Электрофорез для детей на воротниковой зоне, также проведения электрофорез для грудничков в Евромед
Проведение лечебных процедур электрофорезом в Омске. Электрофорез — метод введения лечебных препаратов через кожный покров, под действием электрического поля. При этом лекарство вводится в наиболее химически активной форме.
- Консультация
- Диагностика
- Лечение
Электрофорез — это метод введения лекарственного вещества под действием электрического поля в организм. Лечение электрофорезом проводится как через кожные покровы (в неврологии, травматологии и др.), так и через слизистые оболочки (в стоматологии, оториноларингологии, гинекологии и др.). Вещество вводится непосредственно в очаг воспаления в малых, но достаточно эффективных дозах, накапливаясь в тканях и действуя продолжительное время.
Поэтому особенно актуален электрофорез для детей, т.к. дает возможность местно использовать лекарства, которые в раннем детском возрасте нельзя применять другими способами.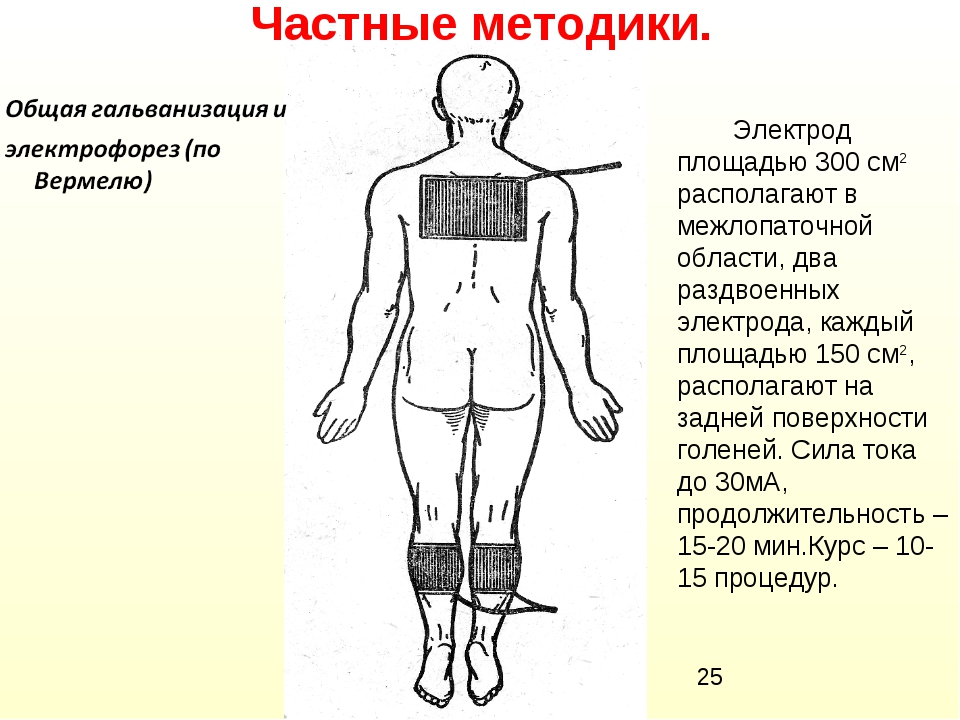 При этом лекарство вводится в наиболее химически активной форме — в виде ионов – и не разрушается, как например, при введении через рот. При проведении электрофореза электрический ток благоприятно влияет на реактивность и иммунобиологический статус тканей.
При этом лекарство вводится в наиболее химически активной форме — в виде ионов – и не разрушается, как например, при введении через рот. При проведении электрофореза электрический ток благоприятно влияет на реактивность и иммунобиологический статус тканей.
Этот метод применяется применяется с самого раннего возраста — например, электрофорез для грудничков используется уже спустя 3-6 недель после рождения. Для детей до года при этой процедуре в качестве лекарственного вещества часто используется эуфиллин как средство, снимающее тонус гладкой мускулатуры и улучшающее кровообращение (в основном, проводится электрофорез на воротниковую зону).
В целом выбор зоны проведения электрофорез зависит от заболевания: так, электрофорез воротниковой зоны проводится при заболеваниях нервной или сосудистой системы, электрофорез конечностей — при заболеваниях суставов, электрофорез поясничной зоны — при гинекологических заболеваниях и т.п.
Вернуться на страницу:
Остались вопросы?
Электрофорез Клиника Прогноз
Эффективное сочетание лечебного воздействия на организм постоянного тока и лекарственных препаратов, которые вводятся с его помощью.
С какого возраста можно проводить?
Процедуры можно проводить младенцам с 3-х месяцев (при условии, что вес ребенка не менее 3-х кг), детям раннего, дошкольного и школьного возраста.
ри каких нарушениях назначаются эти процедуры и как они проводятся?
Электрофорез по Ратнеру (с эуфиллином и папаверином):
- Нарушения кровообращения в шейном отделе позвоночника
- Травмы и гипоксия во время беременности и родов
- Кривошея
- ДЦП
- ЗПР
- Специфические расстройства речевого развития (особенно эффективен в сочетании с логопедическим массажем и логопедическими занятиями)
Электроды накладываются сзади в районе шейно-воротниковой зоны, спереди на правую часть грудной клетки. Время воздействия 15-20 мин. Количество процедур 10-14.
Время воздействия 15-20 мин. Количество процедур 10-14.
Электрофорез с магнием:
- Повышенная возбудимость нервной системы
- СДВГ
- Нарушение сна
- Астено-невротический синдром
- Тики
- Заикания
- Вегетативная дисфункция
- Головные боли
Электроды накладываются сзади в районе шейно-воротниковой зоны. Время воздействия 15-20 мин. Количество процедур 10-14.
Электрофорез со спазмолитиком на нижние конечности:
- ДЦП спастическая форма
- «Цыпочки» (ребенок часто ходит, опирая только на переднюю часть стопы)
- Плоско-вальгусная стопа
Электроды накладываются на икроножные мышцы. Время воздействия 15-20 мин. Количество процедур 10-12.
акие плюсы данной схемы введения лекарственных средств?
- Минимальное количество побочных эффектов, т.к. препарат попадает в организм, минуя пищеварительный тракт
- Местное воздействие с высокой концентрацией вещества помогает доставить лекарство непосредственно в зону патологии, не насыщая весь организм
- Безболезненность и неинвазивность процедуры, во время процедуры пациент не испытывает болезненных ощущений, нет повреждений кожи и слизистых
- Пролонгирующее действие лекарственного вещества, создание в коже своеобразного «депо» с лекарством, которое обеспечивает продолжительный терапевтический эффект и позволяет снизить дозировку препарата
- Действие лекарственного препарата усиливается за счет постоянного электрического тока
- Увеличивается эффективность других методов немедикаментозного лечения, которые используются в клинике «Прогноз».


акие противопоказания?
- Индивидуальная непереносимость постоянного тока или вводимого вещества
- Острые воспалительные и гнойные процессы
- Заболевания кожи(экзема, дерматиты)
- Новообразования
- Эпилепсия
- Системные заболевания внутренних органов
- Нарушение целостности кожного покрова в месте наложения электродов
- Лихорадка
- Не рекомендуется сочетать с другими методами физиотерапии с использованием электротока
Кто составляет протокол проведения процедур?
Врач-физиотерапевт после проведения неврологической функциональной диагностики
ФИЗИОТЕРАПЕВТИЧЕСКОЕ ОТДЕЛЕНИЕ №1
Фельдман Рита Михайловна
Заведущая физиотерапевтического отделения
Врач высшей квалификационной категории.
Физиокабинеты больницы открывались одновременно с открытием взрослой поликлиники и взрослого стационара, затем детского стационара и детской поликлиники. С первого дня функционирования лечебных корпусов физиолечение, было включено в лечение для взрослых больных и детей, оставаясь одним из эффективных методов лечения и до настоящего времени.
С первого дня функционирования лечебных корпусов физиолечение, было включено в лечение для взрослых больных и детей, оставаясь одним из эффективных методов лечения и до настоящего времени.
Физиотерапевтическое отделение обслуживает больных взрослого и детского стационаров, взрослой и детской поликлиники, обслуживающих население Привокзального и Советского районов.
В физиокабинете детского стационара лечатся больные из педиатрического отделения, психоневрологического отделения для детей, хирургического отделения для детей, отделения патологии новорожденных, отделения для выхаживания недоношенных детей, отоларингологического отделения.
В физиокабинете взрослого стационара лечатся больные из урологического отделения, кардиологического отделения, нейрососудистого отделения.
Население детской и взрослой поликлиники получает физиолечение по заболеваниям верхних дыхательных путей, заболеваниям бронхов и легких, заболеваниям нервной системы, заболеваниям органов пищеварения, мочевой системы, заболевания сердечно-сосудистой системы, заболевания костно-мышечной системы.
В состав физиотерапевтического отделения входит кабинет физиотерапии в детской поликлинике, взрослой поликлинике, кабинет электролечения, светолечения, кабинет ультравысокочастотной терапии, ингаляторий, кабинет парафинолечения.
Во всех физиокабинетах проводится электрофорез с эуфиллином на воротниковую зону, на грудную клетку; электрофорез с магнезией на воротниковую зону, на грудную клетку, на область правого подреберья; электрофорез хлористого кальция эндоназально и на область тазобедренных суставов, применяется электрофорез новокаина и анальгина на область позвоночника, суставов, а так же введение этих лекарств с помощью импульсивных токов. При экссудативных отитах, тугоухости используется электрофорез иодида калия эндоурально. Широко используется амплипульстерапия и диадинамотерапия при заболеваниях нервной системы и опорно — двигательного аппарата.
В отдельном кабинете отпускаются процедуры ультравысокочастотной терапии при острых воспалительных и гнойных заболеваниях различных органов и тканей. В отделении имеется достаточное количество аппаратов магнитотерапии «Алмаг 02», поэтому магнитотерапия широко используется при различных заболеваниях.
В отделении имеется достаточное количество аппаратов магнитотерапии «Алмаг 02», поэтому магнитотерапия широко используется при различных заболеваниях.
При многих заболеваниях слизистой оболочки, кожи и подкожной клетчатки используется общее и местное ультрафиолетовое облучение.
При заболеваниях верхних дыхательных путей, бронхитах, пневмониях применяются тепловлажные ультразвуковые ингаляции различных медикаментов. При аденоидитах, риносинуситах эффективно используется лазерная терапия эндоназально.
При различной патологии мочевого пузыря, мочевыводящих путей используется ультразвуковая терапия и введение лекарственных средств с помощью ультразвука. В кабинете парафинолечения с большой эффективностью используется парафино-озокеритовая апликации на различные области организма.
Все физиокабинеты оснащены современной аппаратурой: аппаратами магнитотерапии, ультравысокочастотной терапии, светотерапии, аппаратами лазерной терапии, ультразвуковой терапии, аппаратами электротерапии.
В физиотерапевтическом отделении №1 работают мед сестры с большим стажем работы, имеющие высшую и первую квалификационную категорию , повышающие свою квалификацию каждые 5 лет на базе колледжа последипломного образования средних мед работников.
Все мед сестры быстро осваивают работу на физиотерапевтическом оборудовании, поступающим в отделение. В отделении ежемесячно проводятся сестринские конференции по различной тематике, занятия по технике безопасности и противопожарной безопасности, занятия по гражданской обороне.
В 2014 году усилиями администрации больницы открыт кабинет лечебной физкультуры и кабинет массажа в детском стационаре. Кабинет лечебной физкультуры в течении года был полностью оснащен необходимым оборудованием: а именно, шведской стенкой из трех секций, гимнастической скамейкой, детскими велотренажерами, беговой дорожкой, манупедом и степпером, а также большим количеством различных мячей и гимнастическими палочками, что позволило более эффективно проводить реабилитацию детей при: ДЦП, остеопатиях, пневмониях, ортопедической патологии.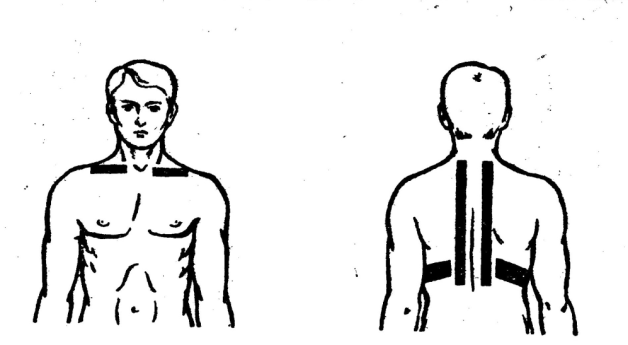 В кабинете массажа установлен современный массажный стол, что помогает массажистке успешно проводить курсы массажа у больных детей.
В кабинете массажа установлен современный массажный стол, что помогает массажистке успешно проводить курсы массажа у больных детей.
Физиотерапия
Из физических методов, воздействующих на ЦНС, в санаториях широко применяются электросон и лекарственный электрофорез. Для достижения успокаивающего эффекта больным с кардиалгическим, гипертензивным, аритмическим синдромами, при выраженных невротических проявлениях электросон в санатории Кисловодска применяют при частоте импульсного тока 10—15 Гц при силе тока — 7—8 мА. Продолжительность процедуры — 20—50 мин. Проводят их ежеднев но или 4—5 раз в неделю. На курс — 15—20 процедур. При гипотензивном типе заболевай и; электросон назначают с постепенно возрастающей частотой импульсов (10—20—40 Гц) при силе тока 7—8 мА. Продолжительность процедур — 20—50 мин. Проводят их ежедневно или 4 — 5 раз в неделю. На курс — 15—20 процедур.
Электрофорез лекарственных средств при НЦД также следует назначать дифференцированно с учетом ведущего клинического синдрома. При гипертензивном синдроме проводят электрофорез 5—10% раствора натрия или калия бромида, 5% раствора магния сульфата, 1% раствора эуфиллина, 2% раствора папаверина, 1% раствора дибазола, анаприлина (40 мг на процедуру) по методике общего воздействия либо по воротниковой методике. Электрофорез магния на воротниковую зону при гипертензивной форме заболевания проводят при силе тока 10—15 мА, время воздействия — 15—20 мин, на курс — 10 ежедневных процедур.
При гипертензивном синдроме проводят электрофорез 5—10% раствора натрия или калия бромида, 5% раствора магния сульфата, 1% раствора эуфиллина, 2% раствора папаверина, 1% раствора дибазола, анаприлина (40 мг на процедуру) по методике общего воздействия либо по воротниковой методике. Электрофорез магния на воротниковую зону при гипертензивной форме заболевания проводят при силе тока 10—15 мА, время воздействия — 15—20 мин, на курс — 10 ежедневных процедур.
При гипотензивном синдроме следует применять электрофорез кофеина. Процедуры длительностью 10—20 мин проводят при силе тока 5—7 мА через день. Курс лечения — 15 процедур. Кроме того, при гипотензивной форме заболевания проводят электрофорез мезатона на воротниковую зону или по общей методике. Сила тока — 10—15 мА, время воздействия — 15—20 мин; на курс — 10 процедур. При бессоннице, выраженных невротических проявлениях назначают электрофорез брома на воротниковую зону при силе тока 10—15 мА.
При выраженной астенизации назначают гальванический анодный воротник по Щербаку, длительность процедуры — 10-20 мин, курс лечения — 15-20 процедур через день.
Методика проведения электрофореза при помощи гальванического воротника по Щербаку: первоначальную силу тока 6 мА ежедневно увеличивают на 1 мА, в результате в конце лечения она составляет 16 мА; процедуры продлевают постепенно с 6 до 16 мин к концу курса лечения.
При выраженных проявлениях кардиалгического синдрома рекомендуется электрофорез
5 — 10% раствора новокаина, 0,5—1% раствора никотиновой кислоты преимущественно по методике общего воздействия или по кардиальной методике (электроды помещают на область сердца и в межлопаточную область). При аритмическом синдроме назначают СМТ-электро- форез анаприлина, электрофорез 5% раствора новокаина, 2% раствора панангина или анаприлина по кардиальной методике. При выраженных клинических проявлениях гипоталами- ческой дисфункции, вегетативно-сосудистых пароксизмах можно рекомендовать интраназальный электрофорез реланиума (седуксена). Курс лечения — 10 процедур.
Дарсонвализацию области сердца назначают при кардиальной форме заболевания. Время воздействия — 8—10 мин. Процедуры проводят ежедневно. На курс лечения — 10 процедур. Диадинамические токи на область шейных симпатических ганглиев рекомендуют при склонности к спазму сосудов головного мозга: катод площадью 4 см2 располагают сзади от угла нижней челюсти, анод таких же размеров помещают ниже катода на 3 см, воздействуют двух- полупериодным током в прямой полярности в течение 2 мин с одной, затем с другой стороны. Процедуры проводят ежедневно. Курс лечения — 6-10 процедур.
Время воздействия — 8—10 мин. Процедуры проводят ежедневно. На курс лечения — 10 процедур. Диадинамические токи на область шейных симпатических ганглиев рекомендуют при склонности к спазму сосудов головного мозга: катод площадью 4 см2 располагают сзади от угла нижней челюсти, анод таких же размеров помещают ниже катода на 3 см, воздействуют двух- полупериодным током в прямой полярности в течение 2 мин с одной, затем с другой стороны. Процедуры проводят ежедневно. Курс лечения — 6-10 процедур.
В настоящее время в лечении больных НЦД получает распространение аэроионотерапия. Применяют аэроионизаторы для индивидуального («Рига») и коллективного пользования (электроэффлювиальный аэроионизатор «плафон Чижевского»). В процессе ионизации воздуха образуются аэроионы отрицательного и положительного знаков с преимущественным количеством аэроионов отрицательного знака. Больные находятся на расстоянии 70—100 см от прибора, продолжительность сеанса — 20—30 мин. В течение этого времени больные вдыхают воздух, заряженный отрицательными ионами. Курс лечения —12-14 сеансов. Под влиянием аэроионотерапии снижается АД (на 5—20 мм рт. ст.), уменьшается число сердечных сокращений, повышается газообмен, увеличивается потребление кислорода, исчезает бессонница, уменьшаются головные боли, слабость.
Курс лечения —12-14 сеансов. Под влиянием аэроионотерапии снижается АД (на 5—20 мм рт. ст.), уменьшается число сердечных сокращений, повышается газообмен, увеличивается потребление кислорода, исчезает бессонница, уменьшаются головные боли, слабость.
Массаж
Положительное влияние на больных НЦД оказывает массаж, особенно общеукрепляющий. Рекомендуется щадящий массаж шейного и грудного отделов позвоночника при остеохондрозе межпозвонковых дисков этих отделов. Эффективным методом лечения НЦД в санатории Ессентуки является точечный массаж, который может выполнить сам больной или его родственники.
Санаторно-курортное лечение
Больным НЦД показано как санаторное, так и амбулаторное лечение. Санаторное лечение проводят на климатических, бальнеологических (с углекислыми, радоновыми, йодобромными водами), приморских, равнинных и горных климатических курортах, а также в местных кардиологических санаториях.
При определении вида бальнеологического курорта следует руководствоваться особенностями действия минеральных вод, а также учитывать повышенную реакцию таких больных на смену погодных и климатических условий. Следует воздерживаться от направления метеолабильных больных на курорты с климатическими условиями, резко отличающимися от таковых в местах их проживания, в сезоны с неустойчивыми погодами. Санатории Ессентуков и Кисловодска идеально подходят для больных с вегетососудистыми заболеваниями.
Следует воздерживаться от направления метеолабильных больных на курорты с климатическими условиями, резко отличающимися от таковых в местах их проживания, в сезоны с неустойчивыми погодами. Санатории Ессентуков и Кисловодска идеально подходят для больных с вегетососудистыми заболеваниями.
Санаторно-курортное в Ессентуках и Кисловодске лечение весьма благоприятно влияет на самочувствие больных НЦД. При этом используются следующие лечебные факторы: отдых, лечебное питание, климатическое и ландшафтное воздействия, минеральные воды, морские купания, бальнеолечение, физиолечение, ЛФК, терренкур, психотерапия.
Семейная медицина > Индивидуальные программы реабилитации после Covid 19
Реабилитация после Covid
19
Для многих пациентов, переболевших
коронавирусной инфекцией, возвращение к
обычной жизни проходит непросто.
Многие, даже молодые пациенты, отмечают
постоянную усталость, утомляемость,
слабость и одышку.
В «Cемейной медицине» разработаны алгоритмы, по которым создается индивидуальная
программа реабилитации после Covid 19 для каждого отдельного пациента с учетом его
особенностей, тяжести перенесенного заболевания, осложнений и наличия хронических
заболеваний.
В восстановительном периоде
используются методы лечения с применением лекарств
(витаминотерапия, антиоксиданты, сорбенты) и без применения лекарств: физиотерапия
(УВЧ-терапия, магнитотерапия, различные варианты лекарственного электрофореза,
импульсная электротерапия, массаж, иглорефлексотерапия.
Все перечисленное позволяет улучшить
микроциркуляцию в поврежденных тканях,
способствует ускорению рассасывания инфильтративных изменений и восстановлению
работы дыхательной мускулатуры, оказывает мощное противовоспалительное действие,
предотвращает возникновение спаечного процесса в легких и других органах.
Персональная программа реабилитации
создается для каждого пациента индивидуально,
при первом посещении, разрабатывается врачом-терапевтом совместно с врачом-
физиотерапевтом, курс восстановительных процедур может иметь различную
продолжительность.
Индивидуальные программы
реабилитации после Covid 19
| Консультация терапевта первичная | 1 000 ₽ |
| Консультация физиотерапевта, КМН | Бесплатно |
| Консультация терапевта повторная | Бесплатно |
Процедуры (1 сеанс)
| Рефлексотерапия | 500 ₽ |
| Электрофорез лекарственный | 160 ₽ |
| Магнитотерапия | 160 ₽ |
| Массаж спина + руки | 850 ₽ |
| Массаж спина + грудь | 950 ₽ |
| Массаж шейно-воротниковой зоны + руки | 600 ₽ |
| Кинезиотейпирование (снятие мышечных спазмов 1 зона) | 500 ₽ |
| Динамическое тейпирование (лимфодренажная аппликация 1 зона) | 600 ₽ |
| Баночный массаж, спина | 500 ₽ |
| Постуральный массаж | 400 ₽ |
| Лимфодренажный массаж | 700 ₽ |
Наши специалисты:
Тамара Давидовна Бестаева — врач — терапевт, ревматолог
⠀
Лариса Магометовна Басиева- врач- физиотерапевт, к.м.н.,
доцент кафедры медицинской реабилитации
⠀
Залина Владимировна Гиоева, врач-рефлексотерапевт, невролог
⠀
Олег Алиханович Губаев, массажист
Записаться на консультацию по вопросам реабилитации после COVID 19
можно по
тел. 8(8672) 40-42-03.
Жалобы, клинические проявления после Covid 19
Физиотерапевтический комплекс
Ожидаемый результат после прохождения программы реабилитации
Жалобы, клинические проявления после Covid 19
Длительный кашель с трудно отделяемой мокротой
Физиотерапевтический комплекс
УВЧ-терапия. Лекарственный электрофорез натрия, магния, эуфиллина,
лидазы, иода, гепарина.
Постуральный дренаж. Различные методики дыхательной гимнастики.
Классический ручной массаж, точечный, вибрационный баночный массаж.
Ожидаемый результат после прохождения программы реабилитации
Процедура УВЧ вызывает расширение капилляров легких в 3-10 раз, увеличивает газообмен в легочной
ткани, усиливает кровообращение и лимфоотток. Происходит уменьшение эксудации, отечности,
уменьшение
роста бактерий,
улучшается общее состояние. Курс 5-8 процедур.
Рекомендуемые варианты лекарственного электрофореза (который вам выберет физиотерапевт) окажут
противовоспалительное и одновременно бронхолитическое действие Назначают курс из 10-15
ежедневных
процедур.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, особенно, у
ослабленных пациентов с
сопутствующими заболеваниями сердечно-сосудистой системы, пожилых людей.
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Электрофорез эуфиллина, никотиновой кислоты, атропина. кальция, магния, иода, гепарина,
димедрола
Для активации дренажной функции показано применение синусоидальных модулированных токов (СМТ или
амплипульстерапия).
Постуральный дренаж и дренажная гимнастика.
Классический ручной массаж, точечный, вибрационный баночный массаж.
Метод волевой ликвидации глубокого дыхания К.П. Бутейко (ВЛГД).
Парадоксальная дыхательная гимнастика А.Н. Стрельниковой.
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанавливается
иммунитет.
Активизируется дренажная функция бронхов. В сочетании с лекарственным электрофорезом на область
грудной клетки устраняет спазм и улучшает отхождения мокроты.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, повышение
артериального давления
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Электрофорез никотиновой кислоты, эуфиллина, папаверина, дибазола, магния сульфат.
Различные методики дыхательной гимнастики.
Классический ручной массаж.
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанавливается
иммунитет.
Электрофорез сосудорасширяющих препаратов позволяет снизить артериальное давление, улучшить
мозговое
кровообращение
Под влиянием массажа также активизируется дренажная функция бронхов, улучшается кровообращение,
оказывается седативный эффект. В сочетании с лекарственным электрофорезом на область грудной
клетки
устраняет спазм и улучшает отхождения мокроты.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, головные боли,
головокружения, шум в голове.
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Электрофорез никотиновой кислоты по воротниковой методике, электрофорез но-шпы. Сочетанный
магний-бром электрофорез
Различные методики дыхательной гимнастики.
Классический ручной массаж.
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанавливается
иммунитет.
Электрофорез никотиновой кислоты, но-шпы позволяет улучшить мозговое кровообращение. Введение со
второго электрода р-ра брома окажет седативный эффект.
Под влиянием массажа также активизируется дренажная функция бронхов, улучшается кровообращение
органов грудной клетки, мозговое кровообращение, оказывается седативный эффект.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, нарушение сна,
раздражительность, возбуждение.
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Сочетанный магний-бром электрофорез, электрофорез трентала.
Классический ручной массаж.
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанавливается
иммунитет.
Сочетанный магний-бром электрофорез, электрофорез трентала.
Электрофорез трентала будет способствовать улучшению церебральной микроциркуляции путём снижения
тромбогенного потенциала, повышению активности фибринолитической системы, устранению
реологических
расстройств.
Под влиянием массажа также активизируется дренажная функция бронхов, улучшается кровообращение
органов грудной клетки, мозговое кровообращение, оказывается седативный эффект.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, гипертоническая
болезнь в сочетании с
атеросклерозом мозговых сосудов
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Элетрофорез магния, иода по воротниковой методике, аминалон-электрофорез по эндоназальной
методике.
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанввливается
иммунитет.
Элетрофорез магния, иода по воротниковой методике, аминолон-электрофорез по эндоназальной
методике
позволят добиться сосудорасширяющего, гипотензивного, седативного и нейротрофического эффекта
Под влиянием массажа также активизируется дренажная функция бронхов, улучшается кровообращение
органов грудной клетки, мозговое кровообращение, оказывается седативный эффект.
Жалобы, клинические проявления после Covid 19
Затруднение дыхания, одышка, чувство нехватки воздуха, нарушение
свёртывающей системы крови, с
тенденцией с гиперкоагуляцией
Физиотерапевтический комплекс
ПеМП-низкочастотное переменное магнитное поле.
Гепарин-электрофорез на воротниковую область.
Электрофорез трентала
Ожидаемый результат после прохождения программы реабилитации
Под влиянием МП происходит активация противосвертывающей системы крови,
уменьшение внутрисосудистого
пристеночного тромбообразования, достигается хороший противовоспалительный эффект,
восстанавливается
иммунитет.
Электрофорез гепарина позволит улучшить реологические свойства крови, предотвратить
тромбообразование. Электрофорез трентала будет способствовать улучшению церебральной
микроциркуляции
путём снижения тромбогенного потенциала, повышению активности фибринолитической системы,
устранению
реологических расстройств.
Электрофорез для грудничков – одно решение для множества проблем
Электрофорез для грудничков – одно решение для множества проблем
Физиотерапевтические процедуры широко применяются в педиатрии, в том числе и при лечении грудных младенцев и детей до года. Электрофорез для грудничков за десятилетия применения в медицинской практике стал одним из самых распространенных методов физиотерапии, хорошо зарекомендовавшим себя как средство против заболеваний опорно-двигательного аппарата – суставов, костей, позвоночника, нервной, дыхательной, эндокринной системы, болезней глаз и полости рта.
Список заболеваний, при которых назначают электрофорез детям до года, очень обширен.Электрофорез грудным детям чаще всего проводят для лечения гипо- и гипертонуса мышц, ДЦП, некоторых родовых травм, ОРЗ и других инфекций. Электрофорез ушей ребенку поможет справиться с хроническим отитом, от которого часто страдают ослабленные малыши, а назальный электрофорез – отличный способ лечения простудных заболеваний. Электрофорез с кальцием для ребенка по методике Вермеля – средство, которое назначают при первых признаках рахита.
Отдельно нужно обратить внимание на электрофорез с эуфиллином для грудничков – для чего его делают? В большинстве случаев электрофорез шейного отдела грудничку проводят при симптомах кривошеи, серьезного заболевания, которое лучше лечить в самом раннем возрасте. Почти всегдаэлектрофорез с эуфиллином грудничку на шейный отдел назначают именно по этой причине.Эуфиллин для электрофореза детям назначают и в качестве вспомогательного средства при введении чрескожно других медикаментов.
Электрофорез по Ратнеру также делают с применением эуфиллина и с наложением электродов на воротниковую зону. Это – отличный метод лечения врожденных или приобретенных нарушений мозгового кровообращения, повышенного внутричерепного давления.
Совместимость с другими видами физиотерапии, положительное воздействие гальванизации на организм обеспечивают самые доброжелательные на электрофорез отзывы. Дети хорошо реагируют на процедуру, так как при правильно подобранной силе тока она не доставляет никаких неприятных ощущений.
Если ребенку назначили электрофорез, следует задуматься над покупкой домашнего аппарата. Конечно, посещение физиотерапевтического кабинета может стать альтернативой покупке, но ежедневные визиты в больницу на протяжении 10-20 дней не всегда безопасны: риск заразиться от других пациентов или подхватить внутрибольничную инфекцию очень велик. Но и заниматься самолечением ни в коем случае нельзя, даже ознакомившись с тем, когда применяется электрофорез, что это такое, для детей какого возраста процедура разрешена и какие средства в каком случае применяются. В отличие от взрослого больного, ребенок не сможет сказать, что чувствует дискомфорт из-за неправильного положения накладок, вызванного неподходящим лекарством раздражения или слишком интенсивного воздействия электричества, поэтому оптимальный вариант – провести первые несколько процедур в условиях клиники под руководством врача, после чего продолжить курс лечения на дому.
Обычно электрофорез в педиатрии проводится при помощи аппаратов, обладающих плавной регуляцией силы тока и специальным детским режимом работы. Среди устройств, при помощи которых разрешено делать электрофорез для грудничков, отзывы специалистов и родителей выделяют аппарат Элфор-Проф. Простой в применении, компактный и надежный, он хорошо подходит для домашнего использования.
Еще одно популярное домашнее устройство, совпадающее с ним по большинству характеристик — аппарат ПоТок, также часто применяется для того, чтобы делать детский электрофорез. Но особого режима для самых маленьких пациентов в установках этого устройства нет – с его помощью можно проводить электрофорез на суставы, электрофорез воротниковой зоны ребенку от года и старше, а вот для того, чтобы делать электрофорез для грудничков на шею и другие зоны, ПоТок не подойдет.
Стоит заметить, что по умолчанию аппараты поставляются без необходимых для процедуры тканевых электродов. Поэтому, чтобы провести электрофорез шейного отдела ребенку, потребуется отдельно купить детские тканевые накладки или приобрести аппарат Элфор-Проф в специальной комплектации. Вместе с устройством идут три набора накладок: 30*60 мм, 50*70 мм, 60*80 мм. Самые маленькие из них идеально подходят для того, чтобы сделать электрофорез для грудничка – шейный, на область туловища, грудной клетки и суставов. А электроды большего размера можно оставить для применения в будущем.
Не только наружный могут назначить электрофорез детям. Методики лечения различных заболеваний включают в себя ванночковый и полостной электрофорез. Для лечения болезней верхних дыхательных путей и среднего уха понадобится приобрести одноразовые эндоназальные-эндонауральные электроды, которые, как и тканевые накладки, выпускаются в разных размерах и подходят для использования с аппаратом Элфор-Проф в его стандартной комплектации. Как и электрофорез на шею ребенку, полостной электрофорез нельзя проводить без рекомендации специалиста, выбора подходящих лекарств и нескольких обучающих процедур в условиях стационара. Электрофорез носовой полости очень полезен для грудничков, которые еще не могут высморкаться: после электрофореза ребенок быстро начинает нормально дышать, отек слизистых спадает после первых же двух-трех сеансов гальванизации.
Физиотерапия — cmtdr.com
ФИЗИОТЕРПИЯ
Необходимо помнить о совместимости процедур, так как у детей с ДЦП имеет место быстрая истощаемость нервных процессов, утомляемость, повышена раздражительность. Необходимо учитывать эффект последействия процедуры. Не назначается 2 электропроцедуры в один день общего воздействия, грязевые процедуры и ванны в один день; ЛФК, массаж, гидрокинезотерапия назначаются не ранее, чем через 1 час между процедурами. Курсовые лечения проводят ребенку систематически по 1.5 – 2 месяца с перерывами по 2 -3 курса в год.
Задача физиотерапии: нормализовать соотношения процессов возбуждения и торможения в центральной нервной системе, снизить ее возбудимость, усилить торможение в головном мозге, ведущие к уменьшению ригидности, спастичности мышц и интенсивности гиперкинезов, развить функциональные связи в головном мозге и выработать новые двигательные условные рефлексы, способность к произвольному торможению, способствовать развитию компенсаторных приспособлений, улучшить координацию движений и осанку.
1.Тепловые процедуры проводят с раннего возраста с последующими упражнениями, направленными на формирование достаточного объема движений в суставах, улучшения трофики спастических мышц, уменьшения ригидности.
Горячие шерстяные укутывания. Проводятся детям с 2 -3 лет.
Парафино-озокеритовые аппликации (температура 45 — 48 градусов, длительность процедуры 20 минут) или грязевые ( 38 -39 градусов) на воротниковую зону, чередуя через день с аппликациями на конечности в виде носков, перчаток, брюк (особенно при наличии контрактур).
При дизартриях ( по методике Анашкина) накладывают грязевую лепешку на слой марли вокруг рта от носа до подбородка, температура 38 -40 градусов, по 10 – 12 минут.
2. Электрофорез грязевого раствора на шейно-воротниковую зону, при явлениях гидроцефалии электрофорез по Ратнеру, на воротниковую зону электрофорез лекарственных веществ, при выраженном болевом синдроме применяется электрофорез новокаина, электрофорез новокаина по шейно- лицевой методике Келлата при гиперкинетических дизартриях, электрофорез прозерина, нивалина, галантамина, йода и лидазы на контрактуры суставов.
3. Электроанестезия по Семеновой с целью снижения афферентной импульсации из пораженных мышц: от аппаратов АСМ-3, УЭИ, «Электросон».
4. Диадинамотерапия области кистей и стоп по Келейникову,.
5. Электросон при энурезе, нервной возбудимости, нарушении сна по газнично-затылочной методике, частота подбирается индивидуально.
6. Электростимуляция от аппарата УЭИ. Амплипульстерапия у детей с целью улучшения трофики тканей спастических мышц, улучшения рецепторной проводимости имеет ряд преимуществ перед другими видами тока, так как не обладает болезненными дискомфортными ощущениями, снижают тонус спастических мышц и может назначаться с 6 месячного возраста.
7. При гиперкинетической форме применяют СМТ.
8. Диадинамические токи от аппаратов «Диадинамик».
9. Индуктотермия назначается детям ДЦП с целью глубокого прогревания тканей, усиления кровообращения, уменьшения болевого синдрома.
При легкой степени спастичности мышц детям дошкольного возраста назначается воздействие индуктором от аппарата «УВЧ-30» на область нижнешейных и верхнегрудных позвонков .
21. Для лечения детей школьного возраста с диплегией используют воздействие от ИКВ-4 индуктором-кабелем последовательно на два поля:
1 поле – паравертебрально от уровня С6 до L2, дозировка слаботепловая, по 10 минут. Затем :
2 поле – обе ноги по 6 – 9 минут на каждую сторону, ежденевно, № 10.
В тяжелых случаях сначала проводят индуктотермию паравертебрально № 10, а затем на ноги № 10.
После процедуры через 30 минут – 1 час проводят лечебную физкультуру.
Комбинация индуктотермии с радоновыми, углекисло — сероводородными ваннами дает выраженный продолжительный эффект.
10. Микроволновую терапию (СМВ и ДМВ) хорошо назначать детям с парапарезом, выраженными гиперкинезами, болезненными явлениями.
11. У детей старшего возраста для лечения контрактур применяют ультразвуковую терапию от аппарата УЗТ- 5.
12. Виброакусическое воздействие от аппарата «Витафон» детям школьного возраста.
13. Лазеротерапия оказывает спазмолитический, сосудорасширяющий, улучшающий местное кровообращение эффекты. Способы воздействия – накожно, внутрисосудистое облучение крови, лазеропунктура.
Целесообразно сочетание и проведение в один день процедур накожного воздействия по соответствующим полям и областям с внутрисосудистым облучением крови. Комбинирование воздействий применяют по принципу чередования процедур накожного облучения и лазеропунктуры по разным дням.
14. КВЧ терапия проводится по аналогичной схеме лазеропунктуры.
Электрофоретический анализ протеаз в Echinostoma caproni и Echinostoma trivolvis на JSTOR
Abstract
Электрофорез в полиакриламидном геле с додецилсульфатом натрия на желатиновой подложке был использован для анализа протеаз у взрослых особей Echinostoma caproni и Echinostoma trivolvis в возрасте 14 дней. При pH 8,0 взрослые особи E. caproni показали 2 полосы протеазы 36 кДа и 58 кДа, тогда как взрослые особи E. trivolvis показали 6 полос 39, 64, 77, 96, 120 и 168 кДа. Каждый вид также демонстрировал различные паттерны образования протеаз в своих экскреторных / секреторных (E / S) продуктах.E / S протеазы E. caproni имели 36 и 58 кДа, тогда как протеазы E. trivolvis были 120 и 168 кДа. Дальнейшая характеристика протеаз взрослых особей E. caproni выявила 2 полосы (58 и 66 кДа) с оптимальной активностью при pH 3,0-4,5 и 3 полосы (38, 61 и 96 кДа), которые были наиболее активными при pH 7,0-8,0. Четыре низкомолекулярные полосы (19, 21, 25 и 30 кДа) появляются, когда экстракты червя E. caproni инкубируются в присутствии CaCl 2 при pH 8,0, но ингибируются этилендиаминтетрауксусной кислотой и 1,10-фенантролином.Полосы протеазы Echinostoma caproni при 58 и 38 кДа во всех образцах червя и продукты E / S и полоса 36 кДа во всех образцах червя ингибировались фенилметилсульфонилфторидом. Показывая различия протеаз в дополнение к недавним исследованиям нуклеотидных различий, это исследование помогает различить эти 2 родственных аллопатрических вида эхиностомы с 37 воротниковыми шипами.
Информация о журнале
Журнал паразитологии — официальное издание Американского общества паразитологов (ASP).Он издается непрерывно с 1914 года, когда его основал Генри Болдуин Уорд. Журнал специализируется на общей паразитологии, а также на паразитах, имеющих медицинское, ветеринарное и экономическое значение. Основное внимание уделяется эукариотическим паразитам, хотя векторы для прокариотических организмов и вирусов также включены. Журнал печатается в издательстве Allen Press в Лоуренсе, штат Канзас, шесть раз в год. Доступны институциональные подписки. Членство в ASP включает получение журнала Parasitology, ежеквартального информационного бюллетеня и выдержек из ежегодных собраний; студенты могут присоединиться по значительно сниженной ставке.Журнал находится в сети через BioOne. Редактор журнала — д-р Майкл В.К. Сухдео. Для получения дополнительной информации о публикации в журнале обращайтесь в редакцию журнала по адресу [email protected]. Для получения информации о членстве в ASP свяжитесь с обществом: http://amsocparasit.org/. Для получения информации о подписке обращайтесь на [email protected].
Информация об издателе
Allen Press играет жизненно важную роль в распространении знаний и информации.
путем партнерства с организациями в научном, техническом и медицинском сообществе
продвигать свои интересы и добиваться поставленных целей.Штаб-квартира в Лоуренсе,
KS, Allen Press предоставляет полный спектр интегрированных услуг в
широкие области управления контентом, онлайн-доставка и печать, ассоциация
менеджмент и издательские услуги. Allen Press — надежный партнер научных
общества, профессиональные ассоциации и корпорации по всей стране.
Для идентификации и количественного анализа этих веществ классические методы, такие как: ПЦР, […] eitplus.pl | Do identityfikacji alkaloidów psychoaktywnych zastosowane zostaną […] eitplus.pl |
| ВЭЖХ a n d капиллярный электрофорез m e th ods используются […] для анализа биологически активных соединений. pan.olsztyn.pl | W analityce wykorzystuje się wysokosprawną […] refresh.pan.olsztyn.pl |
OENO-SCMA 08-385, определение […] eur-lex.europa.eu | OENO-SCMA 08-385, oznaczanie zawartości lizozymu w […] eur-lex.europa.eu |
Динамическое рассеяние света, […] ichf.edu.pl | reom etr ii , электрофорезы ż e lowej i kapi larn ej , хроматография […] w połączeniu z metod Taylora. ichf.edu.pl |
| Практический аспект s o f капиллярный электрофорез u t il ization chem.ug.edu.pl | praktyczne aspekt y wykor zys tan ia elektroforezy kapilarnej chem.ug.edu.pl |
На кафедре проводятся исследования биологически активных […] соединений, таких как фенольные кислоты, дубильные вещества, лигнаны и катехины, используя […] pan.olsztyn.pl | Zakład rozwija analitykę związków biologicznie aktywnych (фенолоквасы, танины, лигнаны, катехины) […] pan.olsztyn.pl |
| utilizatio n o f капиллярный электрофорез ( C E) для идентификации […] веществ с разными химическими свойствами chem.ug.edu.pl | wy korz ysta ni e elektroforezy kapilarnej (C E) do id en tyfikacji […] związków o różnym charakterze chemicznym chem.ug.edu.pl |
ПЦР-продуктов […] вместе с маркерами размера и положительным контролем и […] отрицательные контроли после стадий RT и ПЦР. eur-lex.europa.eu | Продукты PCR po ddaj e si ę elektroforezie w 1% żel u agarowym, […] wzdłuż znaczników rozmiaru wzdłuż i kontroli dodatniej oraz kontroli ujemnych z RT i PCR. eur-lex.europa.eu |
В частности, усиление […] в комнате отдельно от отделения ДНК, […] Приготовление смеси для ПЦР и выдача ДНК. eur-lex.europa.eu | Przede wszystkim […] odbywać się w innym pomieszczeniu ni ekstrakcja […] ДНК, предварительное тестирование реактивной ПЦР и дозирование ДНК. eur-lex.europa.eu |
Применяемая методология основана на полимеразной цепной реакции (ПЦР) и включает отбор проб, […] eur-lex.europa.eu | Методология застосованья опарта на […] reakcji łańcucha polimerazynowego (PCR) и obejmuje […] elektroforezężelową. eur-lex.europa.eu |
Контроль качества и эффективности […] экстракции (например, путем измерения OD (260 нм) ниже […] eur-lex.europa.eu | Kontrolować jakość i skuteczność ekstrakcji (na przykład, mierząc OD (260 nm) za pomocą […] eur-lex.europa.eu |
Разложите переваренные фрагменты […] Соблюдайте характерный фрагмент ограничения […] при УФ-трансиллюминации после окрашивания бромидом этидия и сравнить с непереваренным и расщепленным положительным контролем. eur-lex.europa.eu | Rozdzielić соломионе […] jak wcześniej i oglądać charakterystyczny układ […] fragmentów Strawionych przez фермент restrykcyjne w transiluminatorze UV po uprzednim barwieniu bromkiem etydyny. eur-lex.europa.eu |
Когда гель полимеризуется, гребни удаляются и гель помещается […] Достаточно 1 X TAE для покрытия агарозного геля. eur-lex.europa.eu | Po polimeryzacji żelu usuwa się grzebienie, a el […] wystarczająco 1X TAE, aby pokryć żel agarozowy. eur-lex.europa.eu |
Пример напряжения […] × 4,5 см), электроды без полосок наносили непосредственно на гель. eur-lex.europa.eu | 6.3.2. Przykład programu napięcia elektrycznego dla […] см), elektrody bez pasków stosowane bezpośrednio na elu. eur-lex.europa.eu |
(A) неэтерифицированное оливковое масло, после […] липаза; после силанизации; под этими […] элюируется одновременно с […] фракция диглицеридов или немного позже. eur-lex.europa.eu | A) nie estryfikowanej oliwy z oliwek, po […] zadziałaniu na nią lipazą; по силилованиу; w […] м) frakcja wosków ulega elucji równocześnie […] z frakcją diglicerydów lub nieco później. eur-lex.europa.eu |
Этот метод подходит для определения транс-анетола в ароматизированном анисом […] eur-lex.europa.eu | Metoda ta jest odpowiednia do oznaczania transanetolu w napojach alkoholowych z […] gazowej. eur-lex.europa.eu |
Область глаз перекрывает […] минимальная жировая набивка, что делает его склонным к отечности и темным кругам. amway.pl | Pod skórą wokół oczu znajduje […] oraz znikoma wyściółka tłuszczowa, co czyni ją […] szczególnie podatną na opuchliznę i powstawanie ciemnych plam. amway.pl |
В комплекте: муфты латунные […] устойчив к высоким температурам, изоляция для муфт и латунь […] датчик температуры. Вт / пл | W skład zestawu wchodzą: złączki […] mosiężne odporne na wysokie temperatury, izolacje na złączki, […] температуры коллектора. Вт / пл |
| Капилляр w i th протестированная кровь […] (поз. № 2 на рис. № 1.) следует поместить в канал на индикаторе так, чтобы […] следующий столбец крови совпадает с горизонтальной линией на индикаторе ползунка. Плечо (поз. № 3 на рис. № 1) должно быть установлено так, чтобы разрезанная на нем линия совпадала с верхним уровнем столба крови. mpw.pl | Kapilarę z o dwi row aną p róbką krwi […] (элемент 2 на Рыс. 1) połoyć w kanałku wskaźnika, tak aby dolny poziom prokrywał […] się z poziomą linią na suwaku, linię na ruchomym przeźroczystym ramieniu (element 3 na Rys. 1) pokrywamy z górnym poziomem wypełnienia próbki, następnie suwöwak zakmł osocza. mpw.pl |
Электрофокусирующая камера с охлаждающей пластиной (например, 265 × 125 мм) и подходящей мощностью […] eur-lex.europa.eu | Komora elektroogniskowania z płytą […] chłodzącą (na przykład 265 × 125 мм) oraz odpowiedni układ zasilania (≥ 2,5 […] eur-lex.europa.eu |
Полиакриламидный гель можно разрезать до меньшего размера […] размеров, рекомендуется при малом количестве […] геля размером 4,5 × 5 см). eur-lex.europa.eu | Żel poliakryloamidowy możebyć pocięty na mniejsze części, co jest zalecane w […] przypadku, kiedy jest kilka probek lub kiedy użyte jest […] × 5 см). eur-lex.europa.eu |
Контроль успешного […] не обязательно. eur-lex.europa.eu | Zalecana jest kontrola pomyślnej […] ona obowiązkowa. eur-lex.europa.eu |
Исследовательская группа имеет две основные методические специализации по химии: методы разделения с […] особый упор на хиральный […] на основе дифракционных методов. ichf.edu.pl | Grupa badawcza dysponuje dwiema głównymi specjalizacjami metodycznymi. Są по адресу: techniki […] rozdzielania, w szczególności z […] Badania Strukturalne Oparte o Techniki Dyfrakcyjne. ichf.edu.pl |
Однако, в отличие от других методов, основанных на ПЦР, методология SSP, используемая компанией One Lambda, Inc, различает различные полиморфизмы во время процесса ПЦР.4 Это сокращает […] обработка после усиления […] результаты либо положительные, либо отрицательные. onelambda.com | Odmiennie niż w przypadku innych metod opartych na PCR, metodologia SSP wykorzystana przez firmę One Lambda, Inc.pozwala na rozróżnianie odmiennych poliformizmów podczas processu PCR.4 […] Skraca to czas etapu poamplifikacyjnego […] вики są albo pozytywne albo negatywne. onelambda.com |
После завершения продукты ПЦР […] eur-lex.europa.eu | Po zakończeniu produkty PCR […] Вт, температура 4 ° C, смазка — 20 ° C. eur-lex.europa.eu |
Фрагменты (ампликоны) ПЦР […] eur-lex.europa.eu | Узискане фрагменты ПЦР (субстанция wzmacniających) mają być […] eur-lex.europa.eu |
По механическому составу это обычно рыхлые пески с массовой плотностью в среднем 1,48 г / см-3, содержанием органических веществ 0-5%, […] содержание гигроскопичной воды 0,41% по […] (в среднем 25,4% от веса) и не более […] обводненности (28,4% веса). databases.eucc-d.de | Pod względem składu mechanicznego są to przeważnie piaski lune, wykazujące gęstość objętościową średnio 1,48 г · см-3, zawartość materii Organicznej 0-5% и zawartość . wody Higroskopowej średnio 0,41%, a ponadto […] 25,4% ваг.) Или максимальна (28,4% ваг.). databases.eucc-d.de |
| Если скорость т ч e капилляр a c ti уменьшается, то высота подъема воды также уменьшается, поскольку на нее сильно влияет испарение [.. .] л. Воды. hahne-bautenschutz.de | Kiedy prę dk ość kapilarnego trans po rtu wody jest mniejsza, zmniejsza się również wysokość podciąga ni a kapilarnego w18 …] oddziałuje na nią silnie […] szybkość parowania dyfuzyjnego. hahne-bautenschutz.de |
| В ближнем фу tu r e капилляр m e mb ranes может быть важным […] экспортный товар. pi.gov.pl | W najbliższej […] польским товаром экспортным. pi.gov.pl |
| T h e капиллярный a c ti возможно, потому что […] сосуды узкие. anthura.nl | Z ja wis ka kapilarne są m ożli we , gdyż naczynia […] są wąskie. anthura.nl |
USDA FSIS переходит на полногеномное секвенирование бактерий
понедельник, 17 декабря 2018 г.
Секвенирование всего генома (WGS) определяет порядок всех строительных блоков ДНК (нуклеотидов) во всем геноме организма в одном лабораторном процессе.Сравнение последовательности ДНК изолированного бактериального патогена с последовательностями из других образцов в базе данных ДНК может точно определить источник вспышки болезни пищевого происхождения. Аналитические инструменты для WGS становятся все более точными, простыми и доступными, так что учреждения здравоохранения перешли на WGS в качестве предпочтительного инструмента для исследования болезней пищевого происхождения от Listeria , Salmonella и патогенных штаммов Escherichia coli , включая Э.coli O157: H7.
Как сообщается здесь, Служба безопасности и контроля пищевых продуктов (FSIS) Министерства сельского хозяйства США (USDA) провела встречу, чтобы обсудить состояние дел и планы FSIS по сбору и анализу данных WGS для бактерий, выделенных из официальных образцов. В 2017 финансовом году все бактериальные патогены, выделенные в рамках программ тестирования FSIS, были проанализированы параллельно с использованием гель-электрофореза в импульсном поле (PFGE) и WGS. В официальном обновлении FSIS (14 декабря 2018 г.) сообщается, что FSIS приостановит рутинный анализ PFGE и перейдет на использование только WGS для продуцирующего токсин шига E.coli (STEC), вступающий в силу 15 января 2019 г., и для изолятов Salmonella , вступающий в силу 15 марта 2019 г.
FSIS ежедневно проверяет почти 6 500 предприятий и регулярно собирает очень большие объемы микробных проб. Например, в рамках программы FSIS по микробиологическому тестированию готового к употреблению мяса и птицы (RTE) в 2017 году FSIS собрала и проанализировала 6892 образца пищевых продуктов RTE на сальмонеллу и Listeria monocytogenes (Lm) и проанализировала 4614 образцов, полученных от контакта с пищевыми продуктами. поверхности для Lm.По мере расширения технологии WSG будет становиться все более ценным инструментом для быстрого отслеживания болезней до их источника и, возможно, когда-нибудь достаточно раннего выявления патогенов в окружающей среде, чтобы полностью предотвратить загрязнение пищевых продуктов.
© 2021 Keller and Heckman LLP, National Law Review, Volume VIII, Number 351
Выше воротника и ниже пояса
Лимфомы мочевого пузыря редко являются первичными опухолями и чаще связаны с системной лимфомой, либо как нелокализованная лимфома мочевого пузыря, либо как вторичная лимфома мочевого пузыря.Первичные лимфомы мочевого пузыря (PBL), как правило, относятся к типу лимфоидной ткани, ассоциированной со слизистой оболочкой (MALT) низкой степени, в отличие от диффузных крупноклеточных клеток или клеток фолликулярного центра, которые чаще встречаются при вторичной лимфоме мочевого пузыря. Поражение мочевого пузыря системной лимфомой предполагает плохой прогноз, и у пациентов часто отсутствуют локализующие симптомы (обычно это патологоанатомический диагноз). Другие методы лечения предпочтительнее хирургического лечения всех лимфом мочевого пузыря, за исключением случаев, когда диагноз не определен или для облегчения симптомов раздражения мочевого пузыря.Мы описываем уникальный случай системной B-клеточной лимфомы высокой степени с одновременным поражением кожи почек и мочевого пузыря при обращении.
1. История болезни
70-летний мужчина обратился к дерматологам с двухлетним анамнезом безболезненных увеличивающихся узелков на коже черепа, которые быстро ухудшились за предыдущие 6 месяцев (рис. 1 (а)). Он похудел на 7 кг; других системных симптомов не было. У него не было ни лимфаденопатии, ни гепатоспленомегалии, а его анализ периферической крови, СОЭ, электрофорез белков, функциональные пробы печени и ЛДГ были в норме.Биопсия кожи волосистой части головы показала В-клеточную неходжкинскую лимфому высокой степени злокачественности. Постановочная КТ показала множественную небольшую шейную лимфаденопатию, лимфоматозную инфильтрацию в обеих почках и область утолщения слизистой оболочки мочевого пузыря, что соответствовало первичной карциноме мочевого пузыря. Постановка была завершена с помощью МРТ головы (рис. 1 (b)) и биопсии костного мозга, которая была нормальной.
Пациент не имел симптомов со стороны нижних мочевыводящих путей, эпизодов макроскопической гематурии и не курил.Цитологический образец мочи выявил атипичные клетки, более соответствующие лимфоидному, чем эпителиальному / переходному клеточному происхождению (рис. 2 (а)). При цистоскопии было замечено и удалено подслизистое поражение 3 см на передней поверхности стенки мочевого пузыря. Гистология подтвердила В-клеточную лимфому высокой степени злокачественности при биопсиях мочевого пузыря и почек (рис. 2 (b)).
2. Обсуждение
На сегодняшний день имеется несколько сообщений о случаях не первичной лимфомы мочевого пузыря, что отражает ее редкость как патологического образования.Насколько нам известно, мы описываем первый зарегистрированный случай диссеминированной лимфомы с одновременным поражением кожи, почек и мочевого пузыря, очевидным при обращении.
Заболеваемость неходжкинской лимфомой (НХЛ) растет и сохраняет географическую изменчивость: в 2007 году в Великобритании было 10 900 человек с диагнозом НХЛ [1]. Заболеваемость НХЛ обычно увеличивается с возрастом. Показатели резко возрастают у людей старше 50 лет, и около двух третей (68%) всех случаев диагностируются у людей старше 60 лет.Самая последняя система классификации одобрена Всемирной организацией здравоохранения (классификация REAL) и включает такие подтипы, как диффузные большие B-клетки, фолликулярные и малые лимфоциты. Определение конкретных подтипов заболевания наряду с прогностическими характеристиками пациента, полученными с помощью Международного прогностического индекса, имеет решающее значение для планов лечения и последующего ведения.
Лимфомы мочевого пузыря встречаются редко и составляют 0,2% всех новообразований мочевого пузыря [2]. Преобладают женщины, средний возраст обращения — 58 лет.Наиболее распространенным типом является лимфома экстранодальной маргинальной зоны слизистой лимфоидной ткани (MALT), впервые описанная в 1990 году [3], тогда как тип Ходжкина встречается еще реже. Лимфомы классифицируются в первичную группу, если они проявляются в основном симптомами мочевого пузыря и при этом нет признаков системной лимфомы.
Чаще, хотя существует относительно небольшое количество тематических исследований, лимфома мочевого пузыря встречается вместе с системной лимфомой; это может быть либо поражение мочевого пузыря с активной системной лимфомой (нелокализованная лимфома мочевого пузыря), как описано в этом исследовании, либо рецидив лимфомы в мочевом пузыре после периода ремиссии от предыдущей лимфомы (вторичная лимфома) [3].Диссеминированная лимфома редко проявляется симптомами поражения мочевого пузыря и, как правило, остается посмертным диагнозом [2]. Хотя у некоторого процента пациентов с поражением мочевого пузыря нет симптомов мочеиспускания, были описаны срочность, частота мочеиспускания и гематурия [3]. Сообщается, что большая часть (47%) пациентов в этой группе испытывает обструкцию мочеточника [3].
Непервичные лимфомы мочевого пузыря, как правило, имеют гистологический тип, отличный от тех, которые обнаруживаются в группе PBL, и обычно относятся к типам диффузных крупных клеток или клеток фолликулярного центра [3].В непервичной группе поражение мочевого пузыря системной лимфомой предполагает худший прогноз, чем в PBL, поскольку это подразумевает мультисистемное заболевание. Однако существует четкое прогностическое различие между группами вторичной лимфомы мочевого пузыря и нелокализованной лимфомы со средней выживаемостью 0,6 и 9 лет соответственно [3]. Частично это можно объяснить тем, что некоторые пациенты были отнесены к нелокализованной группе, когда доминирующая масса мочевого пузыря была связана только с локальным распространением на соседние органы или региональные лимфатические узлы [3].Авторы утверждали, что, хотя некоторые могут классифицировать эту категорию пациентов в группу «PBL», их гистология опухолей больше напоминала таковую в общей популяции, а характеристики пациента и заболевания отличались от таковых в группе PBL [3].
Неходжкинскую лимфому с поражением мочевого пузыря можно успешно лечить с помощью химиотерапии, лучевой терапии, иммунотерапии, хирургии и сочетания этих методов [4]. Выбор лечения определяется гистологией и степенью заболевания.Системный характер не первичной лимфомы мочевого пузыря требует системного подхода к лечению. Лимфомы с агрессивными фенотипами, такими как диффузные большие B-клетки, лучше всего лечить с помощью химиотерапии. Чаще всего используется режим циклофосфамида, доксорубицина, винкристина, преднизона и ритуксимаба (CHOP / R) [4]. Адъювантная лучевая терапия используется для пораженных областей при локализованном заболевании. Лечение фолликулярной лимфомы варьируется от политики наблюдения и ожидания до химиоиммунотерапии или радиоиммунотерапии. Иммунотерапия обычно является стандартным компонентом лечения запущенного заболевания [4].
Вялотекущий характер лимфомы MALT означает, что местная лучевая терапия может использоваться с лечебной целью, например, при PBL. Хирургическая резекция играет ограниченную роль при первичной лимфоме мочевого пузыря, помимо получения образцов биопсии, и редко что-либо добавляет к лечению или прогнозу. Однако есть сообщения о пациентах, перенесших цистэктомию из-за неясного диагноза или для облегчения симптомов раздражения [5]. Имеется сообщение о ЛПН низкой степени злокачественности MALT-типа, которое лечится с помощью трансуретральной резекции и адъювантного внутрипузырного введения митоксантрона [6].
В этом случае болезнь пациента составляла стадию IV B. Он прошел восемь курсов химиотерапии CHOP / R с последующей радикальной лучевой терапией кожи головы и головного мозга. Последующая МРТ показала резкое улучшение скальпа и образования твердой мозговой оболочки с минимальным остаточным усилением твердой мозговой оболочки. Повторная компьютерная томография показала полную ремиссию без остаточного поражения почек или мочевого пузыря. Пациент оставался в полной ремиссии в течение 14 месяцев, но затем у него появилось постепенное ухудшение баланса и, как следствие, выраженная периферическая невропатия.Повторная визуализация подтвердила рецидивирующие широко распространенные церебральные метастазы. Он умер после остановки сердца до того, как ему была назначена паллиативная лучевая терапия.
Авторское право
Авторское право © 2012 Эрик К. Майер и др. Это статья в открытом доступе, распространяемая по лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
(PDF) Гниль ошейников и корней оливковых деревьев, вызываемая Phytophthora megasperma на Сицилии
Январь 2001 г., Том 85, номер 1
Page 96
http: // dx.doi.org/10.1094/PDIS.2001.85.1.96A
Примечания к болезни
Гниль ошейников и корней оливковых деревьев, вызванная
Phhytophthora megasperma на Сицилии
SO Cacciola, Istituto di Patologia Vegetale, Италия, Университет Палермо, ; и G.
Э. Агостео и Дж. Маньяно ди Сан-Лио, Dipartimento di Agrochimica ed Agrobiologia, Университет Реджо
Калабрия, 89061 Галлина ди Реджо Калабрия
Открытый доступ.
Оливка (Olea europea L.) выращивается примерно на 154 000 га в Сицилии (юг Италии). Летом 1999 г. в недавно посаженном коммерческом саду в провинции
Энна (Сицилия) было обнаружено несколько оливковых деревьев возрастом 3-
лет с симптомами упадка. Размножали деревья на подвое дикорастущих оливковых (O. europea L. var. Sylvestris Brot.).
Воздушные симптомы, включающие хлороз листьев, увядание, дефолиацию и отмирание веток, за которыми в большинстве случаев следовало
гибели растений, были связаны с корневой гнилью и язвой базальных стеблей.Phytophthora sp. был последовательно выделен
из сгнивших корешков и язв ствола с использованием селективной среды BNPRAH (беномил, нистатин,
пентахлорнитробензол, рифампицин, ампициллин и гимексазол). Чистые культуры
были получены переносом одной гифы. Вид, выделенный из симптоматических оливковых деревьев, был идентифицирован как P.
megasperma Drechsler на основе морфологических и культурных характеристик. Все изоляты были гомоталлическими,
с парагинными антеридиями.Диаметр ооспор варьировал от 28 до 42 мкм (среднее ± SE = 36,3 ± 0,4)
, когда они были получены на картофельно-декстрозном агаре (PDA), и от 30 до 43 мкм (среднее ± SE = 37,8 ± 0,4)
когда они были произведены в физиологическом растворе. Спорангии не сосочковые. Оптимальные и максимальные
температуры для радиального роста колоний на КПК составляли 25 и 30 ° C соответственно. При 25 ° C скорость радиального роста
составляла около 6 мм в сутки. Идентификация была подтверждена электрофорезом мицелиальных белков
на полиакриламидном пластинчатом геле.Диаграммы электрофоретических полос общих растворимых белков и
трех изоферментов (эстераза, фумараза и малатдегидрогеназа) изолята оливы были идентичны таковым для
двух изолятов P. megasperma, полученных из вишни и моркови в Италии и охарактеризованных
ранее (1). Напротив, они четко отличались от электрофоретических паттернов четырех изолятов P.
megasperma var. sojae Hildebr. из сои (= P. sojaeKauf.& Ger.), От таковых из трех изолятов спаржи
, предварительно идентифицированных как P. megasperma sensu lato (1), и от изолятов контрольных изолятов
различных видов, продуцирующих не сосочковые спорангии, включая P. cambivora (Petri) Buisman , P.
cinnamomiRands, P. cryptogea Pethybr. & Laff., P. drechsleri Tucker и P. erythroseptica Pethybr. Патогенность
изолят оливы тестировали в теплице при температуре от 18 до 25 ° C на 18-месячных укорененных черенках оливы
cv.Бианколилла. Черенки инокулировали на нижний стебель, вставляя 3-миллиметровую пробку, взятую из активно растущих колоний
на КПК, в разрез, сделанный стерильным скальпелем. Рану заклеили водостойкой лентой
. Пробки агара без мицелия помещали в стебель черенков, используемых в качестве контроля. Кора была удалена на
, и области повреждений были отслежены и измерены через 60 дней после инокуляции. Изолят оливы
образовал коричневое некротическое поражение (средний размер = 500 мм2) вокруг инокуляционной раны и был повторно изолирован
от очага поражения.На контрольных растениях рана, наоборот, зажила. P. megasperma ранее был
VIVID взаимодействует с комплексом WHITE COLLAR и FREQUENCY-взаимодействующей РНК-геликазой для изменения световых и часовых ответов у Neurospora
Abstract
Фоторецептор и белок PAS / LOV VIVID (VVD) модулируют передачу сигналов синим светом. и влияет на световые и температурные реакции циркадных часов у Neurospora crassa . Одним из основных действий VVD на циркадные часы является влияние на фазу циркадных часов путем регулирования уровней транскриптов, кодируемых центральным часовым геном , частотой ( frq ).Как достигается это регулирование, неизвестно. Здесь мы показываем, что VVD взаимодействует с комплексами, центральными для циркадных часов и передачи сигналов синего света, а именно с комплексом WHITE-COLLAR (WCC) и FREQUENCY-взаимодействующей РНК-геликазой (FRH), компонентом, который объединяется с FRQ, чтобы опосредовать контроль отрицательной обратной связи у Neurospora. . VVD взаимодействует с FRH в отсутствие WCC и FRQ, но, по-видимому, не контролирует опосредованную экзосомами петлю отрицательной обратной связи. Вместо этого VVD действует, чтобы модулировать транскрипционную активность WCC.
Свет, помимо того, что является источником энергии для многих форм жизни на Земле, действует как сигнал, который может запускать развитие, или как повторяющийся сигнал, отмечающий прохождение внешнего времени. Внешние временные сигналы используются клеточными таймерами, такими как циркадные часы, для привязки их периодов к времени внешнего дня. Процесс блокировки периода называется «увлечением» и гарантирует, что клеточная и поведенческая активность происходит в то время суток, когда их адаптивная ценность наиболее высока (1–3).Синий свет играет центральную роль в увлечении циркадных часов. В самом деле, фоторецепторы синего света и циркадные часы, возможно, одновременно произошли от механизма, который изначально служил для обнаружения (фоторецептора) и предотвращения (таймер) вредного излучения (4–6). Наше понимание молекулярных основ циркадных часов и их реакции на свет резко улучшилось за последнее десятилетие или около того, и эукариотический модельный организм Neurospora crassa стал одной из наиболее изученных систем для понимания обоих процессов (7-9 ).
Ключевые компоненты циркадных часов Neurospora являются продуктами white collar ( wc-1 и wc-2 ), частоты ( frq ) и frq — взаимодействующей геликазы ( frh) генов (4, 10, 11). Фоторецептор синего света WC-1 и его партнер по взаимодействию WC-2 образуют транскрипционно и фотоактивный комплекс WHITE COLLAR (WCC), который активирует экспрессию frq (4, 12). Белок FRQ, в свою очередь, объединяется с FRH с образованием комплекса FRQ-FRH (FFC), который подавляет активность WCC (9, 11).Таким образом, фоторецепция и временная организация экспрессии генов связаны через WCC (4, 12–14). Гиперфосфорилированный WCC транскрипционно менее активен, и репрессия WCC с помощью FRQ происходит посредством FRQ-опосредованного фосфорилирования WCC с помощью казеинкиназы 1 и 2 (CK1 и 2) (14, 15).
Вторая петля обратной связи, которая подавляет активность WCC, включает продукт гена vivid ( vvd ) (16). Подобно WC-1, VVD представляет собой белок PAS / LOV и фоторецептор синего света; однако, в отличие от WC-1, его присутствие не является существенным для циркадной ритмики в постоянной темноте (DD) (16-19).Тем не менее, VVD играет важную роль в циркадной системе Neurospora. Без ВВС организм более чувствителен к свету, что приводит к быстрому нарушению циркадной организации при постоянном освещении, тогда как при наличии ВВС временная ритмичность сохраняется при постоянном освещении (LL). Влияя на сброс часов как на рассвете, так и в сумерках, VVD влияет на сброс света и увлечение циркадных часов (16, 20). Наконец, VVD играет роль в поддержании правильной синхронизации управляемых часами выходных путей при различных температурах (21).
Вне циркадной системы VVD играет ключевую роль в фотоадаптации (18, 22, 23). VVD временно димеризуется светозависимым образом (24–26), и это конформационное изменение может передавать световые сигналы нижестоящим мишеням. Однако, несмотря на важность VVD для согласования часов Neurospora с внешним днем, мы до сих пор не знаем, как VVD выполняет эти действия на молекулярном уровне. Партнеров по взаимодействию ВВД (кроме самого ВВД) не выявлено, и, следовательно, отсутствует понимание механизмов действия ВВД.Здесь мы показываем, что VVD взаимодействует с часовыми компонентами WC-1 и FRH. Наши данные предполагают, что VVD действует в ядре как FRH-зависимый корепрессор WCC.
Результаты
VVD регулирует
frq уровней транскрипции в сумерках.
Молекулярные и физиологические данные показали, что ВВД влияет на сброс часов на рассвете и в сумерках. В сумерках влияние VVD на молекулярные события становится очевидным при сравнении уровней мРНК frq WT и vvd -нокаут ( vvd ko ) штаммов.Транскрипт frq остается повышенным дольше в штаммах vvd ko , чем в WT, с задержкой примерно на 4 часа в достижении базальных уровней (две верхние дорожки на рис. 1 A ) (16, 20). Чтобы получить более прямые доказательства роли VVD в регуляции уровней транскрипта frq , мы создали штамм, в котором индуцируемая хинной кислотой (QA) копия гена vvd , меченного эпитопом myc (qa- vvd myc ) был вставлен в локус his-3 .Конструкция qa- vvd myc была интегрирована в штаммы WT или vvd ko , и в дальнейшем эти штаммы упоминаются как qa- vvd myc (WT) и qa- vvd 917 ( ввд КО ) соответственно. Используя эту стратегию, мы смогли отделить экспрессию vvd от его нормальной световой регуляции (фиг. 1 B и фиг. S1). Действительно, эктопическая экспрессия VVD, вызванная добавлением QA в qa- vvd myc ( vvd ko ), восстанавливает нормальное снижение уровней frq (сравните образцы -QA и + QA на рис. .1 A , четвертая панель) и ускоряет деградацию frq в qa- vvd myc (WT) сверх того, что наблюдается в нормальном WT (сравните образцы -QA и + QA в первой и третьей дорожках на рис. 1 A и см. Количественную оценку данных на рис. S1 A ). Наблюдение, что уровни frq несколько ниже в среде QA, было ожидаемым, поскольку полная экспрессия frq зависит от глюкозы (27). Взятые вместе, эти данные иллюстрируют обратную корреляцию между уровнями транскрипта frq и белком VVD.
Рис. 1.
VVD контролирует frq уровней РНК. ( A ) Нозерн-блоты, показывающие уровни транскрипта frq в штаммах vvd + (WT) или vvd ko с эктопической вставкой QA-индуцибельного гена vvd или без него. Культуры выращивали в LL в течение 24 часов в присутствии или в отсутствие индуктора QA перед переносом в DD, и образцы собирали через 24 часа в LL (момент времени 0) или в указанные моменты времени в DD.( B ) Как и в случае A , но Нозерн-блоты гибридизовали с зондом, обнаруживающим транскрипт vvd . Контроль загрузки и количественный анализ показаны на рис. S1.
VVD Взаимодействует с FRH.
Поскольку FFC играет ключевую роль в отрицательной обратной связи frq (11, 28, 29), возможно, что VVD напрямую модулирует активность этого комплекса, чтобы влиять на уровни frq в сумерках. Чтобы проверить, взаимодействует ли VVD с FFC, мы провели эксперименты по коиммунопреципитации (Co-IP) на лизатах цельных клеток Neurospora с использованием антисывороток FRH или FRQ соответственно.Чтобы облегчить обнаружение VVD, мы использовали штамм, который экспрессирует MYC-tagged VVD на фоне vvd ko . Ранее мы показали, что этот штамм спасает все известные мутантные фенотипы vvd , тем самым демонстрируя, что VVD MYC является полностью функциональным (21). Отныне все ссылки на уровни белка VVD основаны на экспрессии VVD MYC . Лизаты из штамма vvd , меченного myc, подвергнутого воздействию LL в течение 30 минут, инкубировали с антисывороткой FRH или FRQ перед зондированием иммунопреципитатов, подвергшихся блоттингу, антисывороткой против MYC для проверки наличия VVD.К сожалению, антисыворотка к FRQ оказалась слишком неспецифической в наших экспериментах с Co-IP, поэтому мы не смогли оценить, взаимодействует ли VVD с FRQ. Однако, когда мы использовали антисыворотку к FRH, VVD специфически иммунопреципитировал (фиг. 2 A ). Сигнал не был обнаружен в штаммах frq ko и vvd ko , в которых отсутствовала помеченная копия гена vvd , что указывает на то, что идентифицированный сигнал представляет собой VVD MYC , а не неспецифический сигнал.
Фиг.2.
VVD взаимодействует с FRH. ( A ) Антитело против FRH иммунопреципитирует VVD в анализе Co-IP. Белковые экстракты из штаммов vvd myc , vvd ko , wc-2 ko и frq ko , выращенных в течение 30 минут в LL, инкубировали с антителом FRH. Антисыворотка против FRH иммунопреципитирует VVD (нижняя полоса). Как и ожидалось, FRH иммунопреципитирует FRQ в штаммах WT и vvd ko , но не в контрольных штаммах frq ko и wc-2 ko .( B ) VVD может взаимодействовать с FRH в штамме (127-11), который лишен функциональных WCC и FFC ( SI Materials and Methods ), но содержит эктопическую qa-2-управляемую копию гена vvd [ qa-vvd myc (127-11)]. Вестерн-блоттинг общих (входных) экстрактов (две верхние дорожки), экстрактов, иммунопреципитированных антисывороткой к FRH (две средние дорожки) или неиммунизированной мышиной сыворотки (MS) (две нижние дорожки), зондировали антисывороткой к FRH или MYC для обнаружения FRH и VVD , соответственно.
Чтобы проверить, необходим ли функциональный WCC или FFC для взаимодействия VVD с FRH, мы использовали QA-индуцибельную систему, как описано выше, и экспрессировали qa-vvd myc в фоновом режиме, в котором wc- Были удалены гены 1 , wc-2 , frq и vvd (штамм 127–11) (рис. 2 B ). В этом штамме qa-vvd myc (127-11), в котором интактным остается только центральный ген часов frh , VVD все еще взаимодействует с FRH, что указывает на то, что для взаимодействия не нужны ни функциональные FFC, ни WCC.Взаимодействие происходит как на рассвете, так и на закате (рис. S2 B ). Как и ожидалось, экстракты показывают значительное истощение FRH после иммунного истощения (две верхние дорожки на фиг. S2 A ). Однако не наблюдается значительного истощения VVD, что позволяет предположить, что только небольшая часть общего VVD взаимодействует с FRH.
VVD влияет на транскрипционную конечность отрицательной обратной связи, опосредованной FFC.
Затем мы исследовали механизм, с помощью которого VVD опосредует оборот РНК frq .Описаны два различных пути, которые регулируют уровни сообщения frq . Во-первых, петля отрицательной обратной связи с участием FRQ и FRH важна для подавления ритма frq на уровне транскрипции (10, 11). Во-вторых, FFC также функционирует на посттранскрипционном уровне, контролируя деградацию мРНК frq через экзосомы (28).
Если VVD влияет на транскрипцию frq , ингибирование транскрипции должно устранить различия в уровнях frq , которые существуют между штаммами WT и vvd ko .Чтобы проверить эту возможность, мы использовали ингибитор транскрипции тиолютин (28) для ингибирования транскрипции за 1 час до или сразу после переноса культур Neurospora из светлого в темное (рис. 3). Когда транскрипция подавлялась перед переходом от света к темноте, мы не видели транскрипта frq или присутствовали очень мало в штаммах WT или vvd ko , что свидетельствует об эффективном подавлении транскрипции препаратом (вторая и четвертая дорожки в Рис.3 А ).Когда транскрипции frq позволяли перейти к границе света и темноты до добавления тиолютина, кинетика снижения транскрипта frq в штамме vvd ko больше не замедлялась и напоминала таковую у необработанного штамма WT ( сравните пятую полосу с верхней и третьей полосами на рис. 3 A ). Эти данные показывают, что VVD нацелен на транскрипцию frq .
Рис. 3.
VVD репрессирует транскрипцию frq . ( A ) Нозерн-блоты, показывающие уровни транскриптов frq в WT (две верхние дорожки) и vvd ko штаммов (третья, четвертая и пятая дорожки сверху) в культурах, выращенных в LL в течение 24 ч (момент времени 0) или для разных отрезков времени (ч) в DD.Культуры выращивали в присутствии (+) или отсутствии (-) ингибитора транскрипции тиолютина. Тиолютин добавляли либо за 1 ч до (-1 ч), либо сразу после (+0 ч) жидкие культуры переводили из светлого состояния в темное. Рибосомную РНК, окрашенную бромидом этидия, использовали для контроля загрузки Нозерн-блотов. ( B ) Количественный анализ Нозерн-блотов (показан в A ) необработанных WT (●), необработанных (▲) и обработанных тиолютином (сразу после переноса из светлого в темное) (△) vvd ко штаммов.В каждом эксперименте максимальные уровни frq РНК были установлены равными 100%.
Этот вывод был подтвержден экспериментом, в котором мы поместили frq под контроль QA-индуцибельного промотора. Конструкция qa- frq была интегрирована в штамм frq ko или frq ko vvd ko с двойным мутантным штаммом с образованием qa- frq ( frq17 ko ) и qa- frq ( frq ko vvd KO ) соответственно.По аналогии с нашей QA-индуцибельной системой экспрессии VVD MYC , описанной выше, этот эксперимент позволил нам отделить экспрессию frq от его нормальной индуцированной светом регуляции транскрипции и изучить снижение уровней транскрипта frq контролируемым образом после высвобождения. от индуктора. Если VVD нацелен на транскрипцию frq , замена промотора frq на промотор QA должна привести к аналогичному падению уровней транскрипта frq как в qa- frq ( frq ko ), так и в qa- . frq ( frq ko vvd KO ) штаммов после высвобождения из индуктора.С другой стороны, посттранскрипционное действие VVD на уровнях frq должно приводить к различию (т.е. задержке) оборота мРНК frq в qa- frq ( frq ko vvd KO ) по сравнению с qa- frq ( frq ko ). После высвобождения из индуктора наблюдали уровни frq в штаммах qa- frq ( frq ko ) и qa- frq ( frq ko vvd KO ) для штаммов периода a. 8 ч в DD, и мы не наблюдали значительной разницы в снижении кинетики транскрипта frq в двух штаммах.Существует некоторая вариабельность уровней frq , индуцированных QA, сразу после высвобождения из индуктора, но кинетика снижения frq аналогична для всех тестируемых штаммов и условий (фиг. S3 C ). Эти данные, таким образом, подтверждают вывод о том, что VVD влияет на frq РНК на уровне транскрипции, а не через опосредованную экзосомами функцию FFC. Интересно, что наличие или отсутствие света не оказало значительного влияния на оборот frq , что позволяет предположить, что свет не требуется для активации VVD (сравните рис.S3 A и B ).
VVD локализуется как в цитоплазме, так и в ядре.
Поскольку наши данные показали, что VVD репрессирует транскрипцию, мы исследовали локализацию VVD. Сообщается, что VVD локализуется в цитоплазме без каких-либо доказательств ядерной локализации (18). Однако использование антитела MYC для обнаружения меченного myc VVD снижает порог, при котором может быть обнаружен VVD (21). Мы приготовили ядерные и цитоплазматические экстракты из культур Neurospora, выращенных в DD и в разные моменты времени после переноса в LL (рис.4). В соответствии с предыдущими результатами, мы не обнаружили VVD в экстрактах ткани Neurospora, выращенной в периоды продолжительной темноты. Напротив, после 30 минут в LL значительные количества VVD были замечены как в полных экстрактах, так и в цитоплазматической фракции (фиг. 4 A ). При освещении слабый сигнал также обнаруживается в ядерных экстрактах, и этот сигнал становится сильнее с увеличением времени в LL. (Рис.4 A ). Мы не могли исключить возможность того, что увеличение ядерных ВВД является просто следствием резкого увеличения уровней ВВД во время воздействия света.Однако, когда они выражены как отношение ядерных уровней VVD к цитоплазматическим (Fig. 4 B ), наши данные подтверждают, что проникновение VVD в ядро может регулироваться светом.
Рис. 4.
VVD является цитоплазматическим и ядерным белком. ( A ) Тотальный (T), ядерный (N) или цитоплазматический (C) экстракты получали из ткани Neurospora, выращенной в течение 24 часов в DD (0) или с воздействием LL в течение указанного времени (h). Вестерн-блоты зондировали антителом MYC для обнаружения VVD MYC . Мембрана, окрашенная черным амидо, служит для контроля загрузки.( B ) График, показывающий процентное отношение ядерного сигнала к цитоплазматическому с использованием данных вестерн-блоттинга, представленных в A . ( C ) Экстракты Neurospora штаммов, экспрессирующих GFP-tagged VVD (под контролем промотора ccg-1 ), выращенные в течение 24 часов в DD или 4 часов в LL. Вестерн-блоты зондировали антителом к GFP для обнаружения VVD GFP . ЦП, цитоплазматический белок; NP, ядерный белок. ( D ) Субклеточная локализация VVD GFP (под контролем промотора ccg-1 ) в конидиоспорах Neurospora, фиксированных в моменты времени DD24 и LL4.Каждая субпанель показывает конфокальные изображения флуоресценции спор, окрашенных DAPI ( верхний левый ), флуоресценции сигнала GFP ( верхний правый ), наложение обоих ( нижний правый ) и соответствующее изображение в светлом поле (). Нижний левый ). (Масштабная шкала, 1 мкм.)
Для сбора независимой информации о локализации VVD в клетке мы создали карбоксиконцевые слияния GFP и VVD с использованием плазмид (30), в которых экспрессия генов контролируется сильным Neurospora ccg-1 промоутер.Созданная ccg-1 конструкция vvd-gfp способна спасать фенотипы каротиноидов и часов штамма vvd ko (рис. S4 A и C ) и уровни 17 VVD 917 GFP несколько выше, чем в штамме, который экспрессирует vvd под собственным промотором (фиг. S4 B ). Фракционирование клеток штаммов VVD, меченных GFP, подтвердило результаты, полученные ранее для VVD, меченных myc (рис. 4 A ), о том, что VVD локализуется как в цитоплазме, так и в ядре (рис.4 С ). Для конфокальной микроскопии мы собирали конидии (которые обычно содержат от одного до трех ядер) из штаммов, хранимых в DD в течение 24 часов, и из штаммов, хранящихся в LL в течение 4 часов. В небольшом количестве конидий можно четко увидеть ядерную локализацию VVD GFP (рис. 4 D ), что еще раз подтверждает результаты, полученные с помощью наших клеточных экстрактов. Как и ожидалось, конидии, полученные из слитого штамма 1 H-GFP, также обнаруживают ядерную локализацию для 1 H GFP (рис.S4 D , Низ ). Напротив, мы никогда не видели ядерный сигнал выше фона в ядрах от штаммов WT, которые не были помечены GFP (рис. S4 D , Top и Middle ). Поскольку опубликованные данные предполагают, что GFP сам по себе является цитоплазматическим и не локализуется в ядре (31), низкий процент ядер, которые флуоресцируют в штаммах с меткой vvd gfp , предполагает, что накопление ядер или проникновение в VVD может быть очень динамичным. В заключение, наши объединенные данные подтвердили, что значительная часть VVD локализуется в ядре.
VVD Взаимодействует с WCC.
Ядерная локализация VVD и взаимодействия VVD с FRH позволила нам проверить, репрессирует ли VVD транскрипцию frq путем связывания с WCC. Такая активность согласуется с наблюдением, что VVD взаимодействует с FRH, потому что FRH опосредует фосфорилирование и инактивацию WCC (15, 32, 33). Более того, было показано, что VVD влияет на фосфорилирование WC-1 и, следовательно, на активность WCC (16, 23). Мы провели эксперименты Co-IP, в которых лизаты цельных клеток Neurospora инкубировали с антисывороткой WC-1 перед зондированием на наличие VVD.Как видно на фиг. 5, VVD иммунопреципитировался как в LL (фиг. 5 A, и B ), так и в DD (фиг. 5 D ).
Рис. 5.
VVD взаимодействует с WCC на свету и в темноте. ( A ) Белковые экстракты штаммов vvd myc , vvd ko и wc-1 ko инкубировали с антителом WC-1. WC-1 иммунопреципитирует VVD в штамме vvd myc , но не в штамме vvd ko или wc-1 ko .( B ) Антитело WC-1 не реагирует перекрестно с VVD. QA-индуцибельный VVD экспрессировался на фоне vvd ko и vvd ko , wc-1 ko и белковых экстрактов, коиммунопреципитированных с антителом WC-1. VVD MYC не был иммунопреципитирован на фоне vvd ko , wc-1 ko . ( C ) Контроль, демонстрирующий, что QA-индуцибельный VVD MYC стабильно экспрессируется в отсутствие WC-1.( D ) Антисыворотка к WC-1 иммунопреципитирует QA-индуцированный VVD MYC в DD в штамме wc-1 + ( qa-vvd myc ). Антисыворотка к WC-1 не может иммунопреципитировать VVD MYC в штамме (127-11), в котором отсутствует WC-1, но экспрессируется QA-индуцированный VVD MYC на уровнях, аналогичных контрольному штамму qa-vvd myc . ( E ) Небольшая часть VVD обнаруживается во фракциях градиента сахарозы, содержащих WC-1, WC-2 и FRH.Вестерн-блоттинг (верхние пять дорожек) и график (, нижний ), показывающий денситометрический анализ VVD MYC , WC-1, WC-2 и FRH из белковых экстрактов, фракционированных по размеру в градиенте сахарозы 10–35%. Фракция 1 соответствует низкой молекулярной массе, а фракция 16 — более высокой молекулярной массе. Звездочки обозначают неспецифические сигналы. Для денситометрии максимальный сигнал для каждого белка был установлен равным 100%.
Поскольку домены PAS / LOV VVD и WC-1 схожи, важно было контролировать специфичность взаимодействия WC-1 – VVD.Поэтому мы создали штаммы, в которых vvd myc могло быть экспрессировано на фоне wc-1 ko . Поскольку экспрессия vvd зависит от WC-1, QA-управляемый ген vvd был экспрессирован, чтобы отделить экспрессию VVD от экспрессии WC-1. Конструкция qa- vvd была интегрирована в vvd ko или vvd ko wc-1 ko с двойным мутантом для создания qa- vvd myc 917 v1528 ko ) и qa- vvd myc ( vvd ko wc-1 KO ) соответственно.В этих штаммах VVD MYC экспрессируется на сопоставимых уровнях, хотя уровни VVD оказались немного ниже у штамма qa- vvd myc ( vvd ko wc-1 KO ) (рис. С ). В этих экспериментах VVD был иммунопреципитирован только в штаммах, которые экспрессируют белок WC-1 (фиг. 5 B ). В другом контрольном эксперименте с использованием штаммов qa-vvd myc (WT) и qa-vvd myc (127-11) мы наблюдали, что VVD иммунопреципитируется только в qa-vvd myc ( WT) (рис.5 D ). Если VVD взаимодействует с WC-1 через WCC, мы ожидали, что VVD также будет взаимодействовать с WC-2. Однако нашим усилиям по тестированию этого взаимодействия с помощью Co-IP препятствовали неспецифические перекрестные взаимодействия антисывороток WC-2 и MYC с VVD , MYC и WC-2, соответственно.
Чтобы проверить, должен ли VVD быть активным в передаче сигналов для взаимодействия с WC-1, мы протестировали два мутантных штамма, у которых отсутствует светоиндуцированная активность: vvd C71S , который является биологически неактивным из-за его неспособности к гомодимеризации (26). , и vvd C108A , штамм, у которого нарушено образование цистеинилового аддукта, который имеет решающее значение для передачи светового сигнала (18) (рис.S2 C ). Оба мутантных белка взаимодействовали с WC-1, предполагая, что VVD не обязательно должен находиться в активном сигнальном состоянии для взаимодействия с WC-1.
Чтобы дополнительно исследовать, является ли VVD частью комплексов с более высокой молекулярной массой, как предполагают наши эксперименты по иммунопреципитации, мы провели эксперименты с градиентом сахарозы (рис. 5 E ). В соответствии с предыдущими результатами, мы обнаружили WC-2 в двух пиках, которые, как сообщается, имели размер около 60 кДа и 200 кДа, и обнаружили, что WC-1 мигрирует одновременно с пиком с более высокой молекулярной массой, идентифицированным для WC-2 (33).FRH мигрировал в виде единственного пика, который перекрывает как WC-1, так и WC-2. Широкий пик VVD MYC , охватывающий фракции, в которых мигрирует мономерный WC-2, согласуется с VVD, образующим гомодимер, как было недавно предположено (24–26). Присутствие VVD во фракциях, которые содержат гораздо более высокомолекулярные комплексы WC-1, WC-2 и FRH, согласуется с нашими данными Co-IP, которые показали взаимодействие VVD с WC-1 и FRH.
Взаимодействует ли VVD с ДНК-связанными WCC? Мы проверили эту возможность в EMSA (рис.S5). Ядерные белки (LL 4 h) экстрагировали из штаммов vvd ko , vvd myc или vvd gfp и инкубировали с ранее идентифицированным проксимальным светочувствительным элементом (pLRE), расположенным в области frq. промотор (12). В соответствии с опубликованными результатами (12, 34, 35), свободный зонд был захвачен высокомолекулярным комплексом при инкубации с ядерными экстрактами, а комплекс был сверхсмещен при инкубации с антисывороткой WC-2 (рис.S5). Однако мы не наблюдали сверхсдвига, когда экстракты инкубировали либо с антителом MYC (фиг. S5 A ), либо с антителом GFP (фиг. S5 B ), что позволяет предположить, что VVD связывает свободный, а не связанный с ДНК WC-1.
Обсуждение
Мы показали, что VVD взаимодействует с центральными компонентами циркадных часов Neurospora и с компонентами синего света, подавляя транскрипционную активность WCC на рассвете и в сумерках. Модель, показанная на рис. 6, показывает, как VVD может выполнять свою функцию при переходе от рассвета к закату.Мы знаем, что WC-1 и WC-2 образуют мультимеры, которые могут связываться с проксимальным и дистальным цис- элементами GATN в промоторе frq (12). Комплекс, образованный в темноте (WCC D ), мигрирует быстрее (т.е. меньше), чем комплекс, образованный на свету (WCC L ). Последний опосредует транскрипцию светоиндуцированных генов, таких как vvd (12, 35). Световая индукция frq регулируется главным образом через проксимальный светочувствительный элемент (LRE), тогда как дистальный элемент, хотя и светочувствительный, необходим для устойчивой ритмичности frq в DD (12).Мы показали здесь, что VVD связывает WC-1 как в LL, так и после высвобождения в DD. Таким образом, кажется вероятным, что VVD нарушает эффективное образование WCC L . Это нарушение д. Приводить к снижению индуцированной светом транскрипции в LL, а также к эффективному нарушению DD WCC L , который присутствует при переходе от света к темноте. В отсутствие VVD равновесие между WCC D и WCC L будет смещено в сторону WCC L , что приведет к увеличению транскрипции frq на свету, а также в первые часы в темноте.Однако, поскольку frq регулирует свою собственную транскрипцию посредством своего собственного генного продукта, подавление frq в DD является не просто функцией уровней и активности WCC, но зависит от уровня и активности FRQ.
Рис. 6.
Модель действия ВВД при регулировании ВКК с отрицательной обратной связью. ( A ) В темноте WC-1 и WC-2 гетеродимеризуются с образованием WCC D . Под воздействием света образуется мультимерный комплекс WCC, WCC L (11, 14), который опосредует транскрипцию индуцированных светом генов (например,г., ввд ). VVD связывает WC-1, чтобы ингибировать эффективное образование WCC L , что приводит к снижению индуцированной светом транскрипции. Без VVD равновесие между светлыми и темными комплексами смещается в сторону WCC L , что приводит к увеличению транскрипции frq на свету. При переносе на DD и без VVD повышенная активность WCC L приводит к пролонгированной активации транскрипции frq (возможно, через его проксимальный LRE), тогда как в WT FRQ-опосредованное фосфорилирование WCC D приводит к быстрой инактивации. транскрипции frq .WCC L и WCC D имеют разные предпочтения в отношении проксимальных и дистальных промоторных элементов, причем WCC D предпочтительно задействован в дистальном часовом ящике, где имеет место регуляция отрицательной обратной связи посредством FRQ (11). ( B ) Упрощенная схема профиля РНК frq после световой индукции и в постоянной темноте в WT (жирная линия) и в штамме vvd KO (тонкая линия). Серая область отображает разницу в активации frq между двумя штаммами и является результатом изменения равновесия между WCC D и WCC L .Длительная активация в темноте приводит к характерной фазовой задержке начала колебаний транскрипта frq и явной циркадной ритмичности.
Поскольку известно, что ( i ) FRQ не может эффективно репрессировать собственную транскрипцию на свету (36), ( ii ), световая индукция frq в значительной степени контролируется проксимальным LRE (12), и ( iii ) WCC L свидетельствует об активированном светом состоянии WCC (12, 35), кажется вероятным, что FRQ не может эффективно репрессировать собственную транскрипцию через WCC L , но проявляет свою основную репрессивную активность в отношении WCC. Д .Эта возможность может предоставить альтернативное объяснение того, почему frq требует больше времени для снижения в штамме vvd ko , потому что большая часть WCC, как ожидается, будет в форме WCC L .
Наши эксперименты показывают, что лишь небольшое количество ВВД находится в комплексе с WC-1. Поскольку влияние VVD на световую перезагрузку циркадных часов и фотоадаптацию достаточно велико, можно ожидать, что большая часть VVD должна образовывать комплекс с FFC и WCC для выполнения своей функции в этих процессах.Следовательно, мы поддерживаем идею о том, что взаимодействие непродолжительное, но не обязательно слабое. Эта ситуация напоминает наблюдение, что только очень небольшие количества FRQ, по-видимому, взаимодействуют с WCC (15). Точно так же наблюдение, что FRQ, по-видимому, не является частью связанного с промотором WCC (12), напоминает наше наблюдение, что антисыворотки против VVD MYC или VVD GFP не вызывают суперсдвига WCC. Однако возможно, что эти антитела менее эффективны в экспериментах с EMSA или что ДНК-связанное взаимодействие слишком слабое или временное, чтобы его можно было обнаружить с помощью EMSA; следовательно, мы не можем исключить ДНК-связанное взаимодействие на этой стадии.Интересно, что мутанты VVD, которые считаются дефектными в отношении фотосигнализации, все еще могут взаимодействовать с WC-1 и FRH, предполагая, что репрессивные функции VVD в передаче световых сигналов могут происходить как часть комплекса, со световой активацией VVD, влияющей на конформацию весь комплекс. Помимо роли VVD в подавлении световых ответов, есть доказательства того, что VVD способствует некоторым аспектам передачи световых сигналов (18), а более недавнее исследование на микроматрицах предполагает, что VVD играет роль в модуляции поздних световых ответов у Neurospora (22).Следовательно, вероятно, что VVD может иметь функции, которые не требуют его взаимодействия с WCC или FFC.
Наконец, наши данные показывают, что VVD может взаимодействовать с FRH в отсутствие функционального FFC или WCC, и есть соблазн предположить, что FRH является первичной платформой, на которой собираются различные комплексы.
Материалы и методы
Плазмиды и штаммы.
Штамм 54–3 ( bd , a ) использовали в качестве WT во всех экспериментах. Подробное описание того, как были созданы штаммы, можно найти в SI Materials and Methods .Все штаммы проверяли с помощью ПЦР или Саузерн-блоттинга. Гомокарионы были получены, как описано ранее (37).
Условия роста.
Эксперименты с гоночной пробиркой и жидкими культурами проводили в камерах Sanyo MLR-350 с регулируемым светом и температурой, как описано в SI Materials and Methods . Для экспериментов с ингибиторами транскрипции добавляли тиолютин (Tocris Biosciences) до конечной концентрации 12 мкг / мл. Контрольные образцы обрабатывали ДМСО.
Фракционирование клеток и EMSA.
Клеточные фракции выделяли по существу, как описано ранее (38), подробности приведены в SI Materials and Methods . Для EMSA последовательности проксимальных LRE-олигонуклеотидов frq (Eurofins MWG Operon), описанные ранее (12), использовали для создания зонда дцДНК. Условия связывания и гель-электрофорез описаны в SI Materials and Methods .
Градиенты сахарозы.
Белковые экстракты получали с использованием стандартного буфера для экстракции белков, как описано ранее (30), и эксперименты с градиентом сахарозы проводили, как подробно описано в S1 Материалы и методы .
Экстракция РНК и Нозерн-блоттинг.
РНК экстрагировали с помощью мини-набора Qiagen RNeasy в соответствии с инструкциями производителя. Нозерн-блоттинг выполняли, как описано ранее (20) и как подробно описано в S1 Materials and Methods .
Конфокальная микроскопия.
Конидии фиксировали в 4% формальдегиде в течение 1,5 ч, промывали дистиллированным H 2 O, инкубировали в 50 мкг / мл DAPI (Sigma) в течение 10 мин при комнатной температуре, а затем промывали и ресуспендировали в 25% глицерине.Подробная информация о сборе изображений приведена в S1 Материалы и методы .
Благодарности
Мы благодарим Сью Кростуэйт (Манчестерский университет, Манчестер, Великобритания) за полезные предложения и критическое прочтение рукописи, а также Джея Данлэпа, Дженнифер Лорос (Дартмутская медицинская школа, Ганновер, штат Нью-Хэмпшир) и Йи Лю (Техасский университет). Southwestern Medical Center, Даллас, Техас) за щедрый подарок FRQ WC-1, WC-2 и антисыворотки FRH. Мы особенно благодарим Джейн Котт за помощь с микроскопией.Микроскопы, использованные в этом исследовании, были приобретены на гранты Исследовательского совета биотехнологии и биологических наук, Wellcome Trust и Стратегического фонда Манчестерского университета. Эта работа была поддержана грантами BB / D00988X / 1 и BB / F012055 / 1 от Исследовательского совета биотехнологии и биологических наук, предоставленным C.H.
Сноски
- 1 Кому следует направлять корреспонденцию. Эл. Почта: christian.heintzen {at} manchester.ac.uk.
Автор: С.M.H. и Ч. спланированное исследование; S.M.H., S.T. и M.E. проводили исследования; S.M.H., S.T., M.E. и C.H. проанализированные данные; и Ч. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
↵ * Для этой статьи с прямым представлением был назначен редактор.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1009474107/-/DCSupplemental.
CKI и CKII опосредуют ЧАСТОТНО-зависимое фосфорилирование комплекса WHITE COLLAR, чтобы закрыть петлю циркадной отрицательной обратной связи Neurospora
- Qun He1,2,
- Джунсок Ча1,
- Qiyang He1,3,
- Хэн-Чи Ли1,
- Yuhong Yang1,4, и
- Йи Лю1,5
- 1 Отделение физиологии Юго-Западного медицинского центра Техасского университета, Даллас, Техас 75390, США;
- 2 Государственная ключевая лаборатория агробиотехнологии, Колледж биологических наук, Китайский сельскохозяйственный университет, Пекин 100094,
Китай
Аннотация
Циркадные осцилляторы эукариот состоят из циркадных петель отрицательной обратной связи.В Neurospora было высказано предположение, что белок FREQUENCY (FRQ) способствует фосфорилированию комплекса WHITE COLLAR (WC), тем самым ингибируя
его деятельность. Киназы, участвующие в этом процессе, неизвестны. В этом исследовании мы показываем, что нарушение взаимодействия
между FRQ и CK-1a (гомолог казеинкиназы I) приводит к гипофосфорилированию FRQ, WC-1 и WC-2. В штамме ck-1a L нокаут-мутант, несущий мутацию, эквивалентную мутации Drosophila dbt L , FRQ, WC-1 и WC-2, являются гипофосфорилированными.У мутанта также наблюдаются ~ 32-часовые циркадные ритмы из-за увеличения
стабильности FRQ и значительной задержке прогрессирующего фосфорилирования FRQ. Кроме того, уровни WC-1 и WC-2 являются
низкий у штамма ck-1a L , что указывает на то, что CK-1a также важен для циркадных петель положительной обратной связи. Несмотря на низкую аккумуляцию
в штамме ck-1a L гипофосфорилированный WCC эффективно связывается с C-боксом внутри промотора frq , предположительно потому, что он не может быть инактивирован посредством FRQ-опосредованного фосфорилирования.Кроме того, WC-1 и WC-2
также гипофосфорилирован в штамме cka RIP , который несет нарушение каталитической субъединицы казеинкиназы II. В штамме cka RIP связывание WCC с C-боксом постоянно высоко и не может быть ингибировано FRQ, несмотря на высокие уровни FRQ, что приводит к
высокие уровни frq РНК. Вместе эти результаты предполагают, что CKI и CKII, помимо того, что являются киназами FRQ, опосредуют FRQ-зависимое фосфорилирование.
WC, которые подавляют их активность и замыкают циркадную отрицательную обратную связь.
Циркадные часы эукариот состоят из саморегулирующих циркадных петель отрицательной обратной связи, включая как положительные, так и положительные.
и отрицательные элементы (Dunlap 1999; Reppert and Weaver 2001; Young and Kay 2001; Sehgal 2004). У Neurospora , Drosophila и млекопитающих все положительные элементы представляют собой гетеродимерные комплексы, состоящие из двух PER-ARNT-SIM (PAS) доменов, содержащих
транскрипционные факторы, которые связываются с цис- -элементами в промоторе отрицательных элементов, чтобы активировать их транскрипцию.С другой стороны, отрицательные элементы
подавляют собственную транскрипцию, подавляя активность положительных элементов через их физические взаимодействия. Это
Неясно, как отрицательные элементы подавляют активность положительных элементов, чтобы закрыть циркадные петли отрицательной обратной связи.
После идентификации гена Drosophila doubletime ( dbt ), который кодирует гомолог казеинкиназы I (CKI), стало ясно, что посттрансляционное фосфорилирование белка
необходим для работы циркадных часов (Kloss et al.1998; Price et al. 1998). Несмотря на эволюционное расстояние, среди различных эукариотических организмов существует замечательная консервация посттрансляционной регуляции.
системы от грибов до человека (см. обсуждение; Liu 2005; Heintzen and Liu 2006).
У нитчатого гриба Neurospora crassa основная циркадная отрицательная обратная связь состоит из четырех основных компонентов: БЕЛЫЙ ОШЕЙНИК-1 (WC-1), WC-2, ЧАСТОТА
(FRQ) и FRQ-взаимодействующая РНК-геклиаза FRH.WC-1 и WC-2, два фактора транскрипции, содержащие домен PAS, образуют гетеродимерный
Комплекс WC (D-WCC) в темноте, который связывается с Clock (C) -box в промоторе frq , чтобы активировать транскрипцию frq (Crosthwaite et al. 1997; Cheng et al. 2001b, 2002; Loros and Dunlap 2001; Froehlich et al.2003; He and Liu 2005). Таким образом, WCC является положительным элементом в петле циркадной отрицательной обратной связи Neurospora . С другой стороны, FFC, комплекс, образованный FRQ и FRH, функционирует как отрицательный элемент.
(Cheng et al.2005). Чтобы подавить транскрипцию frq , FFC, возможно, опосредует ингибирование активности WCC посредством их физического взаимодействия (Aronson et al. 1994; Merrow et al. 1997, 2001; Cheng et al. 2001a, 2003; Denault et al. 2001). ; Froehlich et al.2003). У штамма дикого типа эта циркадная отрицательная обратная связь генерирует устойчивые суточные ритмы frq РНК и белка FRQ (Garceau et al. 1997). Когда циркадная отрицательная обратная связь нарушается мутацией frq или понижающей регуляцией frh , уровни РНК frq остаются на постоянном более высоком уровне, что приводит к аритмии (Aronson et al.1994; Cheng et al. 2005). Помимо своей роли в циркадной петле отрицательной обратной связи в темноте, WC-1 и WC-2 также являются важными компонентами.
для световых откликов и световой переустановки часов, причем WC-1 является фоторецептором синего света (Crosthwaite et al. 1997; Froehlich et al. 2002; He et al. 2002; He and Liu 2005).
Помимо подавления активности D-WCC в темноте, FRQ способствует накоплению WC-1 и WC-2, образуя положительный
петли обратной связи, которые связаны с отрицательной петлей (Lee et al.2000; Cheng et al. 2001b, 2003; Merrow et al. 2001), что характерно для циркадных систем животных (Glossop et al. 1999; Shearman et al. 2000). В Neurospora эти петли положительной обратной связи, как было показано, важны для надежности и функционирования часов (Cheng et al. 2001b; Schafmeier et al. 2006). Недавно было показано, что фосфорилирование области PEST-2 цитоплазматического FRQ важно для накопления
WC-1, но не WC-2 (Schafmeier et al.2006 г.).
FRQ, WC-1 и WC-2 регулируются событиями фосфорилирования. После синтеза FRQ сразу фосфорилируется и становится
прогрессивно более фосфорилируется с течением времени до его деградации через убиквитин-протеасомный путь, опосредованный FWD-1
(Гарсо и др., 1997; Он и др., 2003, 2005a). Таким образом, в темноте FRQ не только сильно ритмичен по количеству, но и по его состояниям фосфорилирования. СК-1а (казеин
киназа 1a), CKII (казеинкиназа II) и CAMK-1 являются тремя идентифицированными киназами FRQ (Gorl et al.2001; Ян и др. 2001, 2002, 2003). Однако только роль CKII в обеспечении фосфорилирования FRQ твердо установлена in vivo (Yang et al. 2002, 2003). In vitro CKII является одной из основных киназ, фосфорилирующих FRQ. У штаммов, в которых либо каталитическая субъединица CKII
( cka ) или одна из его регуляторных субъединиц ( ckb1 ) нарушена, FRQ гипофосфорилирован и более стабилен, а функция часов либо полностью отменена (мутант cka ), либо колеблется с сильно затухающей амплитудой ( ckb1 мутант).Более того, в мутантном штамме cka , который не имеет активности CKII, уровни мРНК frq постоянно высоки, что напоминает уровни frq РНК в штаммах с нарушенной циркадной петлей отрицательной обратной связи (Yang et al., 2002) . Эти данные предполагают, что CKII не только способствует деградации FRQ, но также необходим для репрессорной функции FRQ.
Механизм, с помощью которого CKII выполняет эту последнюю функцию, неизвестен.
CK-1a является одним из двух гомологов Neurospora CKI и может фосфорилировать PEST-1 и PEST-2 домены FRQ in vitro (Gorl et al.2001). Удаление домена PEST-1 привело к повышению стабильности FRQ и долгопериодному ритму. Важнее,
CK-1a, как было обнаружено, ассоциирован с FRQ, предполагая, что он может фосфорилировать FRQ in vivo (Gorl et al. 2001; Cheng et al. 2005). Однако in vivo доказательств участия CK-1a в работе часов не было, потому что CK-1a важен для клеток.
выживаемость в Neurospora .
Подобно FRQ, как WC-1, так и WC-2 фосфорилируются как в темноте, так и в зависимости от света.Их фосфорилирование
играет важную роль в регуляции активности WCC (He and Liu 2005; He et al. 2005b; Schafmeier et al. 2005, 2006). Ранее мы идентифицировали пять основных сайтов фосфорилирования WC-1 in vivo, расположенных непосредственно ниже его ДНК-связывания.
домен (He et al. 2005b). Мутация этих независимых от света участков предполагает, что они имеют решающее значение для функции циркадных часов и отрицательно сказываются на них.
регулируют деятельность D-WCC. Важность фосфорилирования WC в циркадных часах позже была подтверждена неожиданным
наблюдение Schafmeier et al.(2005), что фосфорилирование WC зависит от FRQ. В нулевом штамме frq как WC-1, так и WC-2 гипофосфорилированы. В штамме дикого типа WC-2 демонстрирует устойчивый циркадный ритм
его профиль фосфорилирования при анализе с помощью двумерного электрофореза. Важно отметить, что активация транскрипции frq коррелирует с гипофосфорилированием WCs. В соответствии с этими данными мы показали, что дефосфорилирование
из Neurospora WCC значительно способствует его связыванию с C-боксом (He and Liu 2005).Вместе эти результаты предлагают модель, в которой FFC ингибирует активность WCC, способствуя фосфорилированию белков WC.
Интересно, что PER-зависимое фосфорилирование CLK недавно также наблюдалось у Drosophila (Kim and Edery 2006; Yu et al. 2006), подтверждая общий механизм, который замыкает циркадные петли отрицательной обратной связи.
Киназа (ы), привлекаемая FRQ для фосфорилирования белков WC, не идентифицирована.Как на деятельность WCC влияет
фосфорилирование, опосредованное этой киназой (ами), также неизвестно. В этом исследовании мы показываем, что, как и CKII, CK-1a фосфорилирует
FRQ in vivo способствует его деградации. Что еще более важно, обе киназы опосредуют FRQ-зависимое фосфорилирование WCC, которое
подавляет свою активность, чтобы замкнуть циркадную отрицательную обратную связь. В дополнение к роли CK-1a в отрицательной обратной связи
петля, она также необходима для функционирования циркадных петель положительной обратной связи за счет увеличения уровней WC.
Результаты
Нарушение взаимодействия FRQ / CK-1a приводит к гипофосфорилированию FRQ
Чтобы понять функцию CK-1a in vivo в часах Neurospora , мы исследовали функциональную важность взаимодействия FRQ / CK-1a. Сначала мы сопоставили домен с FRQ, то есть
требуется для взаимодействия FRQ / CK-1a.На рисунке 1А показана доменная структура FRQ. Ранее было показано, что домены coiled-coil (CC) и FFD опосредуют FRQ – FRQ или FRQ – FRH.
взаимодействия, соответственно (Cheng et al. 2001a, 2005). S513 является одним из ранее идентифицированных сайтов фосфорилирования FRQ, который опосредует деградацию FRQ (Liu et al. 2000). PEST-1 и PEST-2 представляют собой два домена PEST, которые могут фосфорилироваться с помощью CK-1a in vitro (Gorl et al. 2001; Schafmeier et al. 2006). Для изучения взаимодействия FRQ / CK-1a мы ввели конструкцию (pMyc-CK-1a), экспрессирующую Myc-tagged CK-1a (под хинной кислотой
индуцибельный промотор) в штамм дикого типа и серию небольших мутантных штаммов FRQ (sFRQ) (на фоне frq 10 [ frq -null]), которые экспрессируют различные внутренние делеции в рамке считывания формы sFRQ покрывающая всю ORF sFRQ (рис.1А; Лю и др. 2000; Cheng et al. 2005). Вестерн-блоттинг показал экспрессию Myc-CK-1a в виде триплетных полос (фиг. 1B), возможно, либо из-за альтернативной инициации белка, либо из-за посттрансляционной модификации CK-1a. Иммунопреципитация
с использованием моноклонального антитела c-Myc показало, что Myc-CK-1a специфически взаимодействует с FRQ дикого типа и всеми формами sFRQ, кроме
для sFRQ4 (рис. 1A, B; дополнительный рис. 1). Эти данные указывают на то, что требуется область белка, удаленная в sFRQ4, но не в других частях FRQ.
для взаимодействия FRQ / CK-1a.
Фигура 1.
Нарушение FRQ / CK-1a приводит к гипофосфорилированию FRQ. ( A ) Графические диаграммы, показывающие доменную структуру FRQ и различных мутантов с внутренней делецией sFRQ. (CC) Спиральный домен;
(FFD) домен взаимодействия FRQ – FRH; (S513) ранее идентифицированный сайт фосфорилирования FRQ.Аминокислоты FRQ удалены в каждой
указаны мутанты. ( B ) Анализ иммунопреципитации, показывающий, что sFRQ4 не может взаимодействовать с Myc-CK-1a. Звездочка указывает на сигнал IgG. ( C ) Графические диаграммы, показывающие домен взаимодействия FRQ / CK-1a (FCD) и внутренние делеции FRQ у различных мутантов. Черный
столбцы указывают на области, удаленные в различных мутантах. Указаны аминокислоты FRQ, удаленные в каждом мутанте.Кроме
FRQ4A1, который экспрессирует полноразмерный белок FRQ с указанной делецией, другие мутанты экспрессируют только sFRQ. Аминокислота
выравнивание доменов FCD различных гомологов FRQ показано в позиции , внизу , а идентичные остатки среди разных гомологов заключены в рамку. (Neu) Neurospora crassra ; (Mag) Magnaporthe grisea ; (Сор) Sordaria fimicola ; (Lep) Leptosphaeria australiensis ; (Gib) Gibberella zeae ; (Hyp) Hypocrea spinulosa .В мутантах 4A11 и 4A12 (выражающих полноразмерный FRQ) аминокислоты, указанные звездочками, были мутированы на аланины.
(p) Три ранее идентифицированных сайта фосфорилирования. ( D ) Анализ иммунопреципитации с использованием антитела c-Myc, показывающий, что FRQ не может взаимодействовать с Myc-CK-1a в FRQ4A11 и FRQ4A12
мутанты. ( E ) Вестерн-блот-анализ, показывающий, что FRQ гипофосфорилируется в мутантах FRQ4A11 и FRQ4A12.Звездочка указывает
неспецифическая полоса, распознаваемая нашим антителом FRQ. Все культуры выращивали в LL.
Интересно, что FRQ в sFRQ4, как было ранее показано, является гипофосфорилированным и более стабильным (Liu et al. 2000; Gorl et al. 2001). Область FRQ, удаленная в sFRQ4, содержит три идентифицированных сайта фосфорилирования FRQ (T501, S513 и S519) и
PEST-1 домен (рис.1С). Дальнейшие делеционные анализы показали, что требуется область из 63 аминокислот (аминокислоты 470–532) перед PEST-1.
для взаимодействия FRQ / CK-1a (рис. 1C; дополнительный рис. 1). Выравнивание аминокислотной последовательности этой области, названной FCD (для домена взаимодействия FRQ / CK-1a), среди
различных гомологов FRQ грибов, показывает, что это одна из наиболее консервативных областей в ORF FRQ, что предполагает ее важность
в функции FRQ.
FCD не содержит мотива F-X-X-X-F, который ранее был идентифицирован как сайт стыковки CK1 в белках mPER и NFAT млекопитающих.
(Окамура и др.2004 г.). Это указывает на то, что существуют разные сайты стыковки, опосредующие взаимодействие CKI с FRQ и mPERs. Три известных FRQ
Сайты фосфорилирования находятся в С-концевой части FCD, которая находится непосредственно перед доменом PEST-1. Это предполагает
что CK-1a может связываться с N-концевой частью этого домена и фосфорилировать FRQ по этим сайтам. Чтобы проверить эту гипотезу, мы
мутировал два участка высококонсервативных последовательностей непосредственно перед T501, аминокислотами 488–491 (LLCN) и аминокислотами
493–496 (QLH) в аланины в полноразмерной конструкции frq соответственно (рис.1С). Анализ иммунопреципитации показал, что эти точечные мутации полностью устраняют взаимодействие FRQ / CK-1a (фиг. 1D), указывая на то, что они являются критическими остатками, опосредующими взаимодействие между FRQ и CK-1a.
Чтобы исследовать важность взаимодействия FRQ / CK-1a в фосфорилировании FRQ, паттерны фосфорилирования FRQ в FRQ4A11
и штаммы FRQ4A12 сравнивали со штаммами дикого типа.Культуры выращивали при постоянном освещении (LL), в условиях
это приводит к равномерному распределению различных фосфорилированных форм FRQ (Garceau et al. 1997; Liu et al. 2000). Как показано на Фигуре 1E, FRQ в этих двух мутантных штаммах был гипофосфорилирован и не имел сильно фосфорилированных форм FRQ, присутствующих в
штамм дикого типа. Эти данные вместе с предыдущими результатами предполагают, что CK-1a связывает и фосфорилирует FRQ in vivo. Несколько
Фосфорилированные формы FRQ все еще можно было наблюдать в мутантных штаммах, что позволяет предположить, что они могут фосфорилироваться другими
киназы, включая CKII (Yang et al.2001, 2002, 2003).
Взаимодействие FRQ / CK-1a необходимо для FRQ-зависимого фосфорилирования WC и важно для функции циркадных часов
Взаимодействие FRQ с WCC и CK-1a предполагает, что WCC может связываться с CK-1a через FRQ и что CK-1a может опосредовать
FRQ-зависимое фосфорилирование WC. Чтобы проверить эту гипотезу, мы исследовали, могут ли белки WC и CK-1a образовывать комплекс
в зависимости от FRQ.Как показано на Фигуре 2A, иммунопреципитация Myc-CK-1a разрушила WC-1 в штамме дикого типа, но не в штамме frq 10 , что указывает на то, что WC-1 и CK-1a могут образовывать комплекс друг с другом. и что это опосредовано FRQ.
Фигура 2.
Нарушение FRQ / CK-1a приводит к гипофосфорилированию FRQ и аритмическим ритмам конидиации.( A ) Анализ иммунопреципитации с использованием антитела Myc, показывающий, что WC-1 образует комплекс с Myc-CK-1a в штамме дикого типа, но
не в штамме frq 10 . ( B , C ) Вестерн-блоттинг показывает, что WC-1 и WC-2 гипофосфорилированы и находятся на низких уровнях в штаммах frq 10 , sFRQ4, FRQ4A11 и FRQ4A12. Стрелки указывают на сильно фосфорилированные виды WC, отсутствующие у мутантов.( D ) Анализ иммунопреципитации с использованием антитела WC-2, показывающий, что мутантный FRQ в штаммах FRQ4A11 и FRQ4A12 все еще может взаимодействовать
с WC комплексом. Все культуры выращивали в LL. (PI) Иммунопреципитация с использованием преиммунной сыворотки. ( E ) Анализы с гоночной трубкой, показывающие аритмическую конидиацию в штаммах FRQ4A11 и FRQ4A12. KAJ120 представляет собой штамм frq 10 , содержащий конструкцию frq дикого типа.Черные линии указывают на ежедневные фронты роста культур.
Если CK-1a фосфорилирует белки WC посредством взаимодействия с FRQ, мы ожидали, что нарушение взаимодействия FRQ / CK-1a
должно привести к потере FRQ-зависимого фосфорилирования WC. Мы обнаружили, что при использовании гелей SDS-PAGE, содержащих соотношение 149: 1
акриламид / бисакриламид привел к значительно лучшему разделению фосфорилированных форм WC, чем обычно используемые
37.5: 1 SDS-PAGE гели. Это особенно верно для более мелкого белка WC-2 (рис. 3C). Предыдущие исследования, проведенные нами и другими группами, показали, что различные формы WC-1 и WC-2 с разным сдвигом подвижности являются
все из-за событий фосфорилирования (Talora et al. 1999; He et al. 2005b; Schafmeier et al. 2005). Как и предполагалось, аналогично штамму frq 10 , и WC-1, и WC-2 гипофосфорилированы в мутантах sFRQ4, FRQ4A11 и FRQ4A12 (рис. 2B, C), в которых FCD либо удален, либо мутирован точечные мутации.Кроме того, уровни WC-1 и WC-2 у этих мутантов
сопоставимы с таковыми в штамме frq 10 , но значительно ниже, чем у штамма дикого типа, что указывает на то, что петли положительной обратной связи также нарушены
нарушением взаимодействия FRQ / CK-1a.
Рисунок 3.
FRQ, WC-1 и WC-2 гипофосфорилированы в штамме ck-1a L , нокаут-мутанте, несущем мутацию dbt L .( A ) Графическая диаграмма, изображающая процедуры создания гомокариотического штамма ck-1a L путем гомологичной рекомбинации (подробности см. В разделе «Материалы и методы»). Звездочка в ORF ck-1a указывает местоположение мутации dbt L . ( B ) Саузерн-блот-анализ, показывающий вставку hph в эндогенный локус ck-1a . Геномную ДНК переваривали с помощью SphI, и мембрану зондировали зондом, специфичным для ck-1a .( C ) Вестерн-блоттинг показывает гипофосфорилирование FRQ, WC-1 и WC-2 в штамме ck-1a L . Культуры выращивали в LL. Стрелки указывают на сильно фосфорилированные виды FRQ или WC, отсутствующие у мутантов.
Числа справа указывают соотношение акриламид / бисакриламид, используемое в SDS-PAGE. Обратите внимание, что для WC-2 обычный SDS-PAGE (37,5: 1) не прошел
для разрешения различных WC-2-фосфорилированных форм.( D ) Вестерн-блоттинг, демонстрирующий стабильность FRQ, WC-1 и WC-2 после добавления CHX (10 мкг / мл) в диком типе
и штаммов ck-1a L . Культуры выращивали в LL. ( E ) Денситометрический анализ четырех независимых экспериментов, показывающий деградацию FRQ в штаммах дикого типа и ck-1a L после добавления CHX.
Чтобы исключить возможность того, что нарушение фосфорилирования WC и низкие уровни WC у этих мутантов связаны с отменой
Взаимодействия FRQ – WCC мы исследовали взаимодействие FRQ – WCC у мутантов FRQ4A11 и FRQ4A12.Как показано на рисунке 2D, иммунопреципитация WC-2 снижает FRQ в штаммах дикого типа, а также в двух мутантных штаммах, указывая на то, что взаимодействие FRQ-WCC
не зависит от взаимодействия FRQ / CK-1a. Следовательно, эти результаты предполагают, что CK-1a фосфорилирует белки WC в зависимости от FRQ.
способ через его тесную связь с FRQ. Кроме того, взаимодействие FRQ / CK-1a или фосфорилирование WC опосредовано
такое взаимодействие важно для функции FRQ, способствующей накоплению белков WC-1 и WC-2.
Для дальнейшего подтверждения важности взаимодействия FRQ / CK-1a, циркадные конидиционные ритмы штаммов FRQ4A11 и FRQ4A12
были исследованы с помощью пробирок с гоночными пробирками. Как показано на Фигуре 2E, аналогично штамму frq 10 , эти мутанты демонстрировали постоянную конидиацию в постоянной темноте без видимой ритмичности. Таким образом, мы делаем вывод
что взаимодействие FRQ / CK-1a необходимо для нормальной функции циркадных часов.
Создание нокаутного мутанта Neurospora ck-1a, несущего мутацию dbt
L
Хотя результаты, представленные выше, предполагают важную роль CK-1a в фосфорилировании FRQ и WC, дальнейшее подтверждение
этому выводу препятствовало отсутствие генетических доказательств. ck-1a является важным геном в Neurospora (Gorl et al.2001), и нам не удалось получить гомокариотический нокаут-штамм Neurospora ck-1a (Q. He and Y. Liu, неопубликовано). Мутант Drosophila dbt L имеет единственную мутацию Met в Ile по консервативному остатку Met в киназном домене DBT (Kloss et al. 1998; Price et al. 1998). Такая мутация приводит к снижению активности DBT и увеличению продолжительности периода часов (Preuss et al. 2004). Поскольку плодовые мушки, несущие мутацию dbt L , были жизнеспособными, но демонстрировали сильный циркадный фенотип, мы надеялись, что эквивалентная мутация в Neurospora ck-1a также будет совместима с выживаемостью клеток и, возможно, повлияет на циркадные часы.Чтобы получить такого мутанта, подбивку
Кассета была сделана из геномной ДНК, которая содержит мутантный ген ck-1a (консервативный Met83 был изменен на Ile) и ген устойчивости к гигромицину ( hph ), вставленный ниже 3 ‘нетранслируемой области (UTR) ck-1a (Рис. 3A). Затем эту кассету трансформировали в штамм bd ku-70 rip , который является дефектным в негомологической рекомбинации (Ninomiya et al. 2004), и было отобрано hph -устойчивых трансформантов.Секвенирование эндогенного гена ck-1a трансформантов использовали для идентификации гетерокарионных штаммов, в которых мутированная кассета ck-1a была интегрирована посредством гомологичной рекомбинации. Гомокариотический штамм с мутацией ck-1a dbt L , обозначенный как ck-1a L (на фоне bd ku-70 rip ), был затем получен путем очистки микроконидий.
Анализ методом Саузерн-блоттинга
подтвердил интеграцию кассеты knock-in в эндогенном локусе ck-1a и гомокариотическую природу штаммов ck-1a L (рис.3Б). Секвенирование ДНК эндогенного гена ck-1a дополнительно подтвердило мутированный ck-1a в этих штаммах (данные не показаны). Штаммы ck-1a L демонстрируют более медленный рост и образуют меньше воздушных гиф и конидий, чем штамм дикого типа. Штамм bd ku-70 rip демонстрирует нормальный рост, развитие и циркадные ритмы, как штамм bd дикого типа, что указывает на то, что мутация ku-70 rip не влияет на эти фенотипы в ck-1a L штаммов (данные не представлены).Поскольку hph интегрирован ниже по течению от ck-1a 3’UTR, области без прогнозируемых генов, фенотипы роста и развития штаммов ck-1a L предполагают, что они обусловлены частичной функцией CK-1a, вызванный мутацией dbt L .
Насколько нам известно, это первый штамм с точечной мутацией, созданный в Neurospora .Описанный здесь подход будет очень полезен при изучении функций основных генов в этом организме.
Гипофосфорилирование FRQ, WC-1 и WC-2 и снижение скорости деградации FRQ в штамме ck-1a
L
Затем мы сравнили профили фосфорилирования FRQ, WC-1 и WC-2 в штаммах дикого типа и ck-1a L для культур, выращенных в LL.Как показано на фиг. 3C, все три белка гипофосфорилированы в штамме ck-1a L . Эти результаты предоставляют убедительные доказательства in vivo, предполагающие, что CK-1a фосфорилирует FRQ, WC-1 и WC-2 in vivo. В
кроме того, оба уровня WC-1 и WC-2 у мутанта были ниже, чем у штамма дикого типа, что указывает на нарушение
петли положительной обратной связи.
Мутация S513 в FCD и делеция домена PEST-1 приводят к повышению стабильности FRQ (Liu et al.2000; Gorl et al. 2001), предполагая, что CK-1a фосфорилирование FRQ регулирует его стабильность. Чтобы проверить эту гипотезу, мы сравнили устойчивость
FRQ в штаммах дикого типа и ck-1a L после добавления ингибитора синтеза белка циклогексимида (CHX). Как показано на фиг. 3, D и E, скорость деградации FRQ была ниже в штамме ck-1a L , что указывает на то, что фосфорилирование FRQ СК-1a способствует его деградации. Напротив, нет существенной разницы
в скоростях разложения WC-1 и WC-2 между штаммами дикого типа и ck-1a L , что позволяет предположить, что низкие уровни WC-1 и WC-2 в штаммах ck-1a L не только из-за изменения их стабильности.
Длиннопериодные циркадные ритмы и гипофосфорилированные белки WC при DD у штамма ck-1a
L
Низкая скорость деградации FRQ в штамме ck-1a L предполагает, что он должен проявлять долгопериодный циркадный фенотип. Как и ожидалось, на гоночных камерах в постоянной темноте
(DD), несмотря на меньшую конидиацию, чем штамм дикого типа, штамм ck-1a L демонстрирует низкоамплитудный циркадный ритм конидиации с периодом ~ 32 ч при комнатной температуре в течение ~ 6-7 дней. , Период
на ~ 10 ч больше, чем у штамма дикого типа (рис.4А). Кроме того, фаза первой полосы конидиации после перехода свет / темнота (LD) резко задерживалась в
ck-1a L штамм (~ 19 ч после перехода LD) по сравнению со штаммом дикого типа (~ 10 ч после). При циклах LD 12/12, несмотря на
При длительном периоде DD ритм конидиации штамма ck-1a L мог быть увлечен до 24 часов, но их пики полос конидиации (около ZT16) задерживались на ~ 5 часов по сравнению с
штамм дикого типа (около ZT11) (рис.4Б).
Рисунок 4.
Анализы
с гоночной трубкой, показывающие, что штаммы ck-1a L демонстрируют длительный ритм конидиации с отсроченной фазой. ( A ) Результаты в гоночной трубе, показывающие ритмы конидиации дикого типа и двух независимых штаммов ck-1a L в DD. Результаты денситометрии, отражающие плотность конидиации, показаны на ниже пробирок.Звездочками отмечен пик полос конидий у штаммов ck-1a L . ( B ) Результаты в гоночной трубе, показывающие ритмы конидиации штаммов дикого типа и ck-1a L в циклах LD (12/12).
Затем мы исследовали колебания FRQ в DD. В штамме дикого типа, аналогично показанному ранее, FRQ показал устойчивый
~ 22-часовые ритмы как по количеству, так и по профилям фосфорилирования (рис.5А). В штамме ck-1a L также были видны ритмы величин FRQ и профили фосфорилирования FRQ, но с значительно более длительным периодом, чем у штамма ck-1a L .
дикого типа. На долгопериодные колебания FRQ у мутанта указывают колебания количества FRQ и фосфорилирования FRQ.
профили. Гипофосфорилированный FRQ (продукты вновь синтезированного FRQ) появился в DD16 и DD48 у мутанта, что соответствует
с его 32-часовым ритмом конидиации.Кроме того, общие уровни FRQ в DD в штамме ck-1a L сопоставимы или немного выше, чем у штамма дикого типа.
Рисунок 5.
Долгопериодический ритм FRQ и гипофосфорилирование WC-1 и WC-2 в штамме ck-1a L . ( A ) Вестерн-блоттинг, показывающий колебания FRQ в штаммах дикого типа и ck-1a L в DD.Денситометрический анализ результатов вестерн-блоттинга показывает ниже . ( B ) Вестерн-блоттинг, показывающий параллельное сравнение профилей фосфорилирования WC-1 и WC-2 в штаммах дикого типа и ck-1a L в разные моменты времени в DD. (W) штамм дикого типа; (M) штамм ck-1a L . ( C ) Нозерн-блот-анализ, показывающий колебания frq РНК в штаммах дикого типа и ck-1a L в DD.Денситометрический анализ результатов Нозерн-блоттинга показан на справа .
После начального ухудшения FRQ после перехода LD произошла небольшая (<4 ч) задержка в начале нового
цикл синтеза FRQ (обозначенный гипофосфорилированным FRQ в обоих штаммах на DD16) и пиком количества FRQ в
мутант по сравнению со штаммом дикого типа, предполагая, что активация транскрипции frq в новом цикле существенно не задерживается у мутанта.Напротив, наиболее значительная задержка колебания FRQ в
мутант, по-видимому, представляет собой процесс прогрессирующего фосфорилирования и деградации FRQ. В штамме дикого типа FRQ с
его промежуточные фосфорилированные формы достигают пика при DD20 и становятся гиперфосфорилированными и разлагаются с DD24-DD32 в процессе
это занимает 12 ч. Однако в штамме ck-1a L такой процесс занял ~ 20 часов; гипофосфорилированный и промежуточно фосфорилированный FRQ по DD20 не
становятся гиперфосфорилированными до DD40.Эти данные свидетельствуют о том, что нарушение активности CK-1a резко замедляет процесс.
прогрессирующего фосфорилирования FRQ в штамме ck-1a L .
Для сравнения профилей фосфорилирования WC-1 и WC-2 в штаммах дикого типа и ck-1a L в DD мы провели параллельные сравнения культур, собранных в разные моменты времени в DD ( Рис. 5Б). Помимо фосфорилирования в темноте, фосфорилирование белков WC может дополнительно стимулироваться воздействием света.
(Талора и др.1999; Швердтфегер и Линден 2000; Он и Лю 2005). Таким образом, WCs становятся гиперфосфорилированными после светового импульса или в LL. Для штамма дикого типа и WC-1, и WC-2 стали дефосфорилированными.
после перехода LD, вероятно, из-за действия протеинфосфатазы (ей), такой как PP2A (Schafmeier et al. 2005), и остается в промежуточных состояниях фосфорилирования в DD (Fig. 5B). Ритмы фосфорилирования WC в DD не были очевидны из-за сдвигов их подвижности в одномерном SDS-PAGE анализах.
из-за пределов чувствительности метода (Schafmeier et al.2005). В штаммах ck-1a L WC-1 и WC-2 были гипофосфорилированы во все моменты времени, что указывает на то, что CK-1a является основной киназой, фосфорилирующей
Белки WC в DD и LL. Для образца LL, хотя он гипофосфорилирован по сравнению со штаммом дикого типа, некоторые WC-2-
фосфорилированные формы можно увидеть в мутанте, что указывает на то, что дополнительная киназа (-ы) также отвечает за светозависимую
Фосфорилирование WC.Кроме того, уровни WC-1 и WC-2 у мутанта были ниже, чем у штамма дикого типа в
DD, что указывает на нарушение циркадных контуров положительной обратной связи при нарушении функции CK-1a.
Несмотря на низкие уровни белков WC в штамме ck-1a L , уровни его РНК frq в DD были сравнимы с таковыми в штамме дикого типа и демонстрировали долгопериодный ритм (рис.5С). Подобно ритмам белка FRQ, наблюдалась лишь небольшая задержка пика мРНК frq в штамме ck-1a L , тогда как наблюдалась длительная задержка в фазе убывания РНК frq . Эти результаты предполагают, что гипофосфорилированные белки WC в мутанте могут быть более активными в активации транскрипции frq , чем штамм дикого типа, что согласуется с предыдущими результатами (He and Liu 2005; He et al. 2005b; Schafmeier et al.2005). Кроме того, длительная задержка фазы уменьшения РНК frq в штамме ck-1a L предполагает, что циркадная отрицательная обратная связь серьезно нарушена, несмотря на наличие нормальных или незначительных
более высокие суммы FRQ.
WCC ритмично связывается с C-боксом frq in vivo
В штамме Neurospora дикого типа профили фосфорилирования WC-2 показывают циркадный ритм, причем время гипофосфорилирования WC-2 коррелирует с
с моментом активации frq (Schafmeier et al.2005). Анализ связывания ДНК in vitro ранее показал, что способность WCC связываться с frq C-box колеблется ежедневно, а дефосфорилирование WCC увеличивает его связывание с ДНК (Froehlich et al. 2003; He and Liu 2005). Чтобы понять, как фосфорилирование WCC регулирует его связывание с промотором frq в циркадном цикле, мы исследовали связывание in vivo WCC с frq C-box в DD с помощью полуколичественного анализа иммунопреципитации хроматина (ChIP). На рисунке 6A показано колебание FRQ для образцов, используемых в анализе ChIP.Как показано на Рисунке 6B, WCC ритмично связывается с C-боксом frq , достигая пика на DD14 и DD36, что соответствует времени пиков мРНК frq (Рис. 5C; Aronson et al. 1994; Garceau et al. 1997). ). Эти временные точки также коррелируют со временем, когда WC-2 наименее фосфорилируется (Schafmeier et al. 2005), предполагая, что гипофосфорилированные виды WCC являются его наиболее активной формой. Таким образом, аналогично продемонстрированной ситуации
у Drosophila и мыши положительные ветви часов ритмично связываются с промотором отрицательных элементов (Ripperger and Schibler 2006; Yu et al.2006 г.).
Рисунок 6.
WCC связывается с С-боксом frq ритмично в DD в штамме дикого типа, и связывание не может быть эффективно ингибировано FRQ в штамме ck-1a L . ( A ) Вестерн-блоттинг, показывающий ритмическую экспрессию FRQ в DD в штамме дикого типа. ( B ) Анализ ChIP с использованием антитела WC-2, показывающий ритмическое связывание WCC с C-боксом frq .Денситометрический анализ результатов Western и ChIP показывает ниже . ( C ) Анализ ChIP, показывающий связывание WCC с frq C-box в штаммах дикого типа и ck-1a L в разные моменты времени. Денситометрический анализ результатов ChIP показывает ниже . ( D ) Нозерн-блоттинг, показывающий индуцированную светом транскрипцию frq в штаммах дикого типа и ck-1a L .Культуры выращивали в DD в течение 24 ч перед переносом в LL.
Сравнение ритмов экспрессии FRQ и связывания C-бокса WCC показало, что пики связывания C-бокса WCC достигаются, когда белок FRQ
был в минимуме, предполагая, что ингибирование WCC с помощью FRQ высвобождается, когда уровни FRQ низкие. Кроме того, после
был получен вновь синтезированный FRQ, связывание WCC с промотором frq было быстро снижено (обратите внимание на значительное падение связывания на DD18) и оставалось на низком уровне до реактивации
ВСЦ.Эти результаты показывают, что ингибирование активности WCC с помощью FRQ происходит быстро в штамме дикого типа, что согласуется с
предыдущие результаты (Merrow et al. 1997; Schafmeier et al. 2006).
Связывание
WCC с C-боксом не может эффективно ингибироваться FRQ в штамме ck-1a
L
Гипофосфорилированные белки WC в штамме ck-1a L и его длительная задержка в фазе уменьшения РНК frq позволяют предположить, что FRQ не может эффективно ингибировать активность WCC у этого мутанта.Чтобы проверить эту гипотезу, мы провели бок о бок
сравнение связывания WCC с C-боксом frq в штаммах дикого типа и мутантных штаммах. Как показано на Фигуре 6C, в отличие от дикого типа, у которого связывание быстро снижалось на DD18, связывание WCC с C-боксом оставалось на высоком уровне.
в течение по крайней мере 8 часов в штамме ck-1a L , несмотря на присутствие высоких уровней FRQ (фиг. 5A). Эти данные предполагают, что циркадная отрицательная обратная связь серьезно нарушена у мутанта в результате
неспособность мутанта CK-1a эффективно фосфорилировать белки WC.
Ранее мы показали, что индуцированное светом фосфорилирование WCC подавляет его роль в качестве фактора транскрипции для индуцированного светом
транскрипция (He and Liu 2005). Гипофосфорилированные белки WC в LL в штамме ck-1a L предполагают, что CK-1a также частично участвует в индуцированном светом фосфорилировании WC. Таким образом, мы ожидали, что
кинетика индуцированной светом транскрипции будет изменена у мутанта.Чтобы дополнительно продемонстрировать, что гипофосфорилированный
WCC является активной формой WCC, мы исследовали световую индукцию транскрипции frq в штамме ck-1a L (фиг. 6D). Как ранее было показано для штамма дикого типа (Crosthwaite et al. 1995; He and Liu 2005), хотя низкий уровень индуцированной светом РНК frq можно было увидеть через 5 минут после воздействия света, frq достиг пика через 15 минут. –30 мин, а затем уменьшилась в результате фотоадаптации (He and Liu 2005).Напротив, в штамме ck-1a L светоиндуцированная РНК frq достигла своего пика на уровне, который был выше, чем пиковый уровень дикого типа, всего за 5 минут и медленно снижался.
впоследствии, что указывает на то, что у мутанта частично нарушен процесс фотоадаптации. Эти результаты и низкие
Уровни белков WC в штамме ck-1a L позволяют предположить, что гипофосфорилированный WCC также является активной формой, которая опосредует индуцированную светом транскрипцию.Следовательно,
CK-1a также играет роль в световом ответе и фотоадаптации у Neurospora .
Нарушение cka, гена каталитической субъединицы CKII, приводит к гипофосфорилированию белков WC и нарушению FRQ
для подавления активности WCC
Медленное уменьшение мРНК frq в штамме ck-1a L предполагает, что в ингибировании WCC с помощью FRQ участвует дополнительный механизм (рис.5С). Присутствие светозависимых форм фосфорилирования WC в штамме ck-1a L (фиг. 5В) указывает на то, что в фосфорилировании WC участвует дополнительная (ые) киназа (ы). Ранее мы идентифицировали CKII как главную киназу
который фосфорилирует FRQ in vivo. Нарушение его каталитической субъединицы cka и одной из его регуляторных субъединиц показало, что у этих мутантов влияет не только стабильность FRQ, но и отрицательная обратная связь.
петля также серьезно нарушена (Yang et al.2002, 2003).
Чтобы понять, как CKII функционирует в этом процессе, мы исследовали профили фосфорилирования WC-1 и WC-2 в мутанте cka RIP . Как показано на фиг. 7A, и WC-1, и WC-2 были гипофосфорилированы в DD и LL в мутанте cka RIP , что позволяет предположить, что CKII является другой киназой, которая может опосредовать фосфорилирование WC-1 и WC-2. Однако в отличие от
ck-1a L , уровни WC не были снижены у мутанта cka RIP , что указывает на то, что CKII не участвует в регуляции петель положительной обратной связи.Кроме того, как и ранее
Как показано, уровни FRQ были высокими и гипофосфорилированы у мутанта cka RIP (Yang et al. 2002).
Рисунок 7.
WC-1 и WC-2 гипофосфорилированы, и WCC связывается с C-боксом на постоянных высоких уровнях в штамме cka RIP .( A ) Вестерн-блоттинг показывает профили фосфорилирования WC-1 и WC-2 в штаммах дикого типа и cka RIP . Стрелки указывают на сильно фосфорилированные формы WC-1, WC-2 и FRQ, отсутствующие в мутантном штамме. ( B ) Анализ ChIP, показывающий связывание WCC с frq C-box в штаммах дикого типа и cka RIP в разные моменты времени. Денситометрический анализ результатов ChIP показывает ниже .( C ) Нозерн-блоттинг, показывающий индуцированную светом транскрипцию frq в штаммах дикого типа и cka RIP . Культуры выращивали в DD в течение 24 ч перед переносом в LL.
Чтобы исследовать, являются ли гипофосфорилированные WC в мутанте cka RIP также результатом неспособности FRQ ингибировать активность WCC, мы сравнили связывание C-бокса WCC в генах дикого типа.
и мутантные штаммы.Как показано на Фигуре 7B, в то время как связывание WCC в DD в штамме дикого типа демонстрирует поведение, аналогичное показанному ранее, у мутанта WCC
привязка к C-box оставалась на более высоком уровне в эти моменты времени, несмотря на присутствие большого количества FRQ. Таким образом, FRQ
не может эффективно ингибировать активность WCC в отсутствие CKII. Хотя CKII не связывается с FRQ прочно (Yang et al. 2002, 2003), временные взаимодействия между FRQ и CKII во время процесса фосфорилирования FRQ приведут к тому, что CKII окажется поблизости.
белков WC через взаимодействие FRQ – WCC.Таким образом, подобно CK-Ia, CKII может также опосредовать фосфорилирование WCC.
в зависимости от FRQ.
Затем мы исследовали, приводят ли гипофосфорилированные WCs в мутанте cka RIP к изменениям кинетики индуцированной светом транскрипции frq . Как показано на Фигуре 7C, аналогично штамму ck-1a L , индуцированные светом уровни frq достигли пика через 5 мин у мутанта cka RIP и повысились до более высокого уровня, чем у мутанта штамм дикого типа.Эти данные позволяют предположить, что гипофосфорилированный WCC
в cka RIP мутант также является активной формой WCC для функции часов и светового ответа, и что CKII также играет роль в регуляции
Активность туалета после воздействия света.
Обсуждение
В этом исследовании наши результаты показывают, что CK-1a и CKII не только две из основных киназ, фосфорилирующих FRQ, но и
также ключевые киназы, которые опосредуют FRQ-зависимое фосфорилирование WCC, чтобы закрыть циркадную петлю отрицательной обратной связи.Кроме того, CK-1a также важен для FRQ-зависимого накопления WC-1 и WC-2.
CK-1a фосфорилирует FRQ in vivo, что способствует деградации FRQ и важно для циркадных петель положительной обратной связи
Ранее CKII была единственной известной основной FRQ-фосфорилирующей киназой in vivo (Yang et al. 2002, 2003). Хотя было показано, что CK-1a связан с FRQ и может фосфорилировать FRQ in vitro, его роль in vivo в фосфорилировании FRQ
неизвестно (Gorl et al.2001; Cheng et al. 2005; Schafmeier et al. 2006 г.). Здесь мы представили убедительные доказательства того, что CK-Ia является киназой, которая фосфорилирует FRQ in vivo. Делеции или мутации
FCD, которые нарушают взаимодействие между FRQ и CK-1a, приводят к гипофосфорилированию FRQ, указывая на то, что
количество событий фосфорилирования FRQ требует взаимодействия FRQ / CK-1a. Что еще более важно, FRQ гипофосфорилируется в
ck-1a L , и процесс прогрессирующего фосфорилирования FRQ у мутанта значительно задерживается.Подобно функции CKI
у Drosophila и млекопитающих (Price et al. 1998; Akashi et al. 2002; Eide et al. 2005a) фосфорилирование FRQ с помощью CK-1a способствует деградации FRQ. Поскольку стабильность FRQ является основным фактором, определяющим период
длина часов в Neurospora (Liu et al. 2000; Gorl et al. 2001; Yang et al. 2003), повышенная стабильность FRQ и значительно замедленный процесс прогрессирующего фосфорилирования FRQ привели к 32-часовому
циркадные ритмы у штамма ck-1a L .
Интересно, что три ранее идентифицированных сайта фосфорилирования FRQ и домен PEST-1 расположены непосредственно ниже по течению.
из FCD (Liu et al. 2000; Gorl et al. 2001), предполагая, что они, вероятно, фосфорилируются CK-1a in vivo. Также очевидно, что значительные количества фосфорилирования FRQ
остаются в мутантах FCD и в штамме ck-1a L , что указывает на участие других киназ, таких как CKII и CAMK-1 (Yang et al.2001, 2002, 2003). Однако также возможно, что некоторые из событий фосфорилирования FRQ в мутанте FCD могут опосредоваться независимым от CK-1a.
тесного взаимодействия FRQ / CK-1a.
FRQ способствует накоплению как WC-1, так и WC-2, формируя петлю положительной обратной связи, которая важна для устойчивости и
функция циркадных часов в Neurospora (Lee et al.2000; Cheng et al.2001b; Schafmeier et al. 2006 г.). Мутация сайтов фосфорилирования FRQ in vitro с помощью CK-1a в домене PEST-2 предполагает, что эти события фосфорилирования
важны для накопления WC-1, но не WC-2 (Schafmeier et al. 2006). Наши результаты, представленные здесь, показали, что CK-1a важен для циркадных петель положительной обратной связи in vivo. Уровни
как WC-1, так и WC-2 низки у мутантов FCD и у штамма ck-1a L (рис.2,3,5). Эти результаты предполагают, что фосфорилирование FRQ с помощью CK-1a важно не только для накопления WC-1, но и
важен для экспрессии WC-2. Кроме того, роль CK-1a в фосфорилировании WCC (обсуждается ниже) повышает вероятность
что такие посттрансляционные модификации могут также способствовать накоплению белков WC. В соответствии с этим представлением,
мутации идентифицированных сайтов фосфорилирования WC-1 вблизи области его Zn-пальца приводят к низким уровням WC-1 (He et al.2005b).
Напротив, хотя CKII также является основной киназой, фосфорилирующей FRQ in vivo, и выполняет функции, аналогичные CK-1a в FRQ
деградации и в петле циркадной отрицательной обратной связи он не требуется для петель циркадной положительной обратной связи (рис. 7; Yang et al. 2002). Это функциональное различие между CK-1a и CKII предполагает, что они могут фосфорилировать функционально разные сайты на FRQ.
или туалеты.
CK-Ia и CKII опосредуют FRQ-зависимое фосфорилирование WCC, чтобы закрыть циркадную петлю отрицательной обратной связи
Наши результаты, представленные здесь, предполагают, что CK-1a и CKII опосредуют FRQ-зависимое фосфорилирование WC и ингибируют связывание C-бокса WCC.
Мероприятия. Для CK-1a FRQ опосредует фосфорилирование CK-1a WCC через его ассоциацию с CK-1a и WCC.Во-первых, нарушение
взаимодействия FRQ / CK-1a путем делеции или точечных мутаций FCD приводит к гипофосфорилированию WC-1 и WC-2 (рис. 2B, C), указывая на то, что взаимодействие FRQ / CK-1a необходимо для нормального фосфорилирования туалетов. Во-вторых, СК-1а и ВК-1 являются
присутствуют в том же комплексе в зависимости от FRQ (рис. 2А). Наконец, в штамме ck-1a L WC-1 и WC-2 гипофосфорилированы, несмотря на присутствие частично функционального CK-1a (рис.3Д, 5Б).
WC-1 и WC-2 также гипофосфорилируются у мутантов cka (фиг. 7A), что позволяет предположить, что CKII может быть другой киназой, опосредующей фосфорилирование WC. В отличие от CK-1a, CKII не связывает прочно
с FRQ (Yang et al. 2002, 2003), указывая на то, что их взаимодействие является слабым и временным, как и большинство взаимодействий киназа-субстрат. Точно так же CKII и WCC
не было обнаружено, что они тесно связаны друг с другом при иммунопреципитации (данные не показаны).Переходное взаимодействие
между FRQ и CKII во время процесса фосфорилирования FRQ может приближать CKII к WCC посредством взаимодействия FRQ-WCC.
Таким образом, CKII может также опосредовать фосфорилирование WC зависимым от FRQ образом. Однако также возможно, что CKII может
опосредуют некоторые события фосфорилирования WC независимо от FRQ.
Взятые вместе, эти результаты предполагают, что FRQ действует как субъединица, рекрутирующая субстрат киназы, чтобы опосредовать фосфорилирование
WC по CK-1a и CKII (рис.8). Какова роль фосфорилирования WC этими двумя киназами? Анализы ChIP показали, что в штамме дикого типа WCC связывает
к frq C-box ритмично. Пики связывания WCC возникают, когда уровень FRQ находится на минимальном уровне, но связывание быстро ингибируется.
увеличением суммы FRQ и остается на низком уровне большую часть дня. Напротив, ингибирование связывания C-бокса WCC
by FRQ сильно нарушена у мутантов ck-1a L и cka (фиг.6,7). В штамме ck-1a L , несмотря на низкие уровни белков WC, WCC связывается с C-боксом на высоком уровне и не может подавляться даже FRQ.
при высоком уровне FRQ. В мутанте cka RIP связывание WCC в C-боксе сохраняется на более высоком уровне, чем у дикого типа при DD, несмотря на то, что он значительно выше, чем у дикого типа.
Уровень FRQ, что приводит к высоким уровням frq РНК. На усиление активности WCC в мутантах ck-1a, L и cka дополнительно указывает быстрая и повышенная светоиндуцированная транскрипция frq в этих штаммах.Таким образом, эти результаты показывают, что фосфорилирование белков WC этими двумя киназами
подавляет активность WCC и имеет важное значение для циркадной отрицательной обратной связи.
Рисунок 8.
Обновленная модель петли циркадной отрицательной обратной связи Neurospora . См текст для деталей.
Участие как CK-1a, так и CKII в фосфорилировании WCC предполагает, что эти две киназы могут действовать совместно. CKI
и CKII имеют разные консенсусные последовательности субстрата: (D / E / S p / T p ) XX (S / T) для CKI и (S / T) XX (D / E / S p / T p ) для CKII (p обозначает фосфорилированный остаток).Таким образом, возможно, что каждый из них фосфорилирует отдельные сайты на
Белки WC. С другой стороны, поскольку они оба предпочитают сайты, окруженные кислотными или фосфорилированными остатками, также вероятно
что одна киназа может действовать как прайминговая киназа для другой (или других неизвестных киназ) для достижения надлежащего фосфорилирования
Белки WC, аналогичные случаю, предполагаемому мутацией при синдроме продвинутой фазы сна у человека (FASPS) (Toh et al.2001). Сайты WC, фосфорилируемые CK-1a и CKII, в настоящее время неизвестны. Ранее мы идентифицировали пять основных in vivo WC-1
сайты фосфорилирования расположены непосредственно ниже его ДНК-связывающего домена (He et al. 2005b). В соответствии с результатами, представленными здесь, мутация этих сайтов предполагает, что фосфорилирование негативно регулирует
функция WCC. Однако эти сайты не похожи на типичные консенсусные сайты CKI или CKII и два из фосфорилированных
за серинами следует остаток пролина.Хотя CKI и CKII широко не известны как пролин-направленные киназы, пролин-направленные киназы
Сообщалось о фосфорилировании CKI in vivo (Alappat et al. 2005). Таким образом, возможно, что CK-1a может быть киназой, которая может фосфорилировать эти пять сайтов WC-1 in vivo.
На рисунке 8 представлена обновленная модель циркадной петли обратной связи Neurospora , основанная на этом и предыдущих исследованиях с акцентом на фосфорилирование WC.В субъективном начале
утром в DD уровень FRQ низкий и WC гипофосфорилированы. Гипофосфорилированный WCC активен и связывает C-бокс
для активации транскрипции frq (Cheng et al. 2001b; Froehlich et al. 2002, 2003). После образования белка FRQ гомодимерный FRQ образует комплекс FRH и связывается с CK-1a (прочно) (Cheng et al. 2001a, 2005; Gorl et al. 2001) и CKII (слабо). После этого FFC взаимодействует с WCC и переносит CK-1a и CKII в непосредственной близости от WCC к фосфорилату.
белки WC (Cheng et al.2001a; Denault et al. 2001). Когда уровень FRQ высок, это приводит к обширному фосфорилированию и инактивации WCC (He and Liu 2005; He et al. 2005b; Schafmeier et al. 2005, 2006). Этот процесс удаляет WCC с промотора frq и приводит к снижению транскрипции frq . В то же время FRQ прогрессивно фосфорилируется CK-1a и CKII и подвергается интенсивному фосфорилированию.
Затем экстенсивно фосфорилированный FRQ распознается убиквитин-лигазой SCF FWD-1 E3, что приводит к убиквитинированию и деградации FRQ через протеасомный путь (He et al.2003, 2005а). После того, как FRQ снижается до определенного уровня, неактивный гиперфосфорилированный WCC дефосфорилируется PP2A и, вероятно,
др. PPases (Schafmeier et al. 2005), что приводит к реактивации WCC и транскрипции frq в новом циркадном цикле. В этой модели количество FRQ определяет количество киназ, которые могут быть задействованы.
фосфорилировать WCC и, таким образом, степень фосфорилирования WCC и его активность. Следовательно, колебания FRQ существенны.
для функции этой циркадной отрицательной обратной связи, а стабильность FRQ играет важную роль в определении продолжительности периода.
(Лю и др.2000; Gorl et al. 2001; Ян и др. 2003; Ruoff et al. 2005).
Сохранение посттрансляционной регуляции циркадных систем эукариот
Сохранение посттрансляционных механизмов замечательно среди различных циркадных систем эукариот, происходящих от грибов.
людям (Heintzen and Liu 2006). Подобно FRQ и WCs, основные тактовые компоненты животных и растений также регулируются посредством фосфорилирования (Young and Kay 2001).Все белки FRQ и PER животных фосфорилируются CKI и CKII, дефосфорилируются одними и теми же фосфатазами и
разрушается системой убиквитин / протеасома с использованием консервативной убиквитинлигазы E3 (Kloss et al. 1998; Price et al. 1998; Lowrey et al. 2000; Gorl et al. 2001; Lee et al. 2001; Grima et al. 2002; Ко и др. 2002; Лин и др. 2002; Янг и др. 2002, 2003, 2004; Актен и др. 2003; Хе и др. 2003; Наватин и Росбаш 2004; Сатьянараян и др. 2004; Эйде и др.2005b; Xu et al. 2005). Подобно Neurospora , CKI, как было обнаружено, также тесно связан с белками PER у Drosophila и млекопитающих (Kloss et al. 1998; Lee et al. 2001, 2004). Было высказано предположение, что у мышей взаимодействие PER / CKI является критическим процессом в регуляции фосфорилирования mPER и функции
часов (Ли и др. 2004). Подобно роли CKI в часовых системах животных (Price et al. 1998; Akashi et al. 2002; Eide et al. 2005a), мы показали здесь, что штамм ck-1a L , который несет ту же мутацию как мутанты Drosophila long period dbt L , проявляющие долгопериодные ритмы и нарушенную деградацию FRQ.У людей два типа семейной продвинутой фазы сна.
Было обнаружено, что синдром (FASPS) возникает из-за мутаций человеческого CKIδ или его сайтов фосфорилирования на hPER2 (Toh et al. 2001; Xu et al. 2005). Для CKII, как мы показали в Neurospora, , также было обнаружено, что он необходим для репрессорной функции PER у Drosophila (Nawathean and Rosbash 2004), хотя как CKII функционирует, способствуя репрессорной активности PER, неясно. В Arabidopsis , где CKII впервые участвовал в работе часов, было показано, что он фосфорилирует CCA1 и регулирует его связывание с ДНК.
активность (Sugano et al.1998).
Как и белки WC, факторы транскрипции, содержащие PAS-домен, которые действуют как положительные конечности циркадного ритма животных.
петли отрицательной обратной связи также регулируются с помощью фосфорилирования, а BMAL1 и CLK млекопитающих и Drosophila CLK также обнаруживают устойчивые ритмы в своих профилях фосфорилирования (Lee et al. 2001; Kim and Edery 2006; Yu et al. 2006). Более примечательно, что, как и в случае с Neurospora , недавно было обнаружено, что фосфорилирование CLK зависит от PER и может опосредоваться DBT (Lee et al.2001; Ким и Эдери 2006; Yu et al. 2006 г.). Кроме того, связывание CLK – CYC с на E-box коррелирует с накоплением гипофосфорилирования CLK, а гиперфосфорилирование CLK коррелирует с
пик на репрессию мРНК (Kim and Edery 2006; Yu et al. 2006). Кроме того, было показано, что PP2A участвует в регуляции фосфорилирования CLK (Kim and Edery 2006). В культивируемых клетках S2 DBT-зависимое фосфорилирование CLK способствует деградации CLK.Хотя роль фосфорилирования CLK
посредством DBT в регуляции ДНК-связывающей активности CLK – CYC все еще отсутствует, и механизм работы CKII неизвестен.
у Drosophila наши результаты, представленные в этом исследовании, и сохранение циркадных систем предполагают, что аналогичный механизм для
закрытие петли отрицательной обратной связи существует у Neurospora , Drosophila и, вероятно, у млекопитающих.
Материалы и методы
Штаммы и условия культивирования
87-3 ( bd a ) использовали в качестве штамма дикого типа в этом исследовании. В этом исследовании были созданы штаммы ku-70 RIP и ck-1a L .Штамм cka RIP , в котором ген cka был нарушен множественными преждевременными стоп-кодонами, был создан ранее (Yang et al. 2002). Штамм 303-3 ( frq 10 , his-3) был штаммом-хозяином для конструкции his-3 , нацеленной на конструкции frq . Различные штаммы sFRQ, которые экспрессируют sFRQ с внутренними делециями в рамке считывания, были созданы ранее (Liu et al. 2000; Cheng et al. 2005). Жидкие культуры выращивали в минимальной среде (1 × Vogel’s, 2% глюкоза).Для штаммов, несущих конструкции FRQ и Myc-CK-1a,
жидкие культуры выращивали в 0,01 M QA (pH 5,8), 1 × Vogel’s, 0,1% глюкозы и 0,17% аргинина. Среда гоночной трубки содержит
1 × Фогеля, 0,1% глюкозы, 0,17% аргинина, 50 нг / мл биотина и 1,5% агара. Денситометрические анализы гоночных трубок были
выполняется с использованием NIH Image.
Создание мутантных штаммов frq для изучения взаимодействия FRQ / CK-1a
pKAJ120 (содержащий весь ген frq , включая его промотор и целевую последовательность his-3 ) является родительской плазмидой для всех конструкций frq , описанных в этом исследовании.Все делеционные и точечные мутации ORF FRQ были сконструированы с использованием либо
набор для сайт-направленного мутагенеза трансформера (Clontech Laboratories) или набор для сайт-направленного мутагенеза QuikChange (Strata-
ген), а pUC19Mfrq использовали в качестве мутагенной матрицы in vitro. Впоследствии мутации были субклонированы в pKAJ120, и
Полученные конструкции трансформировали в frq 10 , his-3 в локусе his-3 с помощью электропорации (Margolin et al.1999). Для изучения взаимодействия FRQ / CK-1a конструкцию Myc-CK-1a вводили в трансформанты различных мутантов FRQ с помощью
котрансформация с помощью pBT6 (содержащего ген устойчивости к беномилу, полученный от Fungal Genetic Stock Center). Myc-CK-1a
конструкцию получали путем вставки фрагмента ПЦР, который содержит ORF для CK-1a и ее 3’UTR, в вектор pqa.5Myc (He et al. 2003).
Создание штамма ku70
RIP
Ген Neurospora ku70 (Ninomiya et al.2004) была нарушена точечной мутацией, индуцированной повтором (RIP) (Cambareri et al. 1989). Фрагмент ПЦР, содержащий всю ORF ku70, и ее 3’UTR, клонировали в pDE3BH и вводили в локус his-3 штамма дикого типа ( bd his-3 A ) путем электропорации. После идентификации положительных трансформантов их скрещивали с другим штаммом дикого типа.
( бд а ). Саузерн-блот-анализ использовали для идентификации потомков с мутированным геном ku70 .После этого было выполнено секвенирование ДНК эндогенного ku70 для идентификации штаммов, в которых эндогенная ORF ku70 была мутирована множеством преждевременных стоп-кодонов. Полученный штамм bd ku70 RIP использовали в качестве штамма-хозяина для введения блокирующей кассеты ck-1a .
Создание штаммов ck-1a
L
Для создания вставной кассеты с мутацией dbt L ген ck-1a дикого типа был клонирован в pDE3BH, и полученная плазмида pCKI-2 была использована в качестве матрицы для in vitro muta — генезис мутировать
CK-1a Met83 на Иль.После этого метод лигирования-ПЦР был использован для создания кассеты для детонации с использованием мутантного гена ck-1a и pCN44 (содержащего ген устойчивости hph ) в качестве матриц ПЦР (Zhao et al. 2004). В полученном фрагменте ПЦР с нокаутом ген hph вставлен ниже 3’UTR ck-1a и фланкирован всем геном ck-1a (с мутацией) и геномной ДНК 0,5 т.п. ck-1a (см. Рис. 3A). Затем кассету трансформировали в штамм bd ku70 RIP для отбора трансформантов, устойчивых к hph .Геномную ДНК из трансформантов получали, и ПЦР использовали для идентификации трансформантов.
с hph , интегрированным в эндогенный локус ck-1a . Продукты ПЦР, содержащие эндогенный ck-1a , секвенировали для идентификации штаммов гетерокарионов с нокаутной мутацией. После этого гомокариотические штаммы ck-1a L были получены путем очистки микроконидий (Ebbole and Sachs, 1990) и подтверждены анализом саузерн-блоттинга и секвенированием ДНК.
Анализ белков и РНК
Экстракция белка, количественная оценка, вестерн-блоттинг и иммунопреципитация выполнялись, как описано ранее.
(Гарсо и др., 1997; Ченг и др., 2001а). Равные количества общего белка (40 мкг) загружали в каждую белковую дорожку SDS-PAGE, и после электрофореза белки
переносили на PVDF-мембрану и проводили вестерн-блоттинг.Для анализа профилей фосфорилирования
Использовали гели WC-1 и WC-2, 10% SDS-PAGE, содержащие акриламид / бисакриламид в соотношении 149: 1. В противном случае 7,5% SDS-PAGE
Использовали гели с соотношением акриламид / бисакриламид 37,5: 1.
Экстракцию
РНК и Нозерн-блоттинг выполняли, как описано ранее (Aronson et al. 1994). Равные количества тотальной РНК (20 мкг) загружали в агарозные гели для электрофореза, и гели подвергали блоттингу и зондированию.
с зондом РНК, специфичным для frq.
ЧИП
Протокол анализа ChIP и используемые праймеры соответствуют описанным ранее (He and Liu 2005). Реакции ПЦР были следующими: 4 мин при 94 ° C, 26–29 циклов при 94 ° C (15 сек), 60 ° C (30 сек) и 72 ° C (1 мин). Разные
Количество циклов ПЦР проверяли, чтобы убедиться, что амплификация ДНК находится в пределах экспоненциального диапазона амплификации.Продукты ПЦР
были разделены электрофорезом на 2% агарозных гелях. Каждый эксперимент независимо проводился трижды, и иммунопреципитация
без антитела WC-2 использовали в качестве отрицательного контроля.
Благодарности
Мы благодарим Lixin Wang за отличную техническую помощь и Dr.Лили Ли за критическое прочтение и редактирование рукописи.
Это исследование было поддержано грантами от Национальных институтов здравоохранения (GM062591 и GM068496) и Фонда Уэлча для YL.
Ю.Л. является стипендиатом Луизы В. Кан в области биомедицинских исследований в Юго-западном медицинском центре Техасского университета.
Сноски
↵3 Текущие адреса: Институт медицинской биотехнологии Китайской академии медицинских наук и Пекинский союзный медицинский колледж,
Пекин Китай 100050;№4 Отделение нефрологии Юго-Западного медицинского центра Техасского университета, Даллас, Техас 75390
№5 Автор, ответственный за переписку.
№5 ЭЛЕКТРОННАЯ ПОЧТА yi.liu {at} utsouthwestern.edu; ФАКС (214) 645-6049.
Дополнительные материалы доступны на http://www.genesdev.org.
Статья размещена на сайте http: // www.genesdev.org/cgi/doi/10.1101/gad.1463506.
- Поступила 27.06.2006 г.
- Принята к печати 27 июля 2006 г.
- Авторские права © 2006, Лаборатория Колд-Спринг-Харбор
.