плодов при сборе урожая, деленное на площадь поперечного сечения ствола (TCSA, см 2 ), оцененную осенью после сбора урожая 33 .
Входные архитектуры
Описание архитектуры было выполнено на одном дереве «solaxe» и одном дереве Y-формы, отображающем значения TCSA (20,4 и 24,7 см. 2 , для деревьев solaxe и Y-shape соответственно), близкие к наблюдаемым средним значениям. в саду. Трехмерные координаты были получены с помощью электромагнитного трехмерного оцифровщика (3Space Fastrak; Polhemus Inc., Colchester, VT, USA) у основания ствола, точек ветвления, а также координаты вершины и основания каждого однолетнего побега (дополнительный рисунок S1). Трехмерные структуры, включая древовидные объекты, организованные в трех топологических масштабах, и их координаты были сохранены в формате MTG. Эти структуры использовались для реконструкции расположения листьев и площади вдоль однолетних побегов, а также для воспроизведения экспериментальных обработок in silico по удалению листьев и плодов. Распределение площади листьев вдоль однолетних побегов было реконструировано на основе аллометрических соотношений, как было предложено ранее 34 (Дополнительные методы S1).Для построения аллометрических соотношений были собраны данные как по 60 коротким (<5 см), так и по длинным побегам (> 5 см), а площадь отдельных листьев была оценена с помощью измерителя площади листа (LI 3100 Area Meter, LI-COR, Lincoln, NE, США) каждые три листа по побегам. Образцы зрелых листьев были взяты после сбора плодов, чтобы построить эти аллометрические отношения. На деревьях ON один плод был добавлен в MTG у основания каждого однолетнего побега (в соответствии с практикой прореживания в поле).
Оценка значений параметров и оценка модели
Выходные данные модели расширяют MTG новыми атрибутами для каждого SAM j , соответствующими количеству активирующих и запрещающих сигналов, достигающих его после транспортировки (Q + F, j и Q — F, j ), его вероятность FI связана с полученными активирующими ( P + j ) или запрещающими сигналами ( P — j ) и его окончательная вероятность FI, объединяющая оба сигнала ( P j ).Параметры, связанные с воздействием сигнала на активацию или ингибирование FI, оценивали отдельно при обработке удаления листьев и плодов, соответственно. Параметры активирующего сигнала ( r + , t + , v + ) оценивали на неплодоносящих структурах, подвергнутых удалению листьев в разных масштабах (побег, ветка, полудерево) или нет (управление деревьями ВЫКЛ). Параметры запрещающего сигнала ( r — , t — , v — ) оценивали на структурах лиственных и плодовых деревьев, подвергнутых удалению плодов в различных масштабах (побег, ветка и полудерево) или нет (контрольные деревья) и на лиственных неплодоносящих деревьях (контрольные деревья выключены).Смоделированные пропорции FI (т. Е. Пропорции меристем, индуцированных цветком) сравнивали с наблюдаемыми пропорциями FI в различных частях деревьев (лиственных или нелистовых, плодоносящих или неплодоносящих). Смоделированные пропорции были рассчитаны как средняя вероятность FI каждой почки в различных частях деревьев. Значения параметров моделирования, отображающие наименьшую ошибку между моделируемым и наблюдаемым FI для всех условий (дерево и локальные обработки), были выбраны как лучшее решение в процедуре калибровки.Выполнено два шага процедуры калибровки. Первый шаг заключался в изучении широкого диапазона значений путем изменения r + и r — от 0 до 15 (шаг = 0,1), t + , t — от 0 до 1 (шаг = 0,1) и v + и v — между 0,01, 0,1, 0,3 и 0,6 для активации и запрещения сигналов соответственно (всего 6 644 моделирования).На втором этапе диапазон значений, близких к лучшим решениям, был сужен для уточнения оценок (1155 симуляций).
Проверка модели проводилась в два этапа. Во-первых, два набора значений параметров для сигналов активации и запрещения, полученные в результате калибровки, были использованы для моделирования вероятности FI на плодовых деревьях, подвергшихся удалению листьев.