Аллостерические взаимодействия
Аллостерические взаимодействия проявляются не только в Б-об-разном характере кривых зависимости скорости ферментативной реакции от концентрации субстрата или аллостерического эффектора. Можно указать еще 2 типа эффектов, имеющих аллостериче-скую природу. В некоторых случаях наблюдается парадоксальный эффект активации фермента малыми концентрациями аналога кофермента или субстрата.[ …]
Аллостерические взаимодействия представляют собой непрямые взаимодействия, которые опосредуются конформационными изменениями белковой молекулы.[ …]
Особый интерес представляет аллостерическая активация. Установлено, что ряд ферментов наряду с активным каталитическим центром — участком молекулы, присоединяющим субстрат, имеет еще один центр, способный взаимодействовать с каким-либо другим веществом, отличающимся по своему строению от субстрата, но способствующим такому изменению третичной структуры ферментного белка, какое сопровождается смещением взаиморасположения атомных групп активного центра, приводящим к активации фермента. Такие ферменты получили название аллостерических (от греческих слов аААо — другой, иной и атерео — пространство), связываемое вещество — аллостерического эффектора, а место связывания — аллостерического центра. В качестве эффекторов могут выступать многие вещества, содержащиеся в клетке или приносимые к ней кровью: АМФ, глюкозоб-фосфат, различные продукты обмена веществ, гормоны и пр.[ …]
Такие ферменты получили название аллостерических (от греческих слов аААо — другой, иной и атерео — пространство), связываемое вещество — аллостерического эффектора, а место связывания — аллостерического центра. В качестве эффекторов могут выступать многие вещества, содержащиеся в клетке или приносимые к ней кровью: АМФ, глюкозоб-фосфат, различные продукты обмена веществ, гормоны и пр.[ …]
Из вышеизложенного ясно, что аллостерические взаимодействия можно разбить на две большие группы: 1) аллостерические взаимодействия в недиссоциирующем ферментативном олигомере и 2) аллостерические взаимодействия, опосредованные изменением степени диссоциации ферментативного олигомера под действием субстрата или эффекторов. Следует подчеркнуть, что эти типы взаимодействий не исключают друг друга, и, возможно, в определенных случаях мы сталкиваемся сразу с обоими типами взаимодей’ ствий.[ …]
До сих пор речь шла о моделях аллостерической регуляции активности ферментов, в которых ферментативный олигомер считается недиссоциирующим. Однако регуляторные ферменты способны в определенных условиях диссоциировать на отдельные субъединицы, причем степень диссоциации, как правило, зависит от присутствия субстратов и аллостерических эффекторов. Это свойство регуляторных ферментов позволяет предложить модель, в которой аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента под действием субстрата или эффекторов. Подобные представления были впервые развиты в 1967 г. в работах Б. И. Курганова [94, 77, 95], американского ученого Frieden [96, 97] и австралийских ученых [98]. Принципиальное отличие работ Б. И. Курганова от этих работ состоит в том, что в них изложены принципы анализа диссоциирующих ферментативных систем, позволяющие выделить кинетические эффекты, связанные с изменением степени диссоциации ферментативного олигомера под действием субстрата и алло-стерческих эффекторов. Кинетическое поведение регуляторного фермента, для которого аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента, должно зависеть от концентрации фермента.
Однако регуляторные ферменты способны в определенных условиях диссоциировать на отдельные субъединицы, причем степень диссоциации, как правило, зависит от присутствия субстратов и аллостерических эффекторов. Это свойство регуляторных ферментов позволяет предложить модель, в которой аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента под действием субстрата или эффекторов. Подобные представления были впервые развиты в 1967 г. в работах Б. И. Курганова [94, 77, 95], американского ученого Frieden [96, 97] и австралийских ученых [98]. Принципиальное отличие работ Б. И. Курганова от этих работ состоит в том, что в них изложены принципы анализа диссоциирующих ферментативных систем, позволяющие выделить кинетические эффекты, связанные с изменением степени диссоциации ферментативного олигомера под действием субстрата и алло-стерческих эффекторов. Кинетическое поведение регуляторного фермента, для которого аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента, должно зависеть от концентрации фермента. [ …]
[ …]
Возможно, что репрессор является «аллостерическим» белком с двумя активными центрами, один из которых способен реагировать с оператором, а второй—с молекулой индуктора или ингибитора. Было высказано предположение, что вследствие взаимодействия репрессора с индуктором может изменяться его конформация, так что ом теряет способность реагировать с оператором, который при этом «дерепрессируется».[ …]
Таким образом, кинетические методы позволяют обнаружить аллостерические взаимодействия в диссоциирующих ферментативных системах и проверить выполнимость модели Monod, Wyman, Changeux. Заметим, что, по мнению Teipel и Koshland [46], кривые зависимости v от [So] с двумя точками перегиба свидетельствуют о выполнимости в подобных случаях модели, развиваемой Koshland. Однако этот критерий не позволяет однозначно выбрать модель регуляторного фермента, поскольку, как было показано ранее Б. И. Кургановым, для диссоциирующего регуляторного фермента при определенных значениях кинетических параметров зависимость v от [S]o также может иметь две точки перегиба [77]. [ …]
[ …]
Предложен ряд моделей регуляторных механизмов, позволяющих количественно описать взаимодействия лигандов, связывающихся на пространственно разделенных центрах ферментов. Наиболее разработанными из них являются модель Monod, Wyman, Changeux, предложенная в 1965 г. [83], и модель Koshland, Nemethy, Filmer, предложенная в 1966 г. [84]. Модель Monod и др. постулирует, что аллостерический фермент представляет собой симметричный олигомер, построенный из идентичных субъединиц (протомеров). Предполагается, что при аллостерическом переходе все протомеры одновременно меняют свою конформацию, так что симметрия олигомера сохраняется. В этой модели аллостерические взаимодействия опосредуются смещением равновесия между, по меньшей мере, двумя конформацион-ными состояниями (R T) под действием субстратов или аллостерических эффекторов. Привлекательность модели Monod объясняется ее относительной простотой и тем, что она предсказывает определенный характер влияния эффекторов на кооперативность связывания субстрата и влияния субстрата на кооперативность связывания эффекторов. Модель Koshland и др. является более сложной и учитывает возникновение промежуточных несимметричных состояний, в которых конформации протомеров неодинаковы.[ …]
Модель Koshland и др. является более сложной и учитывает возникновение промежуточных несимметричных состояний, в которых конформации протомеров неодинаковы.[ …]
Все известные в настоящее время регуляторные ферменты построены из субъединиц и содержат несколько активных и аллостерических центров в молекуле. Кооперативная кинетика, т. е. S-образные кривые зависимости скорости реакции, катализируемой регуляторным ферментом, от концентрации субстрата или аллостерического эффектора, как правило, объясняется кооперативным взаимодействием активных и аллостерических центров.[ …]
Возможно также взаимодействие избыточных молекул субстрата с аллостерическими центрами ацетилхолинэстеразы.[ …]
Все это объясняет причину низкого сродства дезоксигемог-лобина к кислороду по сравнению с миоглобином или искусственно полученными отдельными цепями гемоглобина [13]. При связывании кислорода в четвертичной структуре гемоглобина происходит нарушение взаимодействий, ответственных за пониженное сродство к кислороду. Солевые мостики разрываются, и гидрофобные поверхности обнажаются. Становится понятным и механизм действия аллостерических эффекторов: органические фосфаты прочно связываются с определенным центром в Т-состоянии и затрудняют его переход в R-состояние.
Солевые мостики разрываются, и гидрофобные поверхности обнажаются. Становится понятным и механизм действия аллостерических эффекторов: органические фосфаты прочно связываются с определенным центром в Т-состоянии и затрудняют его переход в R-состояние.
Аллостерические ферменты
Аллостерические взаимодействия проявляются не только в Б-об-разном характере кривых зависимости скорости ферментативной реакции от концентрации субстрата или аллостерического эффектора. Можно указать еще 2 типа эффектов, имеющих аллостериче-скую природу. В некоторых случаях наблюдается парадоксальный эффект активации фермента малыми концентрациями аналога кофермента или субстрата.[ …]
Аллостерические ферменты — регуляторные ферменты, для которых характерно изменение каталитической активности в процессе нековалентного связывания метаболита в участке, не являющемся каталитическим центром.[ …]
Благодаря аллостерическим изменениям фермент может потерять активность или приобрести ее. Изменепие конфигурации фермента происходит под влиянием веществ разного происхождения.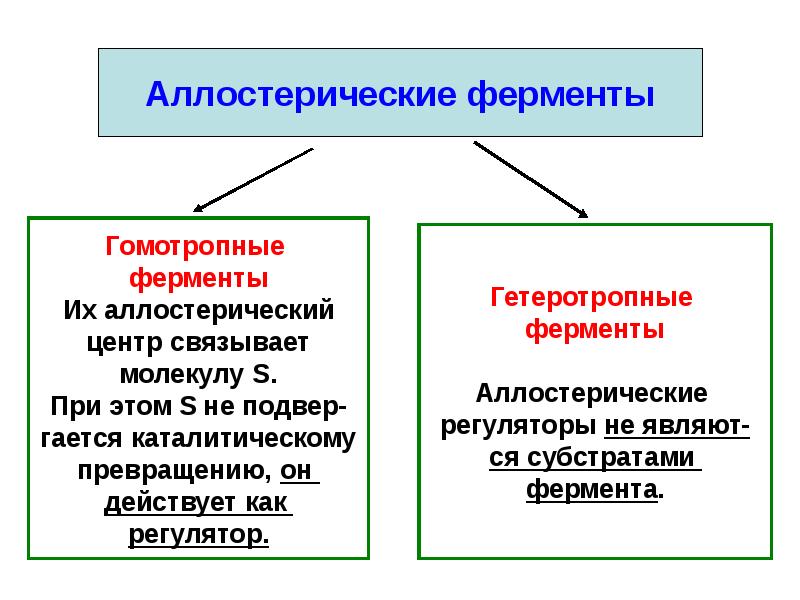 В частности, это может быть конечный продукт данной реакции, который, накапливаясь, вызывает ее замедление.[ …]
В частности, это может быть конечный продукт данной реакции, который, накапливаясь, вызывает ее замедление.[ …]
М.: Наука, 1978.[ …]
Ингибирующим может быть и аллостерический эффект, подобный аллостерической активации, с той лишь разницей, что с присоединением к ферменту аллостериче-ского ингибитора активный центр фермента изменяется так, что активность фермента понижается или утрачивается.[ …]
На примере гликолитических ферментов Э. Фёршт показывает, что эволюция ферментов происходила в сторону увеличения отношения /¡са /См (для эволюционно совершенного фермента оно составляет 108 — 109М-1-с-1) и достижения такого значения /См, которое численно превышает физиологическую концентрацию субстрата. Исключение составляют аллостерические ферменты, занимающие ключевые позиции в метаболических путях и выполняющие важные функции в регуляции скорости процессов метаболизма. Таким образом, анализ каталитической эффективности ферментов следует проводить дифференцированно, с учетом функций, выполняемых ими в живой клетке.![]() [ …]
[ …]
Особый интерес представляет аллостерическая активация. Установлено, что ряд ферментов наряду с активным каталитическим центром — участком молекулы, присоединяющим субстрат, имеет еще один центр, способный взаимодействовать с каким-либо другим веществом, отличающимся по своему строению от субстрата, но способствующим такому изменению третичной структуры ферментного белка, какое сопровождается смещением взаиморасположения атомных групп активного центра, приводящим к активации фермента. Такие ферменты получили название аллостерических (от греческих слов аААо — другой, иной и атерео — пространство), связываемое вещество — аллостерического эффектора, а место связывания — аллостерического центра. В качестве эффекторов могут выступать многие вещества, содержащиеся в клетке или приносимые к ней кровью: АМФ, глюкозоб-фосфат, различные продукты обмена веществ, гормоны и пр.[ …]
До сих пор речь шла о моделях аллостерической регуляции активности ферментов, в которых ферментативный олигомер считается недиссоциирующим. Однако регуляторные ферменты способны в определенных условиях диссоциировать на отдельные субъединицы, причем степень диссоциации, как правило, зависит от присутствия субстратов и аллостерических эффекторов. Это свойство регуляторных ферментов позволяет предложить модель, в которой аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента под действием субстрата или эффекторов. Подобные представления были впервые развиты в 1967 г. в работах Б. И. Курганова [94, 77, 95], американского ученого Frieden [96, 97] и австралийских ученых [98]. Принципиальное отличие работ Б. И. Курганова от этих работ состоит в том, что в них изложены принципы анализа диссоциирующих ферментативных систем, позволяющие выделить кинетические эффекты, связанные с изменением степени диссоциации ферментативного олигомера под дейс
Однако регуляторные ферменты способны в определенных условиях диссоциировать на отдельные субъединицы, причем степень диссоциации, как правило, зависит от присутствия субстратов и аллостерических эффекторов. Это свойство регуляторных ферментов позволяет предложить модель, в которой аллостерические взаимодействия опосредуются смещением равновесия между олигомерными формами фермента под действием субстрата или эффекторов. Подобные представления были впервые развиты в 1967 г. в работах Б. И. Курганова [94, 77, 95], американского ученого Frieden [96, 97] и австралийских ученых [98]. Принципиальное отличие работ Б. И. Курганова от этих работ состоит в том, что в них изложены принципы анализа диссоциирующих ферментативных систем, позволяющие выделить кинетические эффекты, связанные с изменением степени диссоциации ферментативного олигомера под дейс
Фармакология стр.27 | Фармакология
Сродство вещества к рецептору, приводящее к образованию с ним комплекса, обозначается термином аффинитет ‘.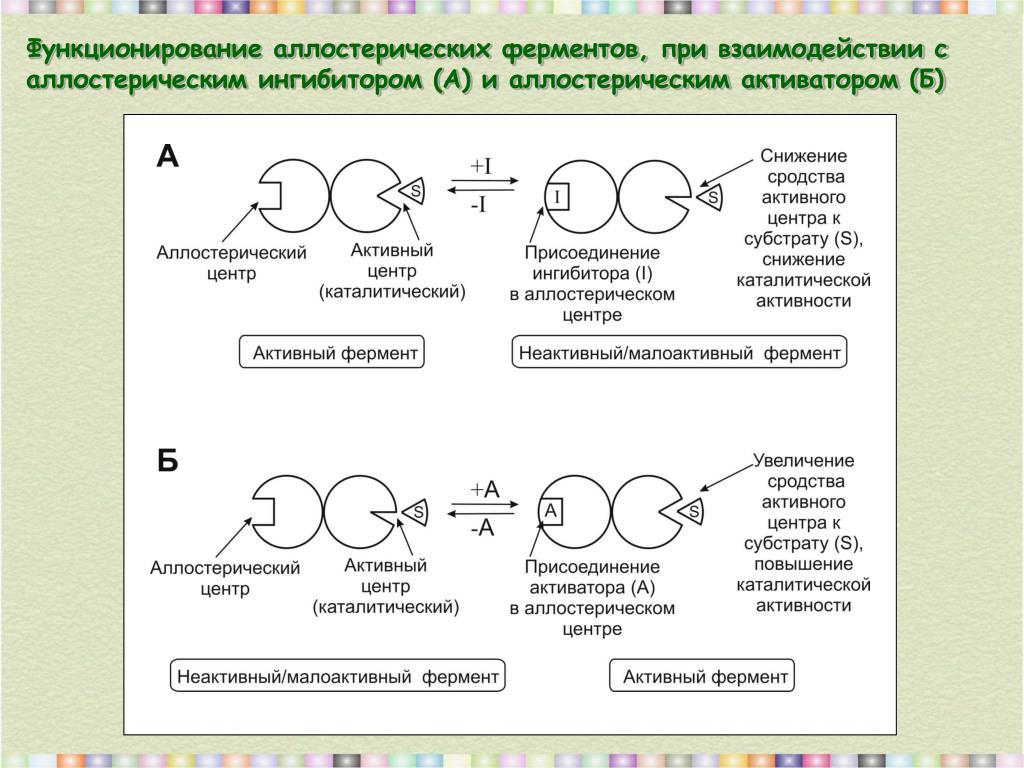 Способность вещества при взаимодействии с рецептором вызывать тог или иной эффект называется внутренней активностью.
Способность вещества при взаимодействии с рецептором вызывать тог или иной эффект называется внутренней активностью.
Вещества, которые при взаимодействии со специфическими рецепторами вызывают в них изменения, приводящие к биологическому эффекту, называют агонистами2 (они и обладают внутренней активностью). Чаще всего это стимулирующее влияние, реже—угнетающее. Если агонист, взаимодействуя с рецепторами, вызывает максимальный эффект, его называют полным агонистом. В отличие от последнего частичные агонисты при взаимодействии с теми же рецепторами не вызывают максимального эффекта.
Возможно связывание двух агонистов с разными участками макромолекулы рецептора. Это так называемое аллостерическое взаимодействие . При этом одно вещество может повышать или снижать аффинитет второго вещества к рецептору. Так, например, анксиолитик сибазон аллосге-рически повышает аффинитет ГАМК к соответствующим рецепторам (см. главу 11.4; рис. 11.2).
Вещества, не вызывающие эффекта при взаимодействии с рецепторами, но уменьшающие или устраняющие эффекты агонистов, называют антагонистами4 (их внутренняя активность равна 0).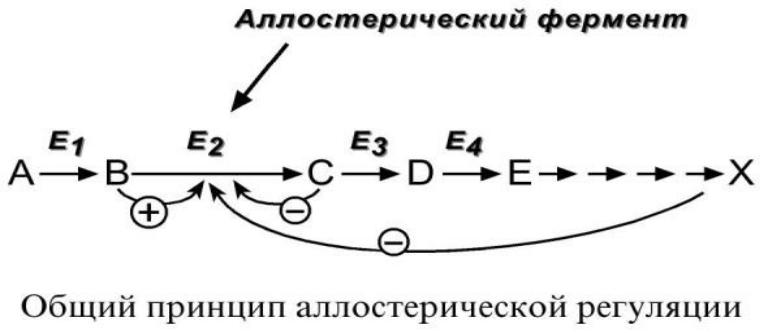 Если они занимают те же рецепторы, с которыми взаимодействуют агонисты, то речь идет о конкурентных антагонистах, если другие участки макромолекулы, не относящиеся к специфическому рецептору, но взаимосвязанные с ним, го—о неконкурентных антагонистах. Если одно и то же соединение обладает одновременно свойствами агониста и антагониста (т. е. вызывает эффект, но устраняет действие другого агониста), то его обозначают агонистом-антагонистом.
Если они занимают те же рецепторы, с которыми взаимодействуют агонисты, то речь идет о конкурентных антагонистах, если другие участки макромолекулы, не относящиеся к специфическому рецептору, но взаимосвязанные с ним, го—о неконкурентных антагонистах. Если одно и то же соединение обладает одновременно свойствами агониста и антагониста (т. е. вызывает эффект, но устраняет действие другого агониста), то его обозначают агонистом-антагонистом.
Вместе с тем существуют и так называемые неспецифические рецепторы, не связанные функционально со специфическими. К ним можно отнести белки плазмы крови, мукополисахариды соединительной ткани и др., с которыми вещества связываются, не вызывая никаких эффектов. Такие рецепторы иногда называют «молчащими» или обозначают как «места потери» веществ. Однако рецепторами целесообразно называть только специфические рецепторы; неспецифические рецепторы удобнее обозначать как «места неспецифического связывания». Взаимодействие «вещество — рецептор» осуществляется за счет межмолекулярных связей. Один из наиболее прочных видов связи—ковалентная связь. Она известна для небольшого числа препаратов (пенициллин, некоторые противобластомные вещества). Менее стойкой является пространственная ионная связь, осуществляемая за счет электростатического взаимодействия веществ с рецепторами. Последняя типична для ганглиоблокаторов, курареподобных средств, ацетилхолина. Важную роль играют ван-дер-ваальсовы силы, составляющие основу гидрофобных взаимодействий, а также водородные связи (табл. II. 1).
Один из наиболее прочных видов связи—ковалентная связь. Она известна для небольшого числа препаратов (пенициллин, некоторые противобластомные вещества). Менее стойкой является пространственная ионная связь, осуществляемая за счет электростатического взаимодействия веществ с рецепторами. Последняя типична для ганглиоблокаторов, курареподобных средств, ацетилхолина. Важную роль играют ван-дер-ваальсовы силы, составляющие основу гидрофобных взаимодействий, а также водородные связи (табл. II. 1).
’ От лат. affinis — родственный.
2 От греч. agonistes—соперник (agon — борьба).
3 От греч. alios — иной, другой; stereos — пространственный.
4 От греч. antagonisma — борьба, соперничество (anti — против, agon — борьба).
Типы взаимодействия | Примерная прочность СВЯЗИ | Уменьшение прочности связи в зависимости от расстояния между атомами, г | |
| ккал/моль | кдж/моль | ||
| Ковалентные | 50—100 | 210—420 | |
| Электростатические | г-2 | ||
| (ионные) | |||
| Ион-дипольные | 2—5 | 8—21 | г-3 |
| Диполь-дипольные | 1—3 | 4—12 | г-4 |
| Водородные связи | 2—5 | 8—21 | г-4 |
| Ван-дер-ваальсовы | 0,5 | г-7 | |
| (дисперсионные) | |||
| Гидрофобные1 | |||
1 Имеется в виду взаимодействие неполярных молекул в водной среде.
⇐ вернуться к прочитанному | | перейти на следующую страницу ⇒
Лекция 5 ФАРМАКОДИНАМИКА — Med24info.com
Фармакодинамика состоит из первичной и вторичной фармакологических реакций. Первичная фармакологическая реакция представляет собой взаимодействие с циторецепторами — биомакромолекулами, генетически детерминированными для взаимодействия с биологически активными веществами, включая лекарственные средства.
Взаимодействие с циторецепторами необходимо для развития вторичной фармакологической реакции в виде изменений метаболизма и функций клеток и органов. Нерецепторные механизмы действия встречаются редко. Отсутствуют циторецепторы для ингаляционных наркозных, осмотических мочегонных средств, плазмозаменителей, комплексонообразователей.
Одна и та же первичная фармакологическая реакция может приводить к различным вторичным изменениям (возбуждение а1-адренорецепторов норадреналином вызывает расширение зрачков и сужение сосудов). В основе вторичной фармакологической реакции иногда лежат различные первичные механизмы (спазм бронхов возникает при возбуждении М-холинорецепторов ацетилхолином или #грецепторов гистамином).
ВЗАИМОДЕЙСТВИЕ ЛЕКАРСТВЕННЫХ СРЕДСТВ С ЦИТОРЕЦЕПТОРАМИ
Циторецепторы (греч. kytos — сосуд, клетка, лат. recipere — получать) созданы природой для эндогенных лигандов — гормонов, факторов роста, нейромедиаторов, аутакоидов. Они имеют структуру липопротеинов, гликопротеинов, металлопротеинов, нуклеопротеинов. Реакция рецепторов на ксенобиотики обусловлена низкой специфичностью взаимодействия. Как правило, ксенобиотики обладают такой же, как и биологически активные вещества организма, стереохимической композицией.
Концепция циторецепторов была предложена Паулем Эрлихом и Джоном Лэнгли в начале XX в. По мнению Эрлиха, функцию рецепторов выполняют молекулярные боковые цепи клеточной протоплазмы, способные воспринимать питательные вещества и токсины. Боковые цепи, циркулирующие в крови, образуют антитоксины (антитела).Рецепторами являются также «крайние образования» чувствительных клеток. Они превращают физическую или химическую энергию раздражения в возбуждение чувствительных нервов.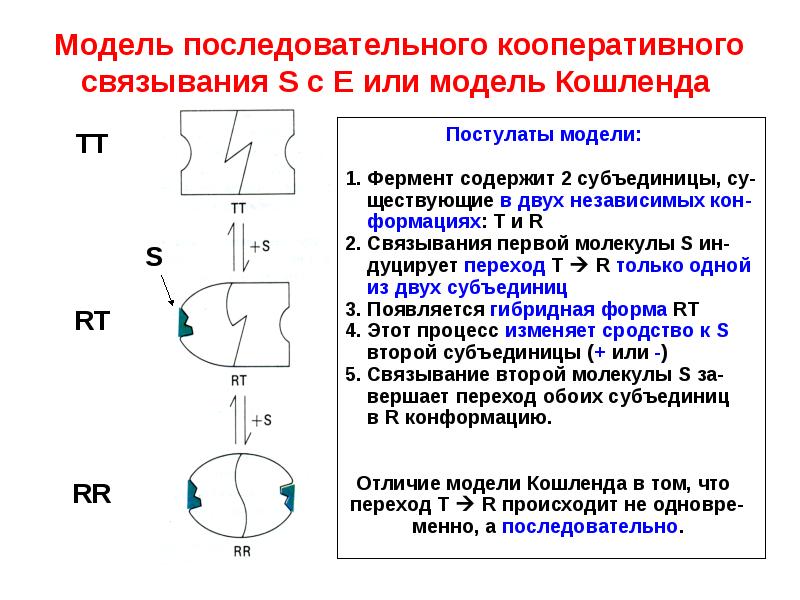 Лэнгли установил, что кураре устраняет сокращение скелетных мышц, вызываемое никотином, но не изменяет возбуждающий эффект электрического тока.
Лэнгли установил, что кураре устраняет сокращение скелетных мышц, вызываемое никотином, но не изменяет возбуждающий эффект электрического тока.
В структуре циторецепторов присутствуют домен для связывания лигандов и эффекторный домен. Активные центры циторецепторов образованы функциональными группами аминокислот, фосфатидов, нуклеотидов, сахаров.
Лекарственные средства устанавливают с циторецепторами непрочные физико-химические связи
- вандерваальсовы, ионные, водородные, дипольные по принципу комплементарности (активные группы лекарств взаимодействуют с соответствующими группами активного центра циторецепторов).
Необратимые ковалентные связи с циторецепторами образуют немногие вещества — необратимые ингибиторы холинэстеразы, тяжелые металлы, цитостатики. Все они высокотоксичны.
По отношению к циторецепторам лекарственные средства обладают аффинитетом (лат. affinis — родственный) и внутренней активностью. Аффинитет (сродство) рассматривают как способность образовывать комплекс с циторецепторами. Внутренняя активность направлена на создание их активной стереоконформации, приводящей к появлению клеточного ответа. В зависимости от выраженности аффинитета и наличия внутренней активности лекарственные средства разделяют на 2 группы:
Внутренняя активность направлена на создание их активной стереоконформации, приводящей к появлению клеточного ответа. В зависимости от выраженности аффинитета и наличия внутренней активности лекарственные средства разделяют на 2 группы:
- агонисты (греч. agonistes — соперник, agon — борьба), или миметики (греч. mimeomai — подражать) — вещества с умеренным аффинитетом и высокой внутренней активностью: полные агонисты вызывают максимально возможный клеточный ответ, частичные (парциальные) агонисты — менее значительную клеточную реакцию;
- антагонисты (греч. antagonisma — соперничество, anti — против, agon — борьба) или блокаторы
- вещества с высоким аффинитетом, но лишенные внутренней активности. Они экранируют циторецепторы от действия эндогенных лигандов и препятствуют развитию клеточного ответа, усиливая эффекты других, неблокированных циторецепторов. Вещества, блокирующие активные центры циторецепторов, являются конкурентными антагонистами.


Возможно сочетание в фармакодинамике свойств агониста и антагониста, например, агонисты- антагонисты возбуждают одни циторецепторы и блокируют другие.
Циторецепторы возбуждаются только в начальный момент взаимодействия с лекарственными средствами. Для агонистов характерна высокая константа диссоциации комплекса лекарство — циторецептор. Антагонисты, обладая высоким аффинитетом, более продолжительно связываются с циторецепторами, при этом в первый момент взаимодействия возможно появление эффекта возбуждения.
Лекарственные средства присоединяются также к аллостерическим центрам циторецепторов, что модифицирует структуру активных центров и изменяет их реакцию на лекарства или эндогенные лиганды. Пример аллостерических рецепторов — бензодиазепиновые рецепторы, повышающие аффинитет ГАМК-рецепторов типа А.
На мембране одной и той же клетки может присутствовать более 10 типов циторецепторов с различным функциональным значением, поэтому клеточный ответ на возбуждение циторецепторов является алгебраической суммой вызываемых ими независимых реакций.
Циторецепторы классифицируют на 4 типа (рис. 3):
- рецепторы-протеинкиназы;
- рецепторы ионных каналов;
- рецепторы, ассоциированные с G-белками;
- рецепторы-регуляторы транскрипции.
Циторецепторы-протеинкиназы связаны с плазматической мембраной клеток. Они имеют внеклеточный домен для взаимодействия с лигандами и внутриклеточный каталитический домен — протеинкиназу. Оба домена соединены внутримембранной цепью из гидрофобных аминокислот. Рецепторы-протеинкиназы фосфорилируют белки клеток — киназы, регуляторные и структурные белки. Чаще всего происходит фосфорилирование тирозина белков-мишеней. Примеры циторецепторов-протеинкиназ — рецепторы инсулина, цитокинов, эпидермального и тромбоцитарного факторов роста.
Циторецепторы к предсердному (атриальному) натрийуретическому пептиду, ассоциированные с гуанилатциклазой, повышают продукцию цГМФ.
- рецепторы, вызывающие гиперполяризацию и торможение функций клеток, — ГАМКА-рецепторы и рецепторы глицина (увеличивают проводимость для СГ).

Циторецепторы, ассоциированные с G-белками, представляют собой группу интегральных мембранных белков с характерной вторичной структурой, включающей внеклеточный N-конец и внутриклеточный С-конец, 7 трансмембранных доменов, связанных тремя внеклеточными и тремя внутриклеточными петлями. Белковая цепь циторецепторов содержит большое количество остатков цистеина, образующих дисульфидные мостики. Внеклеточные петли, выполняя регуляторную функцию, ответственны за распознавание и связывание лигандов. G-белки, расположенные внутри мембраны, передают информацию от внеклеточного регуляторного домена на эффекторную систему, используя энергию ГТФ. Внеклеточные и трансмембранные домены согласованно участвуют в связывании лигандов и активации циторецепторов.
Эффекторная система представлена аденилатциклазой, фосфолипазами А2, С и D, белками ионных каналов, транспортными белками. При возбуждении рецепторов образуются внутриклеточные биологически активные вещества — вторичные мессенджеры (англ.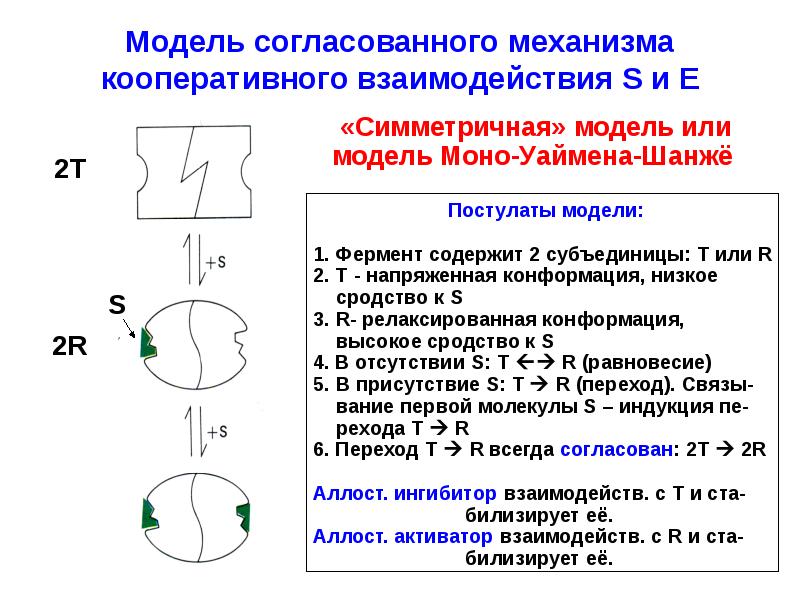 -рецепторы вазопрессина;
-рецепторы вазопрессина;
Рецепторы ингибирования: а2-адренорецепторы; М2 4-холинорецепторы;
D2-рецепторы дофамина; А1 -рецепторы аденозина;
5-HT1-рецепторы серотонина; опиоидные рецепторы.
Наибольшее значение имеют следующие эффекты цАМФ:
- активация протеинкиназ, катализирующих фосфорилирование ферментов и структурных белков клеток;
- транспорт ионов кальция в нервные окончания, клетки желез, миокард, скелетные мышцы, тромбоциты;
- депонирование ионов кальция в гладких мышцах.
Фосфолипаза С катализирует гидролиз фосфатидилинозитолдифосфата. Этот фосфолипид клеточных мембран представляет собой эфир шестиатомного спирта инозитола с диацилглицеролом, имеющим два остатка жирных кислот (одна из них — арахидоновая кислота). Продукты реакции — вторичные мессенджеры инозитолтрифосфат и диацилглицерол. Одна молекула инозитолтрифосфата освобождает 20 ионов кальция из эндоплазматического ретикулума. Диацилглицерол, активируя протеинкиназу С, освобождает нейромедиаторы, гормоны, секреты экзокринных желез, стимулирует рост и деление клеток. Метаболит диацилглицерола — арахидоновая кислота становится источником для синтеза простагландинов, тромбоксанов, простациклина, лейкотриенов.
Диацилглицерол, активируя протеинкиназу С, освобождает нейромедиаторы, гормоны, секреты экзокринных желез, стимулирует рост и деление клеток. Метаболит диацилглицерола — арахидоновая кислота становится источником для синтеза простагландинов, тромбоксанов, простациклина, лейкотриенов.
Циторецепторы, связанные с фосфолипазой С:
а1-адренорецепторы; Р2у -пуриновые рецепторы;
5-HT2 -рецепторы серотонина; Кгрецепторы вазопрессина;
М1,3-холинорецепторы; рецепторы окситоцина;
h2 -рецепторы гистамина; рецепторы холецистокинина.
Циторецепторы-регуляторы транскрипции взаимодействуют с тиреоидными, стероидными гормонами, витамином D и ретиноидами. Транспортные белки крови передают лиганды клеточным белкам, затем комплексы поступают в ядро. Функции рецепторов — активация или ингибирование транскрипции генов.
Рецепторную функцию выполняют также мембраносвязанные и растворимые ферменты (дигидрофолатредуктаза, ацетилхолинэстераза, моноаминоксидаза, циклоксигеназа), транспортные белки (Na+, К+-АТФ-аза) и структурные белки (тубулин).
ВИДЫ ДЕЙСТВИЯ ЛЕКАРСТВЕННЫХ СРЕДСТВ
Местное и резорбтивное действие
Местное действие — эффекты лекарственных средств на месте применения (потеря болевой и температурной чувствительности под влиянием местных анестетиков; боль, гиперемия, отек кожи в области нанесения раздражающих препаратов).
Резорбтивное действие (лат. resorbeo — поглощаю) — эффекты лекарственных средств после всасывания в кровь и проникновения через гистогематические барьеры (анальгезия при применении наркозных средств, наркотических и ненаркотических анальгетиков; повышение умственной и физической работоспособности у людей, принимающих кофеин).
Прямое и косвенное действие
Прямое (первичное) действие — изменение лекарственными средствами функций органов в результате действия на клетки этих органов (сердечные гликозиды усиливают сердечные сокращения, блокируя Na+, К -АТФ-азу мышечных клеток миокарда; мочегонные средства повышают диурез, нарушая реабсорбцию ионов и воды в почечных канальцах). -повышают фильтрацию и образование мочи).
-повышают фильтрацию и образование мочи).
Частным случаем косвенного действия является рефлекторное — изменение функций органов за счет прямой стимуляции чувствительных нервных окончаний. Деполяризация нервных окончаний вызывает импульс, который по рефлекторным дугам при участии нервных центров передается на исполнительные органы. Рефлекторными эффектами в результате возбуждения экстерорецепторов обладают кожные раздражители; интерорецепторов — отхаркивающие, рвотные, желчегонные, слабительные средства; хеморецепторов сосудов — аналептики, проприорецепторов скелетных мышц — миорелаксанты.
Обратимое и необратимое действие
Обратимое действие обусловлено установлением непрочных физико-химических связей с циторецепторами, характерно для большинства лекарственных средств.
Необратимое действие возникает в результате образования ковалентных связей с циторецепторами, характерно для немногих лекарственных средств, как правило, обладающих высокой токсичностью и применяемых местно.
Главное и побочное действие
Главное действие — терапевтические эффекты лекарственных средств.
Побочное действие — дополнительные, нежелательные эффекты.
Фармакологические эффекты одного и того же лекарственного средства могут оказаться главными или побочными при различных заболеваниях. Так, при лечении бронхиальной астмы главное действие адреналина — расширение бронхов, при гипогликемической коме — усиление гликогенолиза и повышение содержания глюкозы в крови.
Побочные реакции наблюдаются при приеме многих лекарственных средств. Частота их при амбулаторном лечении достигает 10 — 20 %, а 0,5 — 5 % больных нуждаются в госпитализации из-за осложнений фармакотерапии.
Избирательное (элективное) действие
Избирательное действие — влияние лекарственных средств на функции только определенных органов и систем. Оно обусловлено в большей степени избирательным связыванием с циторецепторами, в меньшей степени — избирательным накоплением в органах и тканях, хотя известны примеры создания лекарствами высоких концентраций в клетках, на которые они оказывают действие.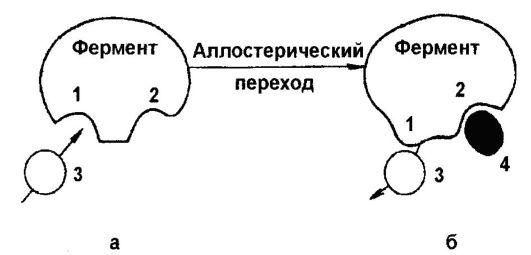 Магния сульфат, не всасываясь из кишечника, усиливает перистальтику и
Магния сульфат, не всасываясь из кишечника, усиливает перистальтику и
вызывает желчегонный эффект. При парентеральном введении ионы магния угнетают ЦНС. Ингаляционные наркозные средства создают в головном мозге концентрацию, в 1,5 — 2 раза более высокую, чем в крови. Йод интенсивно поступает только в щитовидную железу.
ФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ, ВЫЗЫВАЕМЫЕ ЛЕКАРСТВЕННЫМИ СРЕДСТВАМИ В ОРГАНИЗМЕ
Возбуждение — повышение функции выше нормы: положительное возбуждение — сокращение миометрия под влиянием препаратов спорыньи, примененных в большой дозе для остановки маточного кровотечения; отрицательное возбуждение — судороги при отравлении стрихнином, камфорой.
Успокоение — возврат возбужденной функции к норме: снижение температуры тела жаропонижающими средствами при лихорадке, уменьшение боли под влиянием анальгетиков.
Угнетение — снижение функции ниже нормы: положительное угнетение — утрата сознания, подавление рефлексов и снижение тонуса скелетных мышц при наркозе; отрицательное угнетение — торможение дыхательного центра при действии наркозных, снотворных средств, наркотических анальгетиков.
Тонизирование — возврат угнетенной функции к норме: улучшение деятельности дыхательного и сосудодвигательного центров при дыхательной недостаточности, коллапсе, шоке под влиянием камфоры, кофеина и других аналептиков; усиление сокращений декомпенсированного миокарда у больных, получающих сердечные гликозиды.
Паралич — прекращение функции: положительный паралич — обратимое устранение болевой чувствительности при действии сильных местных анестетиков, полное расслабление скелетной мускулатуры, вызываемое миорелаксантами; отрицательный паралич — необратимый паралич дыхательного центра после его возбуждения атропином и аналептиками в токсических дозах.
Аллостерические взаимодействия — Справочник химика 21
Аллостерические взаимодействия проявляются в характере кривых зависимости начальной скорости реакции от концентрации субстрата или эффектора, в частности в -образности этих кривых (отклонение от гиперболической кривой Михаэлиса-Ментен). 8-образный характер зависимости V от [8] в присутствии модулятора обусловлен эффектом кооперативности. Это означает, что связывание одной молекулы субстрата облегчает связывание второй молекулы в активном центре, способствуя тем самым увеличению скорости реакции. Кроме того, для аллостерических регуляторных ферментов характерна нелинейная зависимость скорости реакции от концентрации субстрата. [c.156]
8-образный характер зависимости V от [8] в присутствии модулятора обусловлен эффектом кооперативности. Это означает, что связывание одной молекулы субстрата облегчает связывание второй молекулы в активном центре, способствуя тем самым увеличению скорости реакции. Кроме того, для аллостерических регуляторных ферментов характерна нелинейная зависимость скорости реакции от концентрации субстрата. [c.156]
Системы с аллостерическим взаимодействием, [c.251]
Аллостерические взаимодействия в диссоциирующих ферментных системах [c.479]
КООПЕРАТИВНОЕ СВЯЗЫВАНИЕ ЛИГАНДОВ И АЛЛОСТЕРИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ [c.252]
Детали механизма связывания кислорода гемоглобином сложны, но хорошо изучены. Будучи чрезвычайно важным с физиологической точки зрения, этот процесс служит прекрасной иллюстрацией аллостерических взаимодействий и регуляции. Характерные особенности связывания кислорода гемоглобином могут быть суммированы следующим образом [c.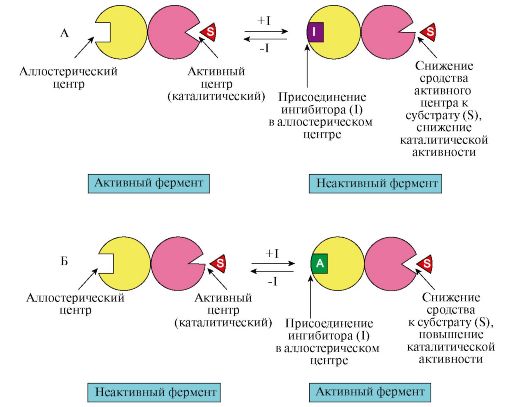 170]
170]
Помимо адениловых нуклеотидов в регулировании энергетических процессов активную роль играют система НАД(Ф)» / /НАД(Ф) Н2-коферментов и величина трансмембранного электрохимического градиента ионов водорода в виде обоих его составляющих и АрН). Преобладание аллостерического взаимодействия восстановленной или окисленной форм НАД(Ф) с ферментами катаболического пути приводит соответственно к понижению или повышению их активности. Достижение определенного порогового значения Арн+ на энергопреобразующей мембране служит определенным сигналом, тормозящим поступление ионов водорода против градиента. [c.124]
Способность связывать спиновые зонды может изменяться в зависимости от состояния белка, поэтому степень связывания спиновых зондов можно использовать для изучения конформационных превращений белковых макромолекул. Так, например, с помощью спин-меченых трифосфатов (таких, например, как радикал AXV), обратимо связывающихся с молекулами гемоглобина, удалось исследовать аллостерические взаимодействия в этом белке [191, 194]. [c.192]
[c.192]
Метаболит может ингибировать или активировать фермент по пути конкурентного или аллостерического взаимодействия. Регулирование биосинтеза может происходить как по типу отрицательной, так и по типу положительной обратной связи. [c.434]
Ферменты, изменяющие активность за счет аллостерического взаимодействия белка-фермента с эффектором или продуктом реакции, как правило, катализируют начальную стадию последовательных реакций, и их называют регуляторными ферментами. Этим белкам приписывается основная роль в регуляции ферментативной активности и в регуляции синтеза специфических белков. [c.436]
Итак, современные представления связывают образование вторичной и третичной структуры глобулярных белков с той информацией, которую несет первичная структура белковых цепей в момент биосинтеза белка в клетке Доказательством суш,ествования предпочтительных трехмерных структур является также то, что синтетические полипептиды и белки проявляют биологическую активность (например, АКТГ, инсулин, рибонуклеаза).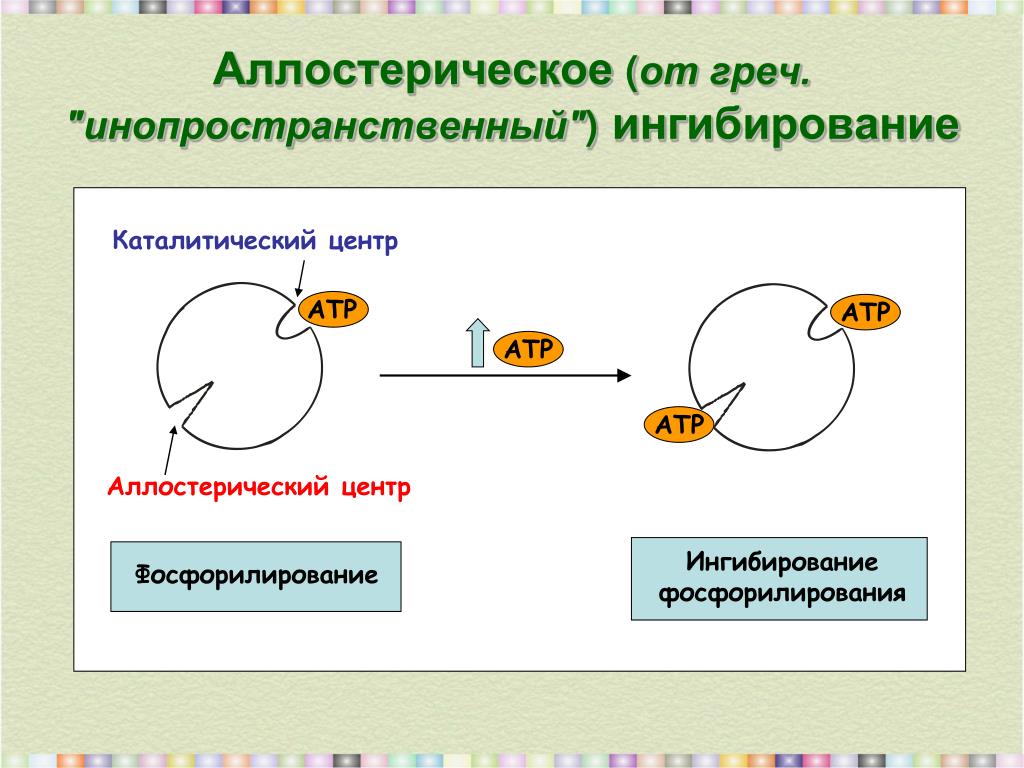 Но, принимая это положение за основу, нельзя забывать, что в физиологических условиях в процессе выполнения биологических функций могут происходить динамичные обратимые сдвиги в конформации глобулярных белков. Эти сдвиги могут явиться, например, результатом так называемых аллостерических взаимодействий в молекулах ферментов (см. главу Ферменты ). Такая способность к обратимой изменчивости тесно связана с регуляцией активности ферментов, с регуляцией процессов жизнедеятельности на клеточном уровне. [c.157]
Но, принимая это положение за основу, нельзя забывать, что в физиологических условиях в процессе выполнения биологических функций могут происходить динамичные обратимые сдвиги в конформации глобулярных белков. Эти сдвиги могут явиться, например, результатом так называемых аллостерических взаимодействий в молекулах ферментов (см. главу Ферменты ). Такая способность к обратимой изменчивости тесно связана с регуляцией активности ферментов, с регуляцией процессов жизнедеятельности на клеточном уровне. [c.157]
Наряду с ингибиторами существует ряд соединений, способствующих активации ферментативных реакций. Это — активаторы ферментов (см. Кофакторы ферментов ). Молекула активатора может вызвать такие изменения в конформации белковой молекулы фермента (аллостерическое взаимодействие), которые будут способствовать связыванию фермента субстратом, т. е. происходит активирование фермента. Наиболее распространенным активатором многих ферментов является восстановленный глутатион, который активирует фермент, восстанавливая дисульфидные группы в сульфгидрильные. [c.234]
[c.234]
Аллостерические взаимодействия проявляются не только в S-образном характере кривых зависимости скорости ферментативной реакции от концентрации субстрата или аллостерического эффектора. Можно указать еще 2 типа эффектов, имеющих аллостерическую природу. В некоторых случаях наблюдается парадоксальный эффект активации фермента малыми концентрациями аналога кофермента или субстрата. [c.101]
Аллостерические взаимодействия представляют собой непрямые взаимодействия, которые опосредуются конформационными изменениями белковой молекулы. [c.105]
Системы с аллостерическим взаимодействием, а. Аллостери-ческое ингибирование —тип ингибирования, который реализуется в ферментных цепях, если действие одного из ферментов, стоящих в начале цепи, подавляется одним или несколькими продуктами последую1цпх 4 ерментатйвных реакций. Для системы трех ферментов [c.197]
Динамическая структура белковых макромолекул ферментов, постулированная Ламри, Линдерштром-Лангом и Кошландом, которая проявляется в локальной тепловой подвижности отдельных участков и в способности к индуцированным конформационным переходам, играет первостепенную роль в реализации таких функционально важных свойств ферментов, как динамическая адаптация формы фермента к структуре каталитических и субстратных групп, меняющаяся в процессе химической реакции, аллостерическое взаимодействие между пространственно разобщенными центрами, реализация принципа компле-ментарности свободных энергий (по Ламри) и индуцированного соответствия (по Кошланду).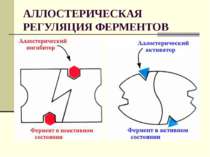
Механизмы аллостерический — Справочник химика 21
Механизм аллостерического регулирования процесса функционирования ферментов обеспечивается узнаванием и связыванием метаболита-регулятора в аллостерическом центре. В результате происходит изменение конформационного состояния и каталитических свойств активного центра белковой молекулы. Конечный этап заключается в воздействии по принципу обратной связи на источник возникновения регулирующего сигнала изменение скорости образования продуктов ферментативной реакции, поступающих в цепь последовательных метаболических реакций, приводит к изменению функционирования фермента, продуцирующего метаболит-регулятор. [c.81]
Биосинтетические пути регулируются преимущественно по механизму аллостерического ингибирования первого фермента и репрессии синтеза ферментов этого пути конечным продуктом. Регулирование разветвленных биосинтетических путей осуществляется с помощью усложненных вариантов этих же механизмов. [c.123]
Все эти вопросы уже выходят за пределы задач физической биохимии и относятся к области молекулярной кибернетики. Однако сам молекулярный механизм аллостерической регуляции активности ферментов имеет физикохимическую природу и заслуживает здесь хотя бы краткого рассмотрения. [c.243]
В результате меняется и способность субстрата связываться с ферментом (чем данное явление и отличается от неконкурентного ингибирования разд. 4.4.2). Действующие таким образом вещества назьшаются аллостерическими ингибиторами. Рис. 4.14 поясняет механизм аллостерического ингибирования. [c.164]
За последние годы было накоплено много данных, на основании которых можно предположить, что активность многих ферментов может быть ослаблена или усилена в результате образования комплекса молекулы эффектора с участком фермента, совершенно отличным от каталитически активного центра [946]. Тот факт, что субстрат и эффектор не присоединяются к одному и тому же участку фермента, можно подтвердить различными путями например, можно настолько модифицировать ферменты химически, что они утратят свою чувствительность по отношению к эффектору, хотя ферментативная активность в отсутствие эффектора остается неизмененной. Эффекторы называются аллостерическими, т. е. они не обладают каким-либо стерическим сходством с субстратом фермента, активность которого они модифицируют. Моно и др. [946] предложили механизм аллостерического ингибирования, согласно которому фермент имеет каталитические и аллостерические активные центры и образование комплекса на аллостерическом центре приводит к модификации конформации фермен- [c.328]
Б. Механизмы аллостерических взаимодействий и кооперативности [c.254]
Механизмы увеличения продукции АТФ. Многие процессы, обеспечивающие работу мышц энергией, рассмотрены в предыдущих разделах. К ним относится увеличение снабжения мышц окисляемыми субстратами мобилизация гликогена печени и мышц, глюконеогенез из молочной кислоты (цикл Кори и глюкозо-аланиновый цикл), мобилизация депонированных жиров и поступление жирных кислот и кетоновых тел в мышцы. Увеличиваются также легочная вентиляция и скорость кровотока, а следовательно, и снабжение мышц кислородом. Эти процессы вместе с механизмами аллостерической регуляции, повышающими активность ключевых ферментов катаболизма, многократно увеличивают скорость синтеза АТФ. [c.527]
В основе механизма аллостерической регуляции лежит взаимодействие пространственно раздепен-ных центров в молекуле фермента (каталитического и аллостерического). Субстрат присоединяется к каталитическому центру, а эффекторы-регуляторы — к аллостерическим центрам. [c.54]
Механизм аллостерических взаимо- [c.85]
Л 7. Согласованный механизм аллостерических взаимодействий [c.118]
В соответствии с моделью согласованного механизма аллостерический ингибитор (изо бражен в виде шестиугольника) стабилизирует форму Т, тогда как аллостерический активатор (изображен в виде треугольника) стабилизирует форму R. [c.120]
Согласованный механизм аллостерических взаимодействий 118
Аллостерическая регуляция — Infogalactic: ядро планетарного знания
В биохимии аллостерическая регуляция (или аллостерическая регуляция ) — это регуляция белка путем связывания эффекторной молекулы в сайте, отличном от активного сайта фермента.
Сайт, с которым связывается эффектор, называется аллостерическим сайтом . Аллостерические сайты позволяют эффекторам связываться с белком, что часто приводит к конформационным изменениям, связанным с динамикой белка.Эффекторы, которые усиливают активность белка, называются аллостерическими активаторами , тогда как те, которые снижают активность белка, называются аллостерическими ингибиторами .
Аллостерические правила — естественный пример контуров управления, таких как обратная связь от последующих продуктов или прямая связь от исходных субстратов. Аллостерия на большие расстояния особенно важна в передаче сигналов клетками. [1] Аллостерическая регуляция также особенно важна для способности клетки регулировать активность ферментов.
Термин аллостерия происходит от греческого allos (ἄλλος), «другой», и стерео (στερεὀς), «твердое тело (объект)». Это относится к тому факту, что регуляторный сайт аллостерического белка физически отличается от его активного сайта.
Модели аллостерической регуляции
Большинство аллостерических эффектов можно объяснить согласованной моделью MWC , предложенной Монодом, Вайманом и Ченжаксом, [2] или последовательной моделью, описанной Кошландом, Немети и Филмером. [3] Оба постулируют, что субъединицы фермента существуют в одной из двух конформаций, напряженной (Т) или расслабленной (R), и что расслабленные субъединицы связывают субстрат легче, чем субъединицы в напряженном состоянии. Две модели больше всего различаются своими предположениями о взаимодействии субъединиц и предсуществовании обоих состояний.
Концертная модель
Согласованная модель аллостерии, также называемая моделью симметрии или моделью MWC, постулирует, что субъединицы фермента связаны таким образом, что конформационные изменения в одной субъединице обязательно передаются всем другим субъединицам.Таким образом, все субъединицы должны существовать в одной и той же конформации. Модель также утверждает, что в отсутствие какого-либо лиганда (субстрата или другого) равновесие благоприятствует одному из конформационных состояний, T или R. аллостерический эффектор или лиганд) на сайт, отличный от активного сайта (аллостерический сайт).
Последовательная модель
Последовательная модель аллостерической регуляции утверждает, что субъединицы не связаны таким образом, чтобы конформационные изменения в одной вызывали аналогичные изменения в других.Таким образом, все субъединицы фермента не требуют одинаковой конформации. Более того, последовательная модель диктует, что молекулы субстрата связываются посредством протокола индуцированной подгонки. В общем, когда субъединица случайным образом сталкивается с молекулой субстрата, активный центр, по сути, образует перчатку вокруг субстрата. Хотя такая индуцированная подгонка переводит субъединицу из напряженного состояния в расслабленное состояние, она не распространяет конформационные изменения на соседние субъединицы. Вместо этого связывание с субстратом одной субъединицы лишь незначительно изменяет структуру других субъединиц, так что их сайты связывания более восприимчивы к субстрату.Подводя итог:
- субъединицы могут не существовать в одной и той же конформации
- молекул субстрата связываются по протоколу индуцированной подгонки
- конформационные изменения не распространяются на все субъединицы
Morpheein модель
Морфеиновая модель аллостерической регуляции — это диссоциативная согласованная модель. [4]
Морфеин — это гомоолигомерная структура, которая может существовать как ансамбль физиологически значимых и функционально различных альтернативных четвертичных ансамблей.Переходы между альтернативными сборками морфеина включают диссоциацию олигомеров, конформационные изменения в диссоциированном состоянии и повторную сборку с образованием другого олигомера. Требуемый этап разборки олигомера отличает модель морфеина для аллостерической регуляции от классических моделей MWC и KNF. Порфобилиногенсинтаза (PBGS) является прототипом морфеина.
Аллостерический ресурс
Аллостерическая база данных
Аллостерия — это прямое и эффективное средство регуляции функции биологических макромолекул, производимое связыванием лиганда в аллостерическом сайте, топографически отличном от ортостерического сайта.Ожидается, что из-за часто высокой селективности рецепторов и более низкой токсичности в отношении мишеней аллостерическая регуляция будет играть все большую роль в открытии лекарств и биоинженерии. База данных AlloSteric (ASD, http://mdl.shsmu.edu.cn/ASD) [5] обеспечивает центральный ресурс для отображения, поиска и анализа структуры, функции и связанных аннотаций для аллостерических молекул. В настоящее время ASD содержит аллостерические белки более 100 видов и модуляторы трех категорий (активаторы, ингибиторы и регуляторы).Каждый белок снабжен аннотацией с подробным описанием аллостерии, биологического процесса и связанных заболеваний, а каждый модулятор — сродством связывания, физико-химическими свойствами и терапевтической областью. Интеграция информации об аллостерических белках в РАС должна позволить предсказывать аллостерию для неизвестных белков с последующей экспериментальной проверкой. Кроме того, модуляторы, курируемые в ASD, могут использоваться для исследования потенциальных аллостерических мишеней для запрашиваемого соединения и могут помочь химикам реализовать модификации структуры для создания новых аллостерических лекарств.
Аллостерическая модуляция
Положительная модуляция
Положительная аллостерическая модуляция (также известная как аллостерическая активация ) происходит, когда связывание одного лиганда усиливает притяжение между молекулами субстрата и другими сайтами связывания. Примером является связывание молекул кислорода с гемоглобином, где кислород эффективно является и субстратом, и эффектором. Аллостерический или «другой» сайт является активным сайтом соседней белковой субъединицы. Связывание кислорода с одной субъединицей вызывает конформационные изменения в этой субъединице, которая взаимодействует с оставшимися активными сайтами, увеличивая их сродство к кислороду .Другой пример аллостерической активации наблюдается в цитозольной IMP-GMP специфической 5′-нуклеотидазе II (cN-II), где сродство к субстрату GMP увеличивается при связывании GTP на границе раздела димеров [6]
Отрицательная модуляция
Отрицательная аллостерическая модуляция (также известная как аллостерическое ингибирование ) происходит, когда связывание одного лиганда снижает сродство к субстрату на других активных сайтах. Например, когда 2,3-BPG связывается с аллостерическим сайтом гемоглобина, сродство к кислороду всех субъединиц снижается.Это когда в сайте связывания отсутствует регулятор.
Прямые ингибиторы тромбина являются прекрасным примером отрицательной аллостерической модуляции. Были обнаружены аллостерические ингибиторы тромбина, которые потенциально могут использоваться в качестве антикоагулянтов.
Другой пример — стрихнин, судорожный яд, который действует как аллостерический ингибитор рецептора глицина. Глицин является основным постсинаптическим тормозным нейромедиатором в спинном мозге и стволе мозга млекопитающих.Стрихнин действует на отдельном сайте связывания на рецепторе глицина аллостерическим образом; то есть его связывание снижает сродство рецептора глицина к глицину. Таким образом, стрихнин подавляет действие тормозящего медиатора, что приводит к судорогам.
Другой пример, в котором можно наблюдать отрицательную аллостерическую модуляцию, — это взаимодействие АТФ и фермента фосфофруктокиназы в петле отрицательной обратной связи, которая регулирует гликолиз. Фосфофруктокиназа (обычно называемая ПФК) — это фермент, который катализирует третью стадию гликолиза: фосфорилирование фруктозо-6-фосфата в фруктозо-1,6-бисфосфат.PFK может аллостерически подавляться высокими уровнями АТФ в клетке. Когда уровни АТФ высоки, АТФ связывается с аллосторным участком фосфофруктокиназы, вызывая изменение трехмерной формы фермента. Это изменение приводит к снижению его сродства к субстрату (фруктозо-6-фосфату и АТФ) в активном центре, и фермент считается неактивным. Это вызывает прекращение гликолиза при высоком уровне АТФ, таким образом сохраняя глюкозу в организме и поддерживая сбалансированный уровень клеточного АТФ.Таким образом, АТФ служит отрицательным аллостерическим модулятором для PFK, несмотря на то, что он также является субстратом фермента.
Типы аллостерической регуляции
Гомотропный
Гомотропный аллостерический модулятор является субстратом для своего целевого фермента, а также регулирующей молекулой активности фермента. Обычно это активатор фермента. Например, O 2 — гомотропный аллостерический модулятор гемоглобина.
Гетеротропный
Гетеротропный аллостерический модулятор — это регуляторная молекула, которая не является субстратом фермента.Это может быть активатор или ингибитор фермента. Например, H + , CO 2 и 2,3-бисфосфоглицерат являются гетеротропными аллостерическими модуляторами гемоглобина. [7]
Некоторые аллостерические белки могут регулироваться как их субстратами, так и другими молекулами. Такие белки способны к гомотропным и гетеротропным взаимодействиям.
Ненормативная аллостерия
Нерегулирующий аллостерический сайт относится к любому нерегулирующему компоненту фермента (или любого белка), который сам не является аминокислотой.Например, многие ферменты требуют связывания натрия для обеспечения правильного функционирования. Однако натрий не обязательно действует как регуляторная субъединица; натрий присутствует всегда, и нет известных биологических способов добавления / удаления натрия для регулирования активности ферментов. Нерегулирующая аллостерия может содержать любые другие ионы, помимо натрия (кальция, магния, цинка), а также другие химические вещества и, возможно, витамины.
Фармакология
Аллостерическая модуляция рецептора возникает в результате связывания аллостерических модуляторов в другом сайте («регуляторный сайт»), отличном от сайта эндогенного лиганда («активный сайт»), и усиливает или ингибирует эффекты эндогенного лиганда.В нормальных условиях он действует, вызывая конформационные изменения в молекуле рецептора, что приводит к изменению аффинности связывания лиганда. Таким образом, аллостерический лиганд модулирует активацию рецептора своим первичным (ортостерическим) лигандом, и можно подумать, что он действует как диммер в электрической цепи, регулируя интенсивность ответа.
Например, рецептор GABA A имеет два активных сайта, которые связывает нейромедиатор гамма-аминомасляная кислота (ГАМК), но также имеет сайты связывания, регулирующие бензодиазепин и анестетик общего назначения.Каждый из этих регуляторных сайтов может производить положительную аллостерическую модуляцию, усиливая активность ГАМК. Диазепам является агонистом регуляторного участка бензодиазепина, а его антидот флумазенил является антагонистом.
Более свежие примеры лекарств, которые аллостерически изменяют свои мишени, включают цинакальцет, имитирующий кальций, и маравирок для лечения ВИЧ.
Аллостерические участки как мишени для лекарств
Аллостерические сайты могут представлять собой новую мишень для лекарств. Существует ряд преимуществ использования аллостерических модуляторов в качестве предпочтительных терапевтических агентов по сравнению с классическими ортостерическими лигандами.Например, сайты аллостерического связывания рецептора, связанного с G-белком (GPCR), не столкнулись с таким же эволюционным давлением, как ортостерические сайты, чтобы приспособиться к эндогенному лиганду, поэтому они более разнообразны. [8] Следовательно, большая селективность GPCR может быть получена путем нацеливания на аллостерические сайты. [8] Это особенно полезно для GPCR, где селективная ортостерическая терапия была затруднена из-за сохранения последовательности ортостерического сайта у подтипов рецепторов. [9] Кроме того, эти модуляторы обладают пониженным потенциалом токсических эффектов, поскольку модуляторы с ограниченным взаимодействием будут иметь максимальный уровень своего действия, независимо от введенной дозы. [8] Другой тип фармакологической селективности, который является уникальным для аллостерических модуляторов, основан на кооперативности. Аллостерический модулятор может проявлять нейтральную кооперативность с ортостерическим лигандом на всех подтипах данного рецептора, кроме интересующего подтипа, который называется «абсолютной селективностью подтипа». [9] Если аллостерический модулятор не обладает заметной эффективностью, он может обеспечить другое мощное терапевтическое преимущество перед ортостерическими лигандами, а именно способность избирательно настраивать повышающие или понижающие тканевые ответы только при наличии эндогенного агониста. [9] Олигомер-специфические сайты связывания малых молекул являются лекарственными мишенями для имеющих медицинское значение морфеинов. [10]
См. Также
Список литературы
- ↑ Bu Z, Callaway DJ (2011). «Белки ДВИГАЮТСЯ! Динамика белков и дальняя аллостерия в передаче сигналов в клетке». Адвокат в области химии белков и структурной биологии . Достижения в химии белков и структурной биологии. 83 : 163–221. DOI: 10.1016 / B978-0-12-381262-9.00005-7.ISBN 9780123812629 . PMID 21570668.
- Перейти ↑ J. Monod, J. Wyman, J.P. Changeux. (1965). О природе аллостерических переходов: правдоподобная модель. J. Mol. Биол. , май; 12: 88-118.
- ↑ D.E. Jr Koshland, G. Némethy, D. Filmer (1966) Сравнение экспериментальных данных связывания и теоретических моделей в белках, содержащих субъединицы. Биохимия . Янв; 5 (1): 365-8
- ↑ E.К. Яффе (2005). «Морфеины — новая структурная парадигма аллостерической регуляции». Trends Biochem. Sci . 30 (9): 490–497. DOI: 10.1016 / j.tibs.2005.07.003. PMID 16023348.
- ↑ Z. Huang, L. Zhu, Y. Cao, G. Wu, X. Liu, et al (2011) ASD: обширная база данных аллостерических белков и модуляторов. Nucleic Acids Res Volume 39, D663-669
- ↑ Шринивасан Б; и другие.(2014). «Аллостерическая регуляция и активация субстрата в цитозольной нуклеотидазе II из Legionella pneumophila». FEBS J . 281 (6): 1613–1628. DOI: 10.1111 / febs.12727. PMID 24456211.
- ↑ Эдельштейн, SJ (1975). «Кооперативные взаимодействия гемоглобина». Анну Рев Биохим . 44 : 209–232. DOI: 10.1146 / annurev.bi.44.070175.001233. PMID 237460.
- ↑ 8,0 8,1 8,2 A. Christopoulos, L.T. Мая, В.А. Авлани и П. Sexton (2004) Аллостеризм рецепторов, связанных с G-белками: перспективы и проблемы. Транзакции Биохимического Общества Том 32, часть 5
- ↑ 9,0 9,1 9,2 May, L.T .; Leach, K .; Секстон, П.М.; Кристопулос, А. (2007). «Аллостерическая модуляция рецепторов, связанных с G-белком». Annu.Rev. Pharmacol. Токсикол . 47 : 1–51. DOI: 10.1146 / annurev.pharmtox.47.120505.105159. PMID 17009927.
- ↑ Э. К. Яффе (2010). «Морфеины — новый путь к открытию аллостерических лекарств». Открытая конф. Proc. J . 1 : 1–6. DOI: 10.2174 / 2210289201001010001. PMC 3107518. PMID 21643557.
Внешние ссылки
Frontiers | Понимание роли комплексов гетерорецепторов GPCR в модуляции мозговых сетей при здоровье и болезнях
Основы гетерорецепторных комплексов GPCR, их аллостерической связи и их функции
Концепция взаимодействий аллостерический рецептор-рецептор в гомо- и гетерорецепторных комплексах G-белковых рецепторов (GPCR) центральной нервной системы (ЦНС) дала новое измерение интеграции мозга и нейропсихофармакологии (Fuxe et al., 1983, 1998, 2008c, 2010b, 2014d; Золи и др., 1993; Лю и др., 2000; Джордж и др., 2002; Guo et al., 2008; Хан и др., 2009; Borroto-Escuela et al., 2012a). Взаимодействия аллостерический рецептор-рецептор, ставшие возможными благодаря олигомеризации рецептора, приводят к новой динамике рецептора, в ходе которой протомеры рецептора изменяют свое распознавание, фармакологию, передачу сигналов и передачу, и могут развиваться новые сайты аллостерического связывания (Borroto-Escuela et al., 2011, 2012a; Fuxe et al. al., 2012a, 2014d; Fuxe, Borroto-Escuela, 2016).Гетерорецепторные комплексы GPCR могут также включать рецепторы ионных каналов, рецепторные тирозинкиназы (RTK), наборы белков, взаимодействующих с G-белком, ионные каналы и / или переносчики трансмиттеров (Fuxe et al., 2007; Flajolet et al., 2008; Guo et al. , 2008; Borroto-Escuela et al., 2012b, 2015c, d, 2016c; Fuxe and Borroto-Escuela, 2016; Di Liberto et al., 2017). Аллостерические взаимодействия в таких динамических рецепторных комплексах более высокого порядка имеют место организованным пространственно-временным образом и участвуют в обучении и формировании молекулярных энграмм для краткосрочной и долгосрочной памяти (Fuxe et al., 2014b; Borroto-Escuela et al., 2015a). Кроме того, комплекс рецепторов NMDA теперь также рассматривается как многофункциональный механизм в глутаматергическом синапсе, включающий внесинаптические и синаптические комплексы гетерорецепторов D1R-NMDAR и D2R-NMDAR (Liu et al., 2000, 2006; Zhang et al., 2016). Необходимо улучшить наше понимание молекулярной организации олигомеров рецепторов, их аллостерической связи и особенностей рецепторного интерфейса (Borroto-Escuela et al., 2010c, 2014a; Tarakanov and Fuxe, 2010).
Недавно молекулярная основа обучения и памяти была предложена на реорганизации гомо- и гетерорецепторных комплексов в постсоединительной мембране синапсов, приводящей к изменениям в предсоединительных рецепторных комплексах, чтобы облегчить изучение паттерна высвобождения медиатора (Fuxe et al. , 2014b; Borroto-Escuela et al., 2015a). Долговременная память может быть создана путем преобразования частей гетерорецепторных комплексов в уникальные факторы транскрипции, что может привести к образованию специфических адаптерных белков, которые могут объединять гетерорецепторные комплексы в долгоживущие комплексы с консервативными аллостерическими взаимодействиями рецептор-рецептор (Fuxe и другие., 2014а, б; Borroto-Escuela et al., 2015a). Таким образом, гомогетерорецепторные комплексы рассматриваются как высокодинамичные сборки, образованные или разрушенные интегрированными синаптическими и объемными сигналами передачи. Эти события необходимы для обучения и могут трансформироваться в консолидированное жесткое состояние с сохраненной аллостерической связью, представляющей молекулярные энграммы, что приводит к значительной долгосрочной модуляции нейронных сетей. Это изменение молекулярной пластичности, временное или долгосрочное, может затем изменить паттерны оттока в мозговых цепях и вызвать временные и долгосрочные изменения в поведении и когнитивных функциях.В соответствии с этой гипотезой блокирование синаптического удаления GluA2-содержащих рецепторов AMPA предотвращает естественное забывание долговременных воспоминаний (Migues et al., 2016).
Особое значение для структурной пластичности, например, в дендритном дереве и его шипах, может иметь рекрутирование RTK в образованные комплексы гетерорецепторов, что может приводить, например, к синергетическому увеличению плотности нейритов и их выступов в первичных культурах нейронов ( Flajolet et al., 2008; Borroto-Escuela et al., 2012b; Либманн и др., 2016).
Концепция предвзятого агонизма GPCR, означающего функциональную избирательность, была разработана Кенакином (2007, 2008, 2011). Было высказано предположение, что агонистическая стабилизация отдельных активных состояний в конформации рецептора является механизмом, участвующим в активации дискретных сигнальных путей GPCR. В 1983/1985 годах взаимодействия рецептор-рецептор и их значение для разнообразия рецепторов были представлены на основе исследований взаимодействий нейропептидов / дофамина (DA) (Agnati et al., 1983а, б; Fuxe et al., 1983; Fuxe и Agnati, 1985). Благодаря демонстрации сети гетеродимеров GPCR (GPCR-HetNet; Borroto-Escuela et al., 2014a) стало ясно, что взаимодействия аллостерический рецептор-рецептор резко увеличивают разнообразие GPCR и смещают распознавание и передачу сигналов, что приводит к повышенной специфичности передачи сигналов (Fuxe et al., 2014f; Borroto-Escuela et al., 2015b; Fuxe and Borroto-Escuela, 2016). Множественные источники разнообразия и специфичности GPCR были элегантно выяснены в 2005 г. (Maudsley et al., 2005).
Дисфункция комплексов гетерорецепторов GPCR может привести к заболеванию головного мозга
Дисфункция или нарушение гетерорецепторных комплексов D2R может быть молекулярной основой патологического изменения в мозговых цепях. Например, усиление функции D2R приводит к изменению активности префронтальных афферентов глутамата (Fuxe et al., 2008c), за которым следует развитие шизофренических симптомов. Понимание этих комплексов гетерорецепторов D2R и их дисфункции при шизофрении может привести к новым стратегиям их лечения и для предотвращения побочных эффектов нейролептиков, которые, как известно, в основном действуют как антагонисты D2R (Seeman, 2010), включая способ оптимизации комбинированного лечения или однократного применения гетеробивалентные препараты, нацеленные на комплексы гетерорецепторов D2R при шизофрении.Это вдохновлено текущими открытиями различных типов комплексов гетерорецепторов D2R (Borroto-Escuela et al., 2014c; Ferraro et al., 2014; Fuxe et al., 2014e, 2015; Pinton et al., 2015b; de la Mora и др., 2016).
С открытием многих изо и гетерорецепторных комплексов 5-HT1A, таких как 5-HT1A-5-HT7 (Renner et al., 2012), FGFR1-5-HT1A (Borroto-Escuela et al., 2012b, 2013a , 2015c, d, 2016c) и предполагаемого тримерного комплекса GalR1-GalR2-5-HT1A (Millón et al., 2014, 2016), на основе гипотезы 5-HT о молекулярной основе большой депрессии было получено более глубокое понимание молекулярной основы большой депрессии. депрессия (Carlsson et al., 1968; Рисунки 1, 2). Постсоединительные рецепторы 5-HT1A могут в значительной степени способствовать опосредованию антидепрессивных эффектов 5-HT (Artigas, 2015). Весьма интересно, что в этих гетерорецепторных комплексах белки, взаимодействующие с рецепторами, такие как каркасный белок p11 (Svenningsson, 2014; Milosevic et al., 2017; Schintu et al., 2016), по-видимому, играют существенную роль (Fuxe and Borroto -Эскуэла, 2016). У жертв самоубийств среди людей было установлено снижение мРНК p11 в гиппокампе и миндалине, а антидепрессанты усиливают экспрессию p11 в лимбических областях грызунов (Svenningsson et al., 2013).
Рис. 1. Восходящие и нисходящие пути 5-HT от рострального и каудального ядер шва показаны на левой панели. На правой панели показаны тельца и дендриты 5-HT иммунореактивных нервных клеток в дорсальном шве. Панорама комплексов гетерорецепторов 5-HT1A, включая изорецепторные комплексы 5-HT1A, продемонстрированные в дорсальном шве и в дорсальном гиппокампе, представлена в нижней правой части, показанной как гетеродимеры и, возможно, сбалансированные друг с другом.Они были описаны на постсинаптическом уровне, вероятно, в синаптических и внесинаптических местах. Существование этих 5-HT1A гетерокомплексов на нервном окончании 5-HT еще предстоит изучить. Они играют главную роль в модуляции 5-HT нейронных сетей в системе шов-гиппокамп вместе с соответствующими гоморецепторными комплексами и мономерами. Аббревиатуры см. В дополнительных материалах.
Рис. 2. Показан баланс различных гомо- и гетерорецепторных комплексов серотонина 5-HT1A и галанина (GalR1 и GalR2). Также представлены комплексы гетерорецепторов 5-HT1A-FGFR1, которые играют роль в депрессии. Взаимодействия аллостерический рецептор-рецептор, индуцируемые конститутивно или посредством активации одного рецепторного протомера, могут влиять на распознавание, связывание G-белка, передачу сигналов и транспорт других протомеров в гомо- и гетерорецепторном комплексе, как указано. Природа взаимодействий аллостерический рецептор-рецептор, обнаруженных в отдельных гетеромерах, указана в верхней части рецепторных комплексов ([-] антагонистическая аллостерическая модуляция, [+] способствующая аллостерическая модуляция).Передача сигналов протомеров рецептора отражает передачу сигналов базальных состояний. Аббревиатуры см. В дополнительных материалах.
Также была продемонстрирована значительная роль адаптерного белка, нарушенного при шизофрении в D2R гетерорецепторных комплексах (Su et al., 2014). Этот гетерокомплекс усиливает опосредованную D2R передачу сигналов киназы-3 гликогенсинтазы и снижает интернализацию D2R агонистами.
Гетерорецепторные комплексы GPCR как мишени для медикаментозного лечения заболеваний головного мозга
Гетерорецепторные комплексы GPCR в ЦНС стали захватывающими новыми мишенями для нейротерапевтов при болезни Паркинсона, шизофрении, расстройстве, вызванном употреблением психоактивных веществ, тревоге и депрессии, открывая новую область в нейропсихофармакологии (Portoghese, 2001; Soriano et al., 2009; Ле Наур и др.
практических тестов вопросы 5-7: вред Карточки
Нам не удалось определить язык звукового сопровождения на ваших карточках. Пожалуйста, выберите правильный язык ниже.
Фронт
Китайский, PinyinChinese, SimplifiedChinese, TraditionalEnglishFrenchGermanItalianJapaneseJapanese, RomajiKoreanMath / SymbolsRussianSpanishAfrikaansAkanAkkadianAlbanianAmharicArabicArmenianAzerbaijaniBasqueBelarusianBengaliBihariBretonBulgarianBurmeseCatalanCebuanoChamorroChemistryCherokeeChinese, PinyinChinese, SimplifiedChinese, TraditionalChoctawCopticCorsicanCroatianCzechDanishDeneDhivehiDutchEnglishEsperantoEstonianFaroeseFilipinoFinnishFrenchFulaGaelicGalicianGeorgianGermanGreekGuaraniGujaratiHaidaHaitianHausaHawaiianHebrewHindiHungarianIcelandicIgboIndonesianInuktitutIrishItalianJapaneseJapanese, RomajiJavaneseKannadaKazakhKhmerKoreanKurdishKyrgyzLakotaLaoLatinLatvianLingalaLithuanianLuba-KasaiLuxembourgishMacedonianMalayMalayalamMalteseMaoriMarathiMarshalleseMath / SymbolsMongolianNepaliNorwegianOccitanOjibweOriyaOromoOther / UnknownPashtoPersianPolishPortuguesePunjabiPāliQuechuaRomanianRomanshRussianSanskritSerbianSindhiSinhaleseSlovakSlovenianSpanishSundaneseSwahiliSwedishTaga logТаджикскийТамильскийТатарскийТелугуТайскийТибетскийТигриньяTohono O’odhamТонгаТурецкийУйгурскийУкраинскийУрдуУзбекскийВьетнамский Валлийский Западно-фризскийИдишЙоруба
аудио еще не доступно для этого языка
Назад
Китайский, PinyinChinese, SimplifiedChinese, TraditionalEnglishFrenchGermanItalianJapaneseJapanese, RomajiKoreanMath / SymbolsRussianSpanishAfrikaansAkanAkkadianAlbanianAmharicArabicArmenianAzerbaijaniBasqueBelarusianBengaliBihariBretonBulgarianBurmeseCatalanCebuanoChamorroChemistryCherokeeChinese, PinyinChinese, SimplifiedChinese, TraditionalChoctawCopticCorsicanCroatianCzechDanishDeneDhivehiDutchEnglishEsperantoEstonianFaroeseFilipinoFinnishFrenchFulaGaelicGalicianGeorgianGermanGreekGuaraniGujaratiHaidaHaitianHausaHawaiianHebrewHindiHungarianIcelandicIgboIndonesianInuktitutIrishItalianJapaneseJapanese, RomajiJavaneseKannadaKazakhKhmerKoreanKurdishKyrgyzLakotaLaoLatinLatvianLingalaLithuanianLuba-KasaiLuxembourgishMacedonianMalayMalayalamMalteseMaoriMarathiMarshalleseMath / SymbolsMongolianNepaliNorwegianOccitanOjibweOriyaOromoOther / UnknownPashtoPersianPolishPortuguesePunjabiPāliQuechuaRomanianRomanshRussianSanskritSerbianSindhiSinhaleseSlovakSlovenianSpanishSundaneseSwahiliSwedishTaga logТаджикскийТамильскийТатарскийТелугуТайскийТибетскийТигриньяTohono O’odhamТонгаТурецкийУйгурскийУкраинскийУрдуУзбекскийВьетнамский Валлийский Западно-фризскийИдишЙоруба
аудио еще не доступно для этого языка
Как понять категориальное при непрерывном взаимодействии? (Stata 12)
Во-первых, давайте начнем с того, что означает значимая категоризация под непрерывным взаимодействием.Это означает, что наклон непрерывной переменной различен для одного или нескольких уровней категориальной переменной.
Мы будем использовать пример из набора данных hsbdemo , который имеет статистически значимую категоризацию путем непрерывного взаимодействия, чтобы проиллюстрировать один из возможных объяснительных подходов.
Категориальная переменная — женщина , переменная ноль / единица, где женщины закодированы как единица (следовательно, мужчины являются контрольной группой). Переменная непрерывного предиктора, socst , представляет собой стандартизированный тестовый результат для социальных исследований.Мы начнем с запуска регрессионной модели и построения графика взаимодействия. Обратите внимание, что мы используем c.socst , чтобы указать, что socst является непрерывной переменной.
используйте https://stats.idre.ucla.edu/stat/data/hsbdemo, очистите
регресс написать женский ## c.socst
Источник | SS df MS Количество набл. = 200
------------- + ------------------------------ F (3, 196) = 49,26
Модель | 7685.43528 3 2561,81176 Вероятность> F = 0,0000
Остаточный | 10193,4397 196 52,0073455 R-квадрат = 0,4299
------------- + ------------------------------ Корректировка R-квадрат = 0,4211
Итого | 17878,875 199 89,843593 Корневой MSE = 7,2116
-------------------------------------------------- ----------------------------
написать | Коэф. Стд. Err. t P> | t | [95% конф. Интервал]
------------- + ------------------------------------ ----------------------------
1.женский | 15,00001 5,09795 2,94 0,004 4,946132 25,05389
socst | .6247968 .0670709 9,32 0,000 .4925236 .7570701
|
женский # |
c.socst |
1 | -.2047288 .0953726 -2,15 0,033 -,3928171 -,0166405
|
_cons | 17,7619 3,554993 5,00 0,000 10,75095 24,77284
-------------------------------------------------- ----------------------------
twoway (scatter write socst, msym (oh) jitter (3)) ///
(lfit написать socst, если ~ женский) (lfit написать socst, если женский), ///
легенда (заказ (2 «самец» 3 «самка»))
Глядя на график, мы видим, что две линии регрессии не параллельны, и что линия для женщин проходит выше линии для мужчин.Как мы могли сказать, что женщины выше мужчин? Коэффициент для женщин положительный (15,00)
что говорит нам, что уровень для женщин выше, чем для мужчин.
Давайте интерпретируем коэффициенты для этой модели, начиная с константы (17,76). Это значение точки пересечения для socst , регрессированное на , запись для мужчин. т.е. ожидаемое значение для запишите , когда и socst , и female равны нулю.
Коэффициент для socst равен 0,6247, который представляет собой наклон линии регрессии для мужской группы (т.е. женщина = 0). Значение для женщин по взаимодействию socst составляет -2047, что представляет собой разницу в наклоне между мужской и женской группой, то есть наклон для женской группы будет примерно 0,6248 — 0,2047 = 0,4201.
Мы также можем получить наклоны для двух групп с помощью команды margins .
поля женские, dydx (socst)
Средние предельные эффекты Количество наблюдений = 200
Модель VCE: OLS
Выражение: линейное предсказание, предсказать ()
dy / dx w.r.t. : socst
-------------------------------------------------- ----------------------------
| Дельта-метод
| dy / dx стандарт. Err. z P> | z | [95% конф. Интервал]
------------- + ------------------------------------ ----------------------------
socst |
женский |
0 | .6247968 .0670709 9,32 0,000 .4933403 .7562533
1 | .420068 .0678044 6,20 0,000.2871739 .5529622
-------------------------------------------------- ---------------------------- Разница между мужчинами и женщинами может существенно отличаться или не отличаться для разных значений socst . Что мы
Достаточно посмотреть на разницу между мужчинами и женщинами для различных значений socst , используя команду margins . Мы позволим socst изменяться от 25 до 70 с шагом 5.
поля, при (женщина = (0 1) socst = (25 (5) 70)) vsquish
Скорректированные прогнозы Количество наблюдений = 200
Модель VCE: OLS
Выражение: линейное предсказание, предсказать ()
1._at: female = 0
socst = 25
2._at: female = 0
socst = 30
3._at: female = 0
socst = 35
4._at: женский = 0
socst = 40
5._at: женский = 0
socst = 45
6._at: female = 0
socst = 50
7._at: female = 0
socst = 55
8._at: female = 0
socst = 60
9._at: female = 0
socst = 65
10._at: female = 0
socst = 70
11._at: female = 1
socst = 25
12._at: female = 1
socst = 30
13._at: female = 1
socst = 35
14._at: female = 1
socst = 40
15._at: female = 1
socst = 45
16._at: женский = 1
socst = 50
17._at: female = 1
socst = 55
18._at: female = 1
socst = 60
19._at: female = 1
socst = 65
20._at: female = 1
socst = 70
-------------------------------------------------- ----------------------------
| Дельта-метод
| Маржа Станд. Err. z P> | z | [95% конф.Интервал]
------------- + ------------------------------------ ----------------------------
_at |
1 | 33,38182 1,94946 17,12 0,000 29,56095 37,20269
2 | 36,5058 1,645495 22,19 0,000 33,28069 39,73091
3 | 39,62979 1,356406 29,22 0,000 36,97128 42,28829
4 | 42.75377 1.094051 39.08 0.000 40.60947 44.89807
5 | 45,87775 .8825999 51,98 0.000 44.14789 47.60762
6 | 49.00174 .7654688 64.02 0.000 47.50145 50.50203
7 | 52.12572 .78602 66.32 0.000 50.58515 53.66629
8 | 55.24971 .9352206 59.08 0.000 53.41671 57.0827
9 | 58.37369 1.164634 50.12 0.000 56.09105 60.65633
10 | 61,49767 1,436326 42,82 0,000 58,68253 64,31282
11 | 43,26361 2,015017 21,47 0.000 39.31425 47.21297
12 | 45,36395 1,700513 26,68 0,000 42,031 48,69689
13 | 47.46429 1.397521 33.96 0.000 44.72519 50.20338
14 | 49,56463 1,115464 44,43 0,000 47,37836 51,75089
15 | 51.66497 .8748286 59.06 0.000 49.95033 53.3796
16 | 53.76531 .7185139 74.83 0.000 52.35704 55.17357
17 | 55,86565 .7050327 79,24 0.000 54,48381 57,24748
18 | 57.96599 .8412798 68.90 0.000 56.31711 59.61486
19 | 60.06633 1.071589 56.05 0.000 57.96605 62.1666
20 | 62.16667 1.348602 46.10 0.000 59.52346 64.80988
-------------------------------------------------- ---------------------------- Итак, записывает значение для мужчин в socst = 25 равно 33,38182, как показано в строке 1. То же значение для женщин — 43.26361, как показано в строке 11. Вот различия в двух значениях: 43,26361 — 33,38182 = 9,88179. Мы можем получить эту разницу для всех значений socst , используя команду margins с опцией dydx .
поля, dydx (женский) at (socst = (25 (5) 70)) vsquish
Условные маргинальные эффекты Количество наблюдений = 200
Модель VCE: OLS
Выражение: линейное предсказание, предсказать ()
dy / dx w.r.t. : 1. женский
1._at: socst = 25
2._at: socst = 30
3._at: socst = 35
4._at: socst = 40
5._at: socst = 45
6._at: socst = 50
7._at: socst = 55
8._at: socst = 60
9._at: socst = 65
10._at: socst = 70
-------------------------------------------------- ----------------------------
| Дельта-метод
| dy / dx стандарт.Err. z P> | z | [95% конф. Интервал]
------------- + ------------------------------------ ----------------------------
1. женский |
_at |
1 | 9,881789 2,803692 3,52 0,000 4,386654 15,37692
2 | 8,858145 2,366305 3,74 0,000 4,220273 13,49602
3 | 7,834501 1,947538 4,02 0,000 4,017396 11,6516
4 | 6,810857 1,562436 4,36 0,000 3,748538 9.873176
5 | 5,787213 1,242702 4,66 0,000 3,351562 8,222863
6 | 4,763569 1,049859 4,54 0,000 2,705882 6,821255
7 | 3,739925 1,055888 3,54 0,000 1,670423 5,809426
8 | 2,716281 1,257931 2,16 0,031 0,250782 5,181779
9 | 1,692637 1,582617 1,07 0,285 -1,409236 4,794509
10 | .6689926 1,970219 0,34 0,734 -3,192565 4.53055
-------------------------------------------------- ----------------------------
Примечание: dy / dx для уровней факторов - это дискретное изменение от базового уровня. Теперь мы можем изобразить эти различия, используя команду marginsplot .
marginsplot, yline (0)
Мы видим, что различия между мужчинами и женщинами значительны для значений socst ниже примерно 60. Этот график хорош и рассказывает историю, которую мы хотим знать, но это не самый красивый график, который мы можем нарисовать.Изменяя линии и доверительные интервалы, мы получаем гораздо более четкий график.
marginsplot, recast (line) recastci (раритет) yline (0)
А, это намного лучше.
График показывает, что разница между мужчинами и женщинами уменьшается с увеличением значения socst . Если 95% доверительный интервал для разницы не включает ноль, это различие можно считать статистически значимым. Похоже, это справедливо для всех значений от socst до примерно 60.Для значений socst больше 60 разница между мужчинами и женщинами несущественна.
Трехуровневая категориальная переменная
Что делать, если у вашей категориальной переменной более двух уровней? Набор данных catcon3l имеет категориальный предиктор, b , с тремя уровнями. Переменная ответа — y, категориальный предиктор — b , и она взаимодействует с непрерывным предиктором x , указанным в Stata как c.х .
используйте https://stats.idre.ucla.edu/stat/data/catcon3l, очистите
регресс y b ## c.x
Источник | SS df MS Количество набл. = 200
------------- + ------------------------------ F (5, 194) = 32,02
Модель | 8083.95798 5 1616.7916 Вероятность> F = 0,0000
Остаточный | 9794,91702 194 50,489263 R-квадрат = 0,4522
------------- + ------------------------------ Корректировка R-квадрат = 0 .4380
Итого | 17878,875 199 89,843593 Корневой MSE = 7,1056
-------------------------------------------------- ----------------------------
y | Коэф. Стд. Err. t P> | t | [95% конф. Интервал]
------------- + ------------------------------------ ----------------------------
б |
2 | -14,56722 7,178201 -2,03 0,044 -28,72455 -4098837
3 | -20,16644 6,779483 -2.97 0,003 -33,53739 -6,795485
|
х | .2728512 .086295 3,16 0,002 .1026544 .4430479
|
b # c.x |
2 | .1349579 .1453856 0,93 0,354 -1517813 .4216972
3 | .3012428 .1203257 2,50 0,013 .0639282 .5385573
|
_cons | 42.03102 4.907897 8.56 0.000 32.35134 51.71071
-------------------------------------------------- ----------------------------
/ * тест общей значимости взаимодействия * /
testparm b # c.х
(1) 2.b # c.x = 0
(2) 3.b # c.x = 0
F (2, 194) = 3,15
Вероятность> F = 0,0453
/* или */
контраст b # c.x
Контрасты предельных линейных предсказаний
Маржа: асбалансированная
------------------------------------------------
| df F P> F
------------- + ----------------------------------
b # c.x | 2 3.15 0,0453
|
Остаточный | 194
------------------------------------------------ Команды testparm и / или Contrast показывают, что общее взаимодействие является статистически значимым.
Затем мы вычислим простые уклоны с помощью команды margins .
поля b, dydx (x)
Средние предельные эффекты Количество наблюдений = 200
Модель VCE: OLS
Выражение: линейное предсказание, предсказать ()
dy / dx w.r.t. : Икс
-------------------------------------------------- ----------------------------
| Дельта-метод
| dy / dx стандарт. Err. z P> | z | [95% конф. Интервал]
------------- + ------------------------------------ ----------------------------
х |
б |
1 | .2728512 .086295 3,16 0,002 .1037161 .4419862
2 | .4078091 .1170049 3,49 0,000.1784837 .6371345
3 | .5740939 .0838538 6,85 0,000 .4097435 .7384444
-------------------------------------------------- ---------------------------- Наклон для b = 1 кажется отличным от наклона b , равного 2 или 3.
Теперь давайте изобразим наклоны вместе с диаграммой рассеяния данных. Мы сделаем это, незаметно запустив margins (чтобы подавить большой вывод), а затем команду marginsplot с добавленной диаграммой рассеяния.
спокойно поля b, при (x = (25 (5) 70)) marginsplot, recast (line) noci addplot (scatter y x, jitter (3) msym (oh))
Давайте посмотрим, существенно ли отличается наклон для b = 3 от каждого из двух других наклонов. Мы проверим это, используя эталонные контрасты с помощью команды margins . Мы укажем, что наклон 3 является эталоном, используя b3 и контрольную группу
кодирование с r , которые объединяются с rb3 .
поля rb3.b, dydx (x)
поля rb3.b, dydx (x)
Контрасты средних предельных эффектов
Модель VCE: OLS
Выражение: линейное предсказание, предсказать ()
dy / dx w.r.t. : Икс
------------------------------------------------
| df chi2 P> chi2
------------- + ----------------------------------
х |
б |
(1 против 3) | 1 6,27 0.0123
(2 против 3) | 1 1,33 0,2480
Совместное | 2 6,29 0,0430
------------------------------------------------
-------------------------------------------------- ------------
| Контраст Дельта-метод
| dy / dx стандарт. Err. [95% конф. Интервал]
------------- + ------------------------------------ ------------
х |
б |
(1 против 3) | -.3012428 .1203257 -.5370769 -.0654086
(2 против 3) | -1662848 .14395 -.4484217 .115852
-------------------------------------------------- ------------ Мы видим, что склон 3 существенно отличается от уклона 1, но не отличается от уклона 2.
Глядя на три наклона, можно задаться вопросом, где различия между группами статистически значимы. Самый естественный способ сделать это — выбрать контрольную группу, на этот раз b = 1, и посмотреть, где значения для b = 2 отличаются, а затем такие же для b1 по сравнению с b3 .Опять же, на ум приходит команда margins с опцией dydx .
поля, dydx (b) at (x = (25 (5) 70)) vsquish Условные маргинальные эффекты Количество наблюдений = 200 Модель VCE: OLS Выражение: линейное предсказание, предсказать () dy / dx w.r.t. : 2.b 3.b 1._at: x = 25 2._at: x = 30 3._at: x = 35 4._at: x = 40 5._at: x = 45 6._at: x = 50 7._at: x = 55 8._at: x = 60 9._at: x = 65 10._at: x = 70 -------------------------------------------------- ---------------------------- | Дельта-метод | dy / dx стандарт.Err. z P> | z | [95% конф. Интервал] ------------- + ------------------------------------ ---------------------------- 2.b | _at | 1 | -11.19327 3.704771 -3.02 0.003 -18.45448 -3.93205 2 | -10,51848 3,055425 -3,44 0,001 -16,507 -4,529954 3 | -9,843688 2,450055 -4,02 0,000 -14,64571 -5,041668 4 | -9.168898 1.930483 -4.75 0.000 -12.95258 -5.385221 5 | -8,494108 1,583543 -5,36 0,000 -11,59779 -5,36 | -7,819319 1,531437 -5,11 0,000 -10,82088 -4,817758
7 | -7,144529 1,799955 -3,97 0,000 -10,67238 -3,616682
8 | -6,469739 2,278427 -2,84 0,005 -10,93537 -2,004105
9 | -5,79495 2,863471 -2,02 0,043 -11,40725 -,1826503
10 | -5,12016 3,502078 -1,46 0,144 -11,98411 1.743787
————- + ———————————— —————————-
3.b |
_at |
1 | -12,63537 3,853374 -3,28 0,001 -20,18784 -5,082896
2 | -11,12916 3,285978 -3,39 0,001 -17,56956 -4,688757
3 | -9,622942 2,733264 -3,52 0,000 -14,98004 -4,265844
4 | -8.116729 2.206291 -3.68 0.000 -12.44098 -3.792477
5 | -6,610515 1.728765 -3,82 0,000 -9,998831 -3,222199
6 | -5,104301 1,354048 -3,77 0,000 -7,758187 -2,450415
7 | -3,598087 1,184137 -3,04 0,002 -5,918953 -1,277221
8 | -2,091873 1,301856 -1,61 0,108 -4,643464,4597175
9 | -,5856595 1,64663 -0,36 0,722 -3,812996 2,641677
10 | .9205543 2,109945 0,44 0,663 -3,214862 5,055971
————————————————— —————————-
Примечание: dy / dx для уровней факторов — это дискретное изменение от базового уровня.
Первый блок результатов, 2.b сравнивает b1 с b2 и значим для значений x меньше 70. Второй блок, b3 сравнивает b1 с b3 и является значимым. для x значений меньше 60. Давайте изобразим эти результаты полей .
marginsplot, recast (line) recastci (раритет) yline (0)
Заштрихованная синим область — b1 против b2, а заштрихованная красным область — b1 против b3.График подтверждает нашу интерпретацию таблицы полей .
PPT — Интерпретация эффектов аллостерических агентов Презентация PowerPoint
100 Переход в состояние R Частичное насыщение Переход в состояние T 50 pO2 Интерпретация эффектов аллостерических агентов, связывающих O2 95 80 O2 OH- Высвобождение CO2 O2 -Hb BPG 10 H +
4 ПРАВИЛА, регулирующие аллостерические белки ПРАВИЛО: Аллостерия — это свойство белков, которые содержат несколько субъединиц или одну субъединицу с несколькими сайтами связывания.ПРАВИЛО: Аллостерические белки должны быть способны связывать более одного лиганда. ПРАВИЛО: лиганд обладает способностью вызывать изменение формы аллостерического белка. ПРАВИЛО: Изменяя форму белка, лиганд может способствовать (положительно) или сдерживать (отрицательно) связывание дополнительных лигандов с белком.
МОДЕЛИ АЛЛОСТЕРИИ См. Стратегии на стр.120 КОНЦЕРТНО: (ВСЕ ИЛИ НЕТ) Субъединицы существуют в двух конформационных состояниях Два состояния находятся в равновесии Все субъединицы имеют одинаковую конформацию; нет гибридов Лиганд связывается с любой субъединицей в любой конформации Сохранение симметрии является движущей силой
Проблемы с согласованной моделью • Симметрия не сохраняется в большинстве олигомерных белков • Некоторые аллостерические олигомерные белки не имеют идентичных субъединиц • Невозможно объяснить в модель с двумя состояниями, как отрицательная и положительная кооперативность возникают в одном и том же белке
Последовательная модель SSSSSSSSSSSSSS Связывание лиганда вызывает изменение субъединицы Изменение прогрессивное Симметрия не сохраняется Механическое сцепление между субъединицами может быть слабым
Серповидная клетка Анемия (генетическое заболевание) Одна аминокислота в белке может иметь значение между жизнью и смертью Искажение Это мембранный белок? Это цитозольный белок? Это что-то другое, кроме протеина?
Hb (альфа) N-Val-Leu-Ser-Pro-Ala-Asp-Lys-Thr …….. Glu Hb (бета) N-Val-His-Leu-Thr-Pro -Glu-Lys ……. Val Sickle Cell Glu6 Val6 (каждая бета-цепь) Glu GAA или GAG Val GUA или GUG Transposition AU заменяет A в кодоне (обмен пиримидин / пурин)
Val Полимеризованный дезокси-Hb Агрегаты Нерастворимые волокна искажают общая форма гемоглобина Искажает общую форму эритроцита. Искаженные эритроциты не могут проходить через капилляры
Антитела Древние греки: фукидиды (460-400 г.В.) Говоря о чуме в Афинах, тем не менее, именно в тех, кто выздоровел от болезни, больные и умирающие находили наибольшее сострадание. Они знали, что это было на собственном опыте, и теперь не боялись за себя; поскольку один и тот же человек никогда не подвергался нападению дважды — по крайней мере, со смертельным исходом.
Ранние теории Полинг 1940: «Инструктивная» теория Антигены действуют как шаблоны, которые направляют сворачивание зарождающейся цепи антитела Макфарлейн Бернет: Селективная или клональная теория Сайт связывания в молекуле антитела полностью определяется до того, как он встречается антиген
Костный мозг Лимфоциты-предшественники лейкоциты макрофаги B-лимфоциты T-лимфоциты (T-клетки) Антитела Цитотоксические T-клетки (TC-клетки) Гуморальные T-клетки-помощники (TH-клетки) Цитокины, опосредованные клетками
905 Основные характеристики • Каждая В-клетка, вырабатывающая антитела, вырабатывает один вид антител… антиген не требуется.• Специфичность определяется аминокислотной последовательностью… ДНК каждой клетки уникальна. • Незрелые клетки, которые вырабатывают антитела, разрушаются в раннем возрасте… самотолерантность • Зрелые клетки вырабатывают и отображают антитела на своей поверхности • Взаимодействие с антигеном запускает клетку деление, создавая большое количество определенного антитела, которое сохраняется до тех пор, пока антиген не исчезнет
Антитела представляют собой большие сложные молекулы Состав: антигенсвязывающий сайт Постоянная переменная Шарнирная область 2 тяжелые, 2 легкие цепи Дисульфидные связи
5 классов антител (иммуноглобулины, Ig) • IgA… наружные выделения, слезы, слюна, слизистая оболочка бронхов и кишечника • IgG… основное антитело в сыворотке, первоначально названное гамма-глобулином • IgD… наименее изучено • IgE… не имеет четкой функции, имеет рецептор на тучных клетках и стимулирует выработку гистамина и связано с аллергическими реакциями • IgM… первый класс появляются, самые высокие сайты комбинирования, эффективны против бактерий
Механизм • Синхронизация предсуществующих В-клеток размеры сначала IgM • IgG, IgA, IgD, IgE той же специфичности создаются позже • Легкая цепь не изменяется во время переключения • Вариабельная область тяжелой цепи не изменяется во время переключения • Только постоянная область тяжелой цепи изменяется… (переключение класса или переключение CH ) • У мышей при переключении используется соответствующий ген константной области, [C, C, C, C, C] для класса антител
Клеточно-опосредованный иммунитет Главный комплекс гистосовместимости (MHC) Функция: В сочетании с контролем Т-клеток, MHC представляет собой серию полиморфных белков, предназначенных для отображения фрагментов переваренных пептидов на поверхности клетки.Основа клеточного иммунитета MHC-1 На всех поверхностях клеток Отображает фрагменты из внутренних перевариваемых белков Взаимодействует с клетками TC, чтобы инициировать разрушение инфицированных клеток MHC-2 На поверхности фагоцитарных клеток (макрофагов и т. Д.) Отображает переваренные фрагменты из внешних источников белка Взаимодействует с клетками TH, чтобы предупредить систему об инфекции
Важно! Внутренние белки, переваренные протеазами, не могут связываться с MHC-2. Взаимодействие MHC-1 является сигналом для TC-клеток о разрушении инфицированной клетки. Внешние белки, которые проникают в клетку по эндосомному пути, не могут связываться с MHC-1.Взаимодействие MHC-2 является сигналом для TH-клеток о том, что система подвергается атаке вируса, бактерий и т. Д., И для отправки за помощью в виде продуцирующих антитела клеток
Модели взаимодействия | TeachingEnglish | Британский Совет
перейти к содержанию
- Справка
- Вход
- Зарегистрироваться
- Информационный бюллетень
- LearnEnglish Teens
- УзнатьАнглийский
- LearnEnglish Kids
- Домой
- Повышение квалификации
- Учителя
- Преподаватели-преподаватели
- Публикации
- Справочные книги
- Примеры из практики, идеи и исследования
- Основные этапы развития ELT
- База данных исследований ELT
- Награды за исследования ELT
- магистров ELT докторских диссертаций
- Учебные ресурсы
- Начальная школа
- Средняя школа
- Обучение взрослых
- Новости и события
- Главные новости
- IATEFL Online
- Вебинары
- Конференции
- Всемирный день учителя
- Семинары
- Награды за инновации ELTons
- Обучение
Форма поиска
Поиск
- Дом
- Повышение квалификации
- Публикации
- Учебные ресурсы
- Новости и события
- Обучение
- Справка
- Вход
- Зарегистрироваться
- Информационный бюллетень
- LearnEnglish Teens
- УзнатьАнглийский
- LearnEnglish Kids
Вы находитесь здесь
Главная »Знать предмет» База данных педагогических знаний
3.