Баришевская Ольга Николаевна |
 Важным открытием стало обнаружение в ткани мозга мышей, а также в клеточной культуре, содержащей эндотелиальные клетки, мРНК порообразующих белков клаудина-2 и -17, тогда как по классическому представлению в состав плотных контактов в ЦНС входят только барьерообразующие клаудин-1, -3 и -5. Однако работ, показывающих точную локализацию этих белков в ткани мозга и подтверждающих их локализацию именно в ГЭБ на сегодняшний момент нет, что делает исследования с использованием иммунофлуоресценции к данным белкам крайне актуальными. Огромное влияние на функционирование ГЭБ играют глюкокортикоиды. Многочисленные клинические наблюдения и научные исследования говорят о снижении проницаемости ГЭБ под действием глюкокортикоидов. Не смотря на это, работ, раскрывающих молекулярные механизмы, лежащие в основе этого эффекта, на сегодняшний день крайне мало. На изучение этого вопроса и была направлена данная работа.
Важным открытием стало обнаружение в ткани мозга мышей, а также в клеточной культуре, содержащей эндотелиальные клетки, мРНК порообразующих белков клаудина-2 и -17, тогда как по классическому представлению в состав плотных контактов в ЦНС входят только барьерообразующие клаудин-1, -3 и -5. Однако работ, показывающих точную локализацию этих белков в ткани мозга и подтверждающих их локализацию именно в ГЭБ на сегодняшний момент нет, что делает исследования с использованием иммунофлуоресценции к данным белкам крайне актуальными. Огромное влияние на функционирование ГЭБ играют глюкокортикоиды. Многочисленные клинические наблюдения и научные исследования говорят о снижении проницаемости ГЭБ под действием глюкокортикоидов. Не смотря на это, работ, раскрывающих молекулярные механизмы, лежащие в основе этого эффекта, на сегодняшний день крайне мало. На изучение этого вопроса и была направлена данная работа. После хронического введения преднизолона снизилась масса тела и количество альдостерона у опытной крупы по сравнению с контрольной. Вестерн-блот и иммунофлуоресценция показали снижение уровня барьерообразующих клаудина-1, -3, и порообразующих клаудина-2 и -17. Количество клаудина-5, -12 и окклюдина не изменилось. Также с помощью иммунофлуоресценции была показана локализация клаудина-1, -2, -3, -5, 12 и 17 в эндотелии сосудов головного мозга мышей.
После хронического введения преднизолона снизилась масса тела и количество альдостерона у опытной крупы по сравнению с контрольной. Вестерн-блот и иммунофлуоресценция показали снижение уровня барьерообразующих клаудина-1, -3, и порообразующих клаудина-2 и -17. Количество клаудина-5, -12 и окклюдина не изменилось. Также с помощью иммунофлуоресценции была показана локализация клаудина-1, -2, -3, -5, 12 и 17 в эндотелии сосудов головного мозга мышей.Обходя гематоэнцефалический барьер
Все большее количество заболеваний ученые объясняют нарушением функций гематоэнцефалического барьера (ГЭБ). Его патологическая проницаемость развивается практически при всех видах патологии ЦНС. С другой же стороны, для обеспечения проникновения некоторых препаратов в мозг преодоление ГЭБ становится первоочередной задачей. Методики, позволяющие целенаправленно преодолевать защитный барьер между кровеносным руслом и мозговыми структурами, могут дать существенный толчок в терапии множества заболеваний.
С другой же стороны, для обеспечения проникновения некоторых препаратов в мозг преодоление ГЭБ становится первоочередной задачей. Методики, позволяющие целенаправленно преодолевать защитный барьер между кровеносным руслом и мозговыми структурами, могут дать существенный толчок в терапии множества заболеваний.
В одном из своих знаменитых экспериментов с красителями ныне всем известный ученый Пауль Эрлих обнаружил в конце XIX века интересный феномен, который занимает умы ученых и по сей день: после введения в кровь подопытной мыши органического красителя, наблюдая в микроскоп клетки различных органов, в том числе и клетки, относящиеся к органам центральной нервной системы, Эрлих отметил, что краситель проник во все ткани, за исключением головного мозга. После того, как помощник ученого сделал инъекцию красителя непосредственно в мозг, наблюдаемая в микроскоп картина была прямо противоположна: вещество мозга было окрашено красителем в темный фиолетово-синий цвет, тогда как в клетках других органов красителя обнаружено не было. Из своих наблюдений Эрлих заключил, что между мозгом и системным кровотоком должен существовать некий барьер.
Из своих наблюдений Эрлих заключил, что между мозгом и системным кровотоком должен существовать некий барьер.
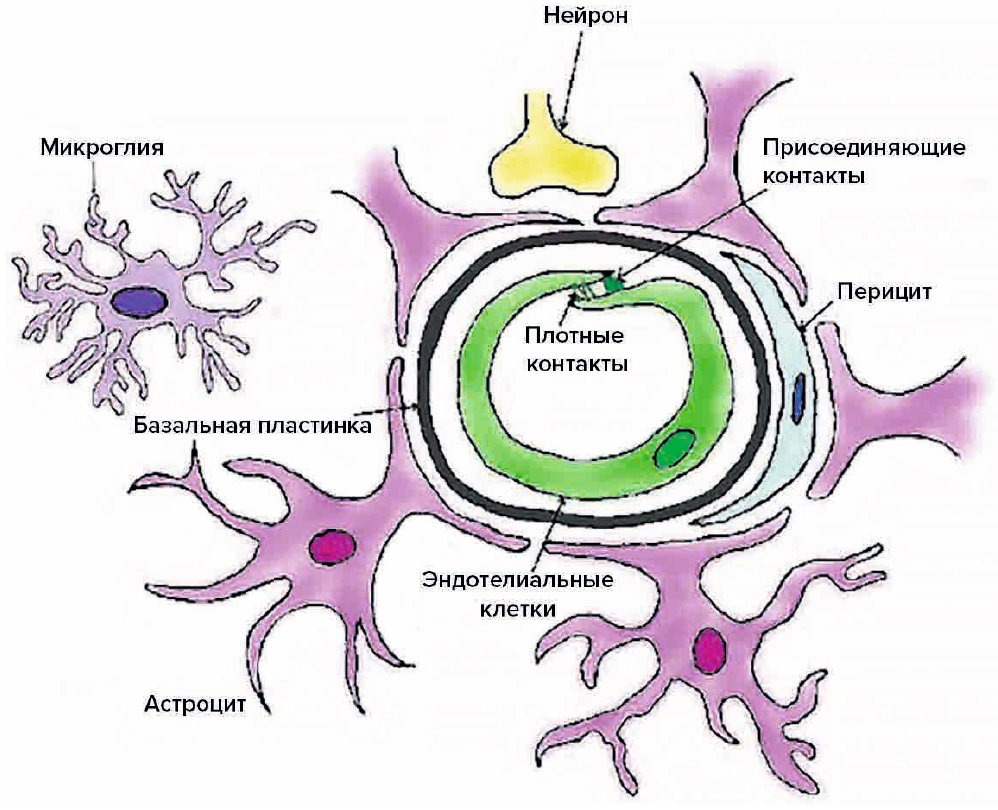
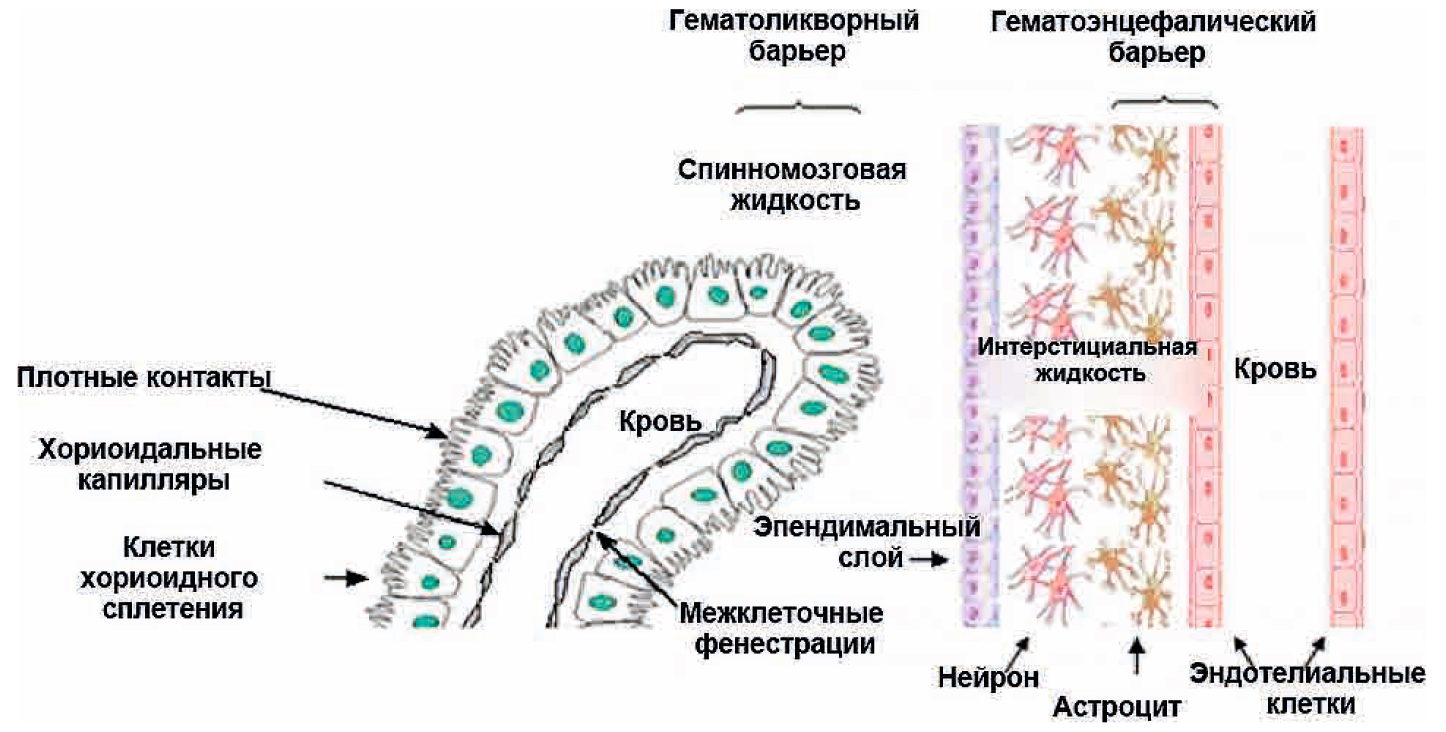
Через полвека после открытия Пауля Эрлиха, с появлением более мощных микроскопов, позволяющих наблюдать объекты с увеличением в 5000 раз большим, чем позволял микроскоп, используемый Эрлихом, удалось действительно идентифицировать гематоэнцефалический барьер. Он кроется в стенках многокилометровой сети кровеносных сосудов, снабжающей каждую из сотни миллиардов нервных клеток человеческого мозга. Как и все кровеносные сосуды, сосуды головного мозга выстланы изнутри эндотелиальными клетками. Однако эндотелиоциты, входящие в состав нейроваскулярной единицы головного мозга, прилегают друг к другу плотнее, чем на протяжении остального сосудистого русла. Межклеточные контакты между ними получили название „tight junctions“ (плотные контакты). Возможность формирования компактного нефенестрированного монослоя, экспрессия высокоспециализированных транспортных молекул и белков клеточной адгезии позволяют эндотелиоцитам поддерживать низкий уровень трансцитоза. Также эндотелий находится под действием регуляции со стороны перицитов, астроцитов, нейронов и молекул внеклеточного матрикса, что дает понять, что ГЭБ – это не просто слой эндотелиоцитов, а активный орган, включающий в себя разные типы клеток. Такое взаимодействие клеток, обеспечивающее барьерную функцию, препятствуя свободному перемещению жидкостей, макромолекул, ионов, объясняет, почему же ни краситель Пауля Эрлиха, ни некоторые лекарственные препараты не могут проникнуть из крови в ткани мозга.
Также эндотелий находится под действием регуляции со стороны перицитов, астроцитов, нейронов и молекул внеклеточного матрикса, что дает понять, что ГЭБ – это не просто слой эндотелиоцитов, а активный орган, включающий в себя разные типы клеток. Такое взаимодействие клеток, обеспечивающее барьерную функцию, препятствуя свободному перемещению жидкостей, макромолекул, ионов, объясняет, почему же ни краситель Пауля Эрлиха, ни некоторые лекарственные препараты не могут проникнуть из крови в ткани мозга.
Еще до того, как наличие ГЭБ стало явственным, врачи и ученые осознавали его значение. И вмешиваться в функционирование этого барьера считалось плохой идеей. Со временем это представление менялось, поскольку ГЭБ оказался высокоактивной структурой. Клетки с обеих сторон барьера находятся в постоянном контакте, оказывая взаимное влияние друг на друга. Разнообразные внутриклеточные молекулярные сигнальные пути определяют пропускную способность ГЭБ по отношению к разного типа молекулам (тут хотелось бы вспомнить сигнальный путь Wnt, координирующий множество процессов, связанных с дифференцировкой клеток, участвующий и в поддержании целостности ГЭБ). Лейкоциты, например, долгое время считавшиеся слишком крупными клетками для проникновения через ГЭБ, на самом деле преодолевают его, осуществляя «иммунологический надзор». Микроскопическая техника и сами микроскопы и сейчас не останавливаются в развитии, постоянно усложняются и открывают все больше возможностей для визуализации тонко устроенных структур живого организма. Например, использование двухфотонного микроскопа позволяет наблюдать живую ткань коры головного мозга на глубине около 300 мкм, что и было осуществлено доктором медицинских наук Майкен Недергаард из университета Рочестера. Ею были проведены следующие манипуляции: часть черепа мыши была удалена, затем была произведена инъекция красителя в кровеносное русло, что и позволило наблюдать ГЭБ в действии в реальном времени. Исследовательнице удалось отследить, как отдельные клетки перемещались из кровотока через стенку капилляров – через тот самый слой эндотелиальных клеток, который еще буквально 20 лет назад считался для них непроницаемым.
Лейкоциты, например, долгое время считавшиеся слишком крупными клетками для проникновения через ГЭБ, на самом деле преодолевают его, осуществляя «иммунологический надзор». Микроскопическая техника и сами микроскопы и сейчас не останавливаются в развитии, постоянно усложняются и открывают все больше возможностей для визуализации тонко устроенных структур живого организма. Например, использование двухфотонного микроскопа позволяет наблюдать живую ткань коры головного мозга на глубине около 300 мкм, что и было осуществлено доктором медицинских наук Майкен Недергаард из университета Рочестера. Ею были проведены следующие манипуляции: часть черепа мыши была удалена, затем была произведена инъекция красителя в кровеносное русло, что и позволило наблюдать ГЭБ в действии в реальном времени. Исследовательнице удалось отследить, как отдельные клетки перемещались из кровотока через стенку капилляров – через тот самый слой эндотелиальных клеток, который еще буквально 20 лет назад считался для них непроницаемым.
До того же, как был сконструирован двухфотонный микроскоп, исследователи пользовались классическими методами: например, наблюдали через микроскоп мертвые клетки ткани, что давало не много объяснений касательно функционирования ГЭБ. Ценно же наблюдение работы ГЭБ в динамике. В ряде экспериментов Недергаард и ее коллеги стимулировали определенную группу нервных клеток, с помощью чего была обнаружена невероятная динамичность ГЭБ: окружающие нейроны кровеносные сосуды расширялись при стимуляции нервных клеток, обеспечивая усиленный приток крови, поскольку стимулируемые нейроны начинали распространять потенциал действия; при снижении раздражающих импульсов сосуды сразу снова сужались. Также при оценке функций ГЭБ важно уделять внимание не только эндотелиоцитам, но и уже упомянутым астроцитам и перицитам, которые окружают сосуды и облегчают взаимодействие между кровью, эндотелием и нейронами. Не стоит недооценивать и циркулирующие вокруг клетки микроглии, поскольку дефекты их функций могут играть не последнюю роль в возникновении нейродегенеративных заболеваний, т. к. в этом случае ослабляется иммунная защита ГЭБ. При гибели эндотелиоцитов – по естественным причинам или вследствие повреждения – в гематоэнцефалическом барьере образуются «бреши», и эндотелиальные клетки не в состоянии сразу же закрыть данный участок, поскольку формирование плотных контактов требует времени. Значит, эндотелиоциты на этом участке должны быть временно замещены каким-то другим типом клеток. И именно клетки микроглии приходят на помощь, восстанавливая барьер, пока эндотелиальные клетки полностью не восстановятся. Это было показано в эксперименте командой доктора Недергаард, когда через 10-20 минут после повреждения капилляра головного мозга мыши лазерными лучами клетки микроглии заполнили повреждение. По этой причине, одна из гипотез, с помощью которой ученые пробуют объяснить возникновение нейродегенеративных заболеваний, – это нарушение функции микроглиальных клеток. Например, роль нарушений ГЭБ подтверждается в развитии атак рассеянного склероза: иммунные клетки в большом количестве мигрируют в ткани мозга, запуская синтез антител, атакующих миелин, вследствие чего разрушается миелиновая оболочка аксонов.
к. в этом случае ослабляется иммунная защита ГЭБ. При гибели эндотелиоцитов – по естественным причинам или вследствие повреждения – в гематоэнцефалическом барьере образуются «бреши», и эндотелиальные клетки не в состоянии сразу же закрыть данный участок, поскольку формирование плотных контактов требует времени. Значит, эндотелиоциты на этом участке должны быть временно замещены каким-то другим типом клеток. И именно клетки микроглии приходят на помощь, восстанавливая барьер, пока эндотелиальные клетки полностью не восстановятся. Это было показано в эксперименте командой доктора Недергаард, когда через 10-20 минут после повреждения капилляра головного мозга мыши лазерными лучами клетки микроглии заполнили повреждение. По этой причине, одна из гипотез, с помощью которой ученые пробуют объяснить возникновение нейродегенеративных заболеваний, – это нарушение функции микроглиальных клеток. Например, роль нарушений ГЭБ подтверждается в развитии атак рассеянного склероза: иммунные клетки в большом количестве мигрируют в ткани мозга, запуская синтез антител, атакующих миелин, вследствие чего разрушается миелиновая оболочка аксонов.
Патологическая проницаемость ГЭБ также играет роль в возникновении и течении эпилепсии. Уже достаточно давно известно, что эпилептические припадки связаны с преходящим нарушением целостности ГЭБ. Правда, до недавнего времени считалось, что это последствие приступов эпилепсии, а не причина. Но с получением новых результатов исследований эта точка зрения постепенно изменилась. Например, по данным лаборатории университета Амстердама, частота припадков у крыс повышалась соответственно открытию ГЭБ. Чем более выраженным было нарушение барьера, тем более вероятно у животных развивалась височная форма эпилепсии. С этими данными коррелируют также результаты, полученные в Кливлендской клинике (США) при проведении испытаний на свиньях, а также на примере людей: в обоих случаях судорожные припадки происходили после открытия ГЭБ, но никогда — до этого.
Также ученые занимаются и взаимосвязью функционирования ГЭБ с болезнью Альцгеймера. К примеру, удалось идентифицировать два белка ГЭБ, которые, вероятно, играют роль в развитии данного заболевания. Один из этих белков – RAGE – опосредует проникновение молекул бета-амилоида из крови в ткань головного мозга, а другой – LRP1 – транспортирует их наружу. Если равновесие в деятельности этих белков нарушается, формируются характерные амилоидные бляшки. И хотя применение этих знаний для терапии еще только в будущем, есть дающие надежды результаты: на модели мышей удается предотвратить отложение бета-амилоида, заблокировав ген, ответственный за синтез RAGE-белков в эндотелиальных клетках. Возможно, препараты, блокирующие белок RAGE, работа над созданием которых уже ведется, будут иметь сходный эффект и у человека.
Один из этих белков – RAGE – опосредует проникновение молекул бета-амилоида из крови в ткань головного мозга, а другой – LRP1 – транспортирует их наружу. Если равновесие в деятельности этих белков нарушается, формируются характерные амилоидные бляшки. И хотя применение этих знаний для терапии еще только в будущем, есть дающие надежды результаты: на модели мышей удается предотвратить отложение бета-амилоида, заблокировав ген, ответственный за синтез RAGE-белков в эндотелиальных клетках. Возможно, препараты, блокирующие белок RAGE, работа над созданием которых уже ведется, будут иметь сходный эффект и у человека.
Помимо проблемы восстановления целостности ГЭБ, другая проблема, связанная с его функционированием – это, как уже было сказано, переправление лекарственных препаратов через преграду между кровотоком и мозгом. Обмен веществ, осуществляемый через ГЭБ, подчиняется определенным правилам. Чтобы пересечь барьер, вещество должно либо по массе не превышать 500 кДа (этому параметру соответствует большинство антидепрессивных, антипсихотических и снотворных средств), либо использовать естественные механизмы для перехода ГЭБ, как это делает, например, L-дофа, представляющая собой предшественник дофамина и транспортируемая через ГЭБ специальным переносчиком; либо вещество должно быть липофильным, поскольку аффинитет к жиросодержащим соединениям обеспечивает прохождение через базальную мембрану. 98% препаратов не выполняют и по одному из этих трех критериев, а значит, не могут реализовать свой фармакологический эффект в мозге.
98% препаратов не выполняют и по одному из этих трех критериев, а значит, не могут реализовать свой фармакологический эффект в мозге.
Вышеперечисленные критерии технологи безуспешно пытаются реализовать в ходе разработки лекарственных форм. Хотя жирорастворимые формы легко проникают через ГЭБ, некоторые из них тут же снова выводятся обратно в кровоток, другие застревают в толще мембраны, не достигая конечной цели. Кроме того, липофильность не является избирательным свойством мембран ГЭБ, а потому такие препараты могут практически без разбора проходить через мембраны клеток любых органов организма, что тоже, безусловно, минус.
Способы преодоления гемато-энцефалического барьера
Настоящим прорывом стало использование хирургического метода преодоления ГЭБ, разработанного нейрохирургом из Техасского университета в Далласе. Метод заключается во введении гиперосмолярного раствора маннита в ведущую к мозгу артерию. За счет осмолярного воздействия (количество растворенного вещества в гиперосмолярном растворе маннита превышает таковое внутри эндотелиальных клеток, поэтому, по закону осмоса, вода перемещается в сторону большей концентрации растворенного вещества) эндотелиоциты теряют воду, сморщиваются, плотные контакты между ними разрываются, и образуется временный дефект в ГЭБ, позволяющий вводимым в ту же артерию препаратам проходить в ткань мозга. Такое временное открытие ГЭБ длится от 40 минут до 2-х часов, после чего происходит восстановление эндотелиоцитов и контактов между ними. Такая методика оказывается спасительной для пациентов с диагностированными опухолями головного мозга, когда опухоль хорошо откликается на химиотерапию, но только в том случае, пока химиотерапевтический препарат достигает ткани мозга и накапливается в зоне инфильтрации злокачественных клеток в необходимой концентрации.
Такое временное открытие ГЭБ длится от 40 минут до 2-х часов, после чего происходит восстановление эндотелиоцитов и контактов между ними. Такая методика оказывается спасительной для пациентов с диагностированными опухолями головного мозга, когда опухоль хорошо откликается на химиотерапию, но только в том случае, пока химиотерапевтический препарат достигает ткани мозга и накапливается в зоне инфильтрации злокачественных клеток в необходимой концентрации.
Это только один из способов преодоления ГЭБ. Существуют не менее интересные способы, они обзорно представлены на схеме внизу. Надеюсь, ознакомившись с ними, кто-то захочет углубиться в тему, чтобы разобраться в возможностях манипуляций с гематоэнцефалическим барьером и тем, как именно контроль над его функционированием может помочь в борьбе с различными заболеваниями.
Источники:
- Engaging neuroscience to advance translational research in brain barrier biology – полный текст статьи, выдержки из которой использовались в посте, об участии ГЭБ в развитии различных заболеваний и способы его преодоления
- J.
 Interlandi Wege durch die Blut-Hirn-Schranke, Spektrum der Wissenschaft, spezielle Auflage, 2/2016
Interlandi Wege durch die Blut-Hirn-Schranke, Spektrum der Wissenschaft, spezielle Auflage, 2/2016 - Blood-Brain Barrier Opening — обзор способов открытия ГЭБ
- Эндотелиальные прогениторные клетки в развитии и восстановлении церебрального эндотелия — о формировании и моделировании ГЭБ
 Interlandi Wege durch die Blut-Hirn-Schranke, Spektrum der Wissenschaft, spezielle Auflage, 2/2016
Interlandi Wege durch die Blut-Hirn-Schranke, Spektrum der Wissenschaft, spezielle Auflage, 2/2016
Церебролизин. Фармакологическое действие
Автор: Систряк З. С., ветеринарный врач, руководитель сообщества «Борьба с лженаукой в ветеринарии».
Состав: 1 мл водного раствора препарата содержит 215,2 мг концентрата церебролизина (комплекс пептидов, полученных из головного мозга свиньи). Активная фракция церебролизина представлена пептидами, молекулярный вес которых не превышает 10 000 дальтон. Вспомогательные вещества – натрия гидроксид и вода для инъекций.
Фармакотерапевтическая группа: ноотропное средство.
Фармакологическое действие
Фармакодинамика: церебролизин содержит низкомолекулярные биологически активные нейропептиды, которые проникают через гематоэнцефалический барьер и непосредственно поступают к нервным клеткам. Препарат обладает органоспецифическим мультимодальным действием на головной мозг, т. е. обеспечивает метаболическую регуляцию, нейропротекцию, функциональную нейромодуляцию и нейротрофическую активность.
Препарат обладает органоспецифическим мультимодальным действием на головной мозг, т. е. обеспечивает метаболическую регуляцию, нейропротекцию, функциональную нейромодуляцию и нейротрофическую активность.
а) метаболическая регуляция: церебролизин повышает эффективность аэробного энергетического метаболизма головного мозга, улучшает внутриклеточный синтез белка в развивающемся и стареющем головном мозге.
б) нейропротекция: церебролизин защищает нейроны от повреждающего действия лактацидоза, предотвращает образование свободных радикалов, повышает выживаемость и предотвращает гибель нейронов в условиях гипоксии и ишемии, снижает повреждающее нейротоксическое действие возбуждающих аминокислот (глутамата).
в) нейротрофическая активность: церебролизин – единственный пептидергический препарат с доказанной нейротрофической активностью, аналогичной действию естественных факторов нейронального роста (NGF), но проявляющейся в условиях периферического введения.
г) функциональная нейромодуляция: церебролизин оказывает положительное влияние при нарушениях когнитивных функций на процессы запоминания.
Фармакокинетика: сложный состав церебролизина, активная фракция которого состоит из сбалансированной и стабильной смеси биологически активных олигопептидов, обладающих суммарным полифункциональным действием, не позволяет провести обычный фармакокинетический анализ отдельных компонентов.
Показания к применению: болезнь Альцгеймера, синдром деменции различного генеза, хроническая цереброваскулярная недостаточность, ишемический инсульт, травматические повреждения головного и спинного мозга, задержка умственного развития у детей, гиперактивность и дефицит внимания у детей. В комплексной терапии используется при эндогенной депрессии, резистентной к антидепрессантам.
Разберемся
Сам препарат имеет долгую историю, он производится и изучается с начала 1970-х годов, является главным продуктом австрийской фармацевтической компании EVER Neuro Pharma. В городе Йена (Германия) находится завод компании, который (по словам представителей компании) считается признанным партнером в отрасли фармацевтического производства, поскольку его сотрудники имеют многолетний опыт в такой сложной отрасли, как разработка и производство стерильных лекарственных форм. Международное название у препарата «Церебролизин» отсутствует.
В городе Йена (Германия) находится завод компании, который (по словам представителей компании) считается признанным партнером в отрасли фармацевтического производства, поскольку его сотрудники имеют многолетний опыт в такой сложной отрасли, как разработка и производство стерильных лекарственных форм. Международное название у препарата «Церебролизин» отсутствует.
Существует официальный сайт препарата «Церебролизин», где есть такая информация о его составе: «Препарат имеет постоянный качественный и количественный состав. Это означает, что, несмотря на сложность и многокомпонентность состава, содержание основных веществ в церебролизине стандартизировано не только качественно, но и количественно. Раствор церебролизина содержит сбалансированное соотношение низкомолекулярных пептидов (25 %), свободных аминокислот (75 %), витаминов и микроэлементов. Именно такое соотношение пептидов к аминокислотам является оптимальным и обеспечивает высокую устойчивость раствора церебролизина, так как молекулы аминокислот образуют водородные связи с молекулами пептидов и обеспечивают их пространственную стабильность. В 1 мл раствора препарата содержится 215,2 мг концентрата церебролизина. Активная фракция препарата представлена пептидами, молекулярный вес которых не превышает 10 000 дальтон». Там же размещена таблица, где перечислены аминокислоты, которые содержатся в составе препарата.
В 1 мл раствора препарата содержится 215,2 мг концентрата церебролизина. Активная фракция препарата представлена пептидами, молекулярный вес которых не превышает 10 000 дальтон». Там же размещена таблица, где перечислены аминокислоты, которые содержатся в составе препарата.
Далее сказано следующее: «Церебролизин – продукт строго контролируемого биотехнологического процесса. Он производится методом ферментативного протеолитического гидролиза из очищенной белковой фракции, полученной из головного мозга молодых свиней».
Следует обратить внимание еще раз на тот момент, что в состав препарата, по заявлению разработчика, входят пептиды и аминокислоты. Пептиды – это вещества, состоящие из двух и более остатков аминокислот, соединенных между собой пептидными (амидными) связями. Пептиды, содержащие до 10 аминокислотных остатков, называются олигопептидами, а содержащие более 10 аминокислотных остатков – полипептидами. Полипептиды с молекулярной массой более 6 тысяч дальтон называются белками.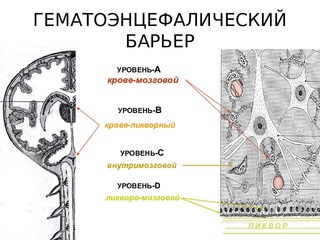 Пептиды отличаются от белков по размеру, и в качестве произвольного эталона можно понимать, что они содержит приблизительно 50 или менее аминокислот. Пептидов существует очень много: антимикробные пептиды, пептиды тахикинина, вазоактивные кишечные пептиды, опиоидные пептиды и т. д. На сайте разработчика есть такое упоминание, что в составе присутствуют не просто пептиды, а нейропептиды (то есть некое уточнение относительно пептидов все-таки есть). Нейропептиды – это также разновидность молекул белка, они образуются в центральной или периферической нервной системе и регулируют физиологические функции организма человека и животных. По своей функции, месту синтеза и структуре все нейропептиды, включая медиаторы и гормоны, подразделяются на 18 семейств. В некоторых из этих семейств имеется по 20–30 различных нейропептидов. Эти вещества отнесены к нейропептидам потому, что все они образуются определенными нейронами головного мозга или (как эндорфины) в гипофизе и, выступая затем в качестве нейромодуляторов, оказывают то или иное нейротропное или даже психотропное действие (например, эндорфины, вазопрессин, окситоцин, соматостатин).
Пептиды отличаются от белков по размеру, и в качестве произвольного эталона можно понимать, что они содержит приблизительно 50 или менее аминокислот. Пептидов существует очень много: антимикробные пептиды, пептиды тахикинина, вазоактивные кишечные пептиды, опиоидные пептиды и т. д. На сайте разработчика есть такое упоминание, что в составе присутствуют не просто пептиды, а нейропептиды (то есть некое уточнение относительно пептидов все-таки есть). Нейропептиды – это также разновидность молекул белка, они образуются в центральной или периферической нервной системе и регулируют физиологические функции организма человека и животных. По своей функции, месту синтеза и структуре все нейропептиды, включая медиаторы и гормоны, подразделяются на 18 семейств. В некоторых из этих семейств имеется по 20–30 различных нейропептидов. Эти вещества отнесены к нейропептидам потому, что все они образуются определенными нейронами головного мозга или (как эндорфины) в гипофизе и, выступая затем в качестве нейромодуляторов, оказывают то или иное нейротропное или даже психотропное действие (например, эндорфины, вазопрессин, окситоцин, соматостатин).
То есть, по логике производителя, получается, что нейропептиды сами должны складываться в различные пептиды из аминокислот.
Что конкретно входит в эти 25 % нейропептидов церебролизина, неизвестно и, собственно, вполне объяснимо. Ведь, как мы понимаем, если указать конкретные нейропептиды, то разработчику надо будет описывать все возможные эффекты для конкретно указанного нейропептида, его функции.
Если мы еще раз вернемся к инструкции, то обнаружим, что фармакокинетики там нет, фармакодинамика тоже не описана, зато механизм действия препарата представлен так: «Нейропептиды Церебролизина по своей структуре и фармакологическим свойствам оказались аналогичны действию естественных нейротрофических факторов, или нейротрофинов (НТФ) — сигнальных нейропептидов, которые в естественных условиях регулируют функции нервной ткани. Механизм действия Церебролизина аналогичен естественным НТФ и основан на так называемом «сигнальном пути»: активация мембранных тирозин-киназных рецепторов, программируемая экспрессия генов, внутриклеточный белковый синтез, активация и поддержка эндогенной системы защиты и регенерации клетки»
Подобный механизм действия не имеет никакого отношения к фармакологии, это описание больше похоже на размышления. Нам не объясняется, на какие мишени воздействует препарат и каким образом. Такие формулировки к истинным лекарственным средствам не относятся.
Нам не объясняется, на какие мишени воздействует препарат и каким образом. Такие формулировки к истинным лекарственным средствам не относятся.
Давайте на примере нейропептида вазопрессина рассмотрим механизм действия, чтобы наглядно понимать ситуацию. Вазопрессин, или антидиуретический гормон (АДГ) – пептидный гормон гипоталамуса, найденный у большинства млекопитающих. Гормон накапливается в задней доле гипофиза (в нейрогипофизе) и оттуда секретируется в кровь. Он состоит из 9 аминокислот: Cys-Tyr-Phe-Gln-Asn-Cys-Pro-(Arg или Lys)-Gly. У большинства млекопитающих в позиции 8 находится аргинин (аргинин-вазопрессин, AVP), у свиней и некоторых родственных животных – лизин (лизин-вазопрессин, LVP). Между остатками Cys1 и Cys6 формируется дисульфидная связь. Большая часть гормона синтезируется крупноклеточными нейронами супраоптического ядра гипоталамуса, аксоны которых направляются в заднюю долю гипофиза (нейрогипофиз) и образуют синаптоподобные контакты с кровеносными сосудами. Вазопрессин, синтезированный в телах нейронов, переносится аксонным транспортом к окончаниям аксонов, накапливается в пресинаптических везикулах и секретируется в кровь при возбуждении нейрона. Его связывание с V2-рецепторами собирательной трубки приводит к встраиванию в апикальную мембрану ее главных клеток белка водных каналов аквапорина 2, что увеличивает проницаемость эпителия собирательной трубки для воды и ведет к усилению ее реабсорбции. В отсутствие вазопрессина, например при несахарном диабете, суточный диурез у человека может достигать 20 л, тогда как в норме он составляет 1,5 литра. В экспериментах на изолированных почечных канальцах вазопрессин увеличивает реабсорбцию натрия, тогда как на целых животных вызывает увеличение экскреции этого катиона. Каким образом разрешить это противоречие, до настоящего времени неясно. Через V1a-рецепторы вазопрессин повышает тонус гладкой мускулатуры внутренних органов (в особенности ЖКТ), сосудистый тонус и таким образом вызывает увеличение периферического сопротивления. Благодаря этому, а также за счет роста объема циркулирующей крови (ОЦК) вазопрессин повышает артериальное давление. Главным стимулом для секреции вазопрессина является повышение осмолярности плазмы крови, обнаруживаемое осморецепторами в самих паравентрикулярном и супраоптическом ядрах гипоталамуса, в области передней стенки третьего желудочка, а также, по-видимому, в печени и ряде других органов.
Его связывание с V2-рецепторами собирательной трубки приводит к встраиванию в апикальную мембрану ее главных клеток белка водных каналов аквапорина 2, что увеличивает проницаемость эпителия собирательной трубки для воды и ведет к усилению ее реабсорбции. В отсутствие вазопрессина, например при несахарном диабете, суточный диурез у человека может достигать 20 л, тогда как в норме он составляет 1,5 литра. В экспериментах на изолированных почечных канальцах вазопрессин увеличивает реабсорбцию натрия, тогда как на целых животных вызывает увеличение экскреции этого катиона. Каким образом разрешить это противоречие, до настоящего времени неясно. Через V1a-рецепторы вазопрессин повышает тонус гладкой мускулатуры внутренних органов (в особенности ЖКТ), сосудистый тонус и таким образом вызывает увеличение периферического сопротивления. Благодаря этому, а также за счет роста объема циркулирующей крови (ОЦК) вазопрессин повышает артериальное давление. Главным стимулом для секреции вазопрессина является повышение осмолярности плазмы крови, обнаруживаемое осморецепторами в самих паравентрикулярном и супраоптическом ядрах гипоталамуса, в области передней стенки третьего желудочка, а также, по-видимому, в печени и ряде других органов. Кроме того, секреция гормона повышается при уменьшении ОЦК, которое воспринимают волюморецепторы внутригрудных вен и предсердий.
Кроме того, секреция гормона повышается при уменьшении ОЦК, которое воспринимают волюморецепторы внутригрудных вен и предсердий.
В приведенном примере механизм действия понятен, все изучено и описано. У церебролизина конкретные нейропептиды не описаны, об их присутствии лишь абстрактно заявлено в инструкции. А если не описано, какие именно нейропептиды присутствуют, возникает вопрос, есть ли они там вообще? И если нет кинетики в инструкции, то почему производитель решил, что эти некие пептиды проникают через гематоэнцефалический барьер (ГЭБ)? Видимо (по логике производителя), только потому, что ГЭБ пропускает молекулы весом до 30 тысяч дальтон в норме.
Зато производитель четко и ясно описал аминокислоты, которые находятся в составе, даже описал сколько миллиграмм каждой аминокислоты содержится в 1 миллилитре раствора. А вот указанные в составе витамины и микроэлементы не описаны. Они там действительно есть? Какие именно? Их количество? Все дело в том, что аминокислоты проще всего стандартизировать, так как существуют определенные требования контроля качества при производстве препаратов и аминокислоты легко определить в составе, ведь они представляют собой низкомолекулярные соединения, а полипептиды – это полимеры, поэтому аминокислоты и указаны конкретно в инструкции.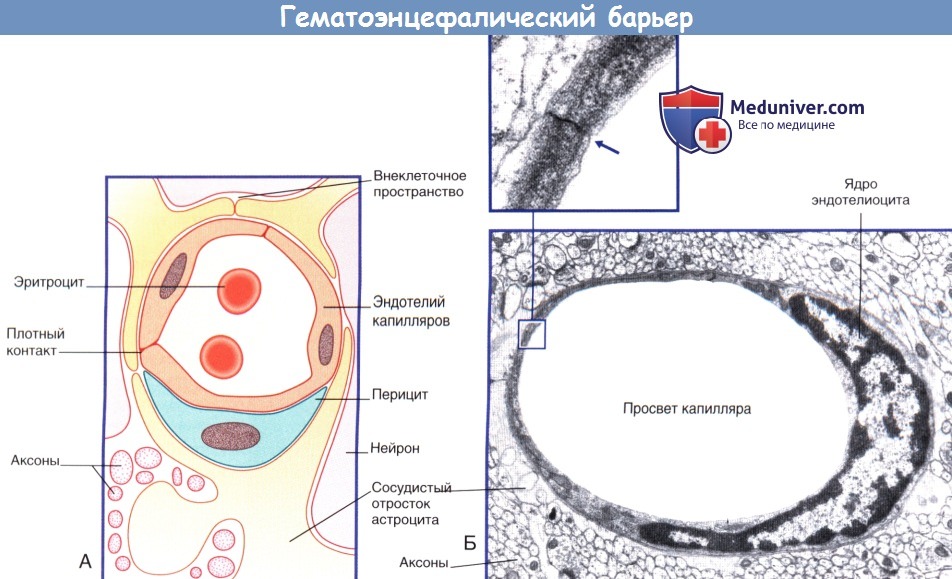 Но этот факт сам по себе ни о чем не говорит, просто вводит в заблуждение.
Но этот факт сам по себе ни о чем не говорит, просто вводит в заблуждение.
Следовательно, церебролизин – это набор аминокислот, а именуемый производителем «концентрат церебролизина» – это вводящее в заблуждение словосочетание, не имеющее под собой никакой основы. Так чем же тогда церебролизин отличается от парентерального питания?
Давайте для примера рассмотрим препарат «Аминовен». Мы увидим в составе тот же набор аминокислот, что и у церебролизина, только почему-то показания к применению отличаются: «Аминовен предназначен для парентерального питания больных с различными патологиями, с низкой, нормальной или повышенной потребностью в белке, когда энтеральное питание неэффективно или невозможно». Никаких упоминаний о лечении болезни Альцгеймера, ишемического инсульта, задержки умственного развития у детей, травматического повреждения спинного и головного мозга нет. Только содержание аминокислот аминовена указаны в граммах, а также четко обозначена дозировка 0,8–1 г/кг аминокислот в сутки. Дозы же церебролизина, согласно инструкции, в среднем на человека составляют 10–30 мл, а в 1 мл содержится от 0,21 до 7,20 мг аминокислот, это означает, что пациент получает 2,1–216 мг аминокислот в сутки. Несложно сделать вывод, что эти почти гомеопатические дозы не окажут на организм вообще никакого терапевтического влияния. То есть можно взять 1 мл аминовена, развести в 20 мл воды, и у нас получится не парентеральное питание, а средство от инсульта и лекарство от умственной недостаточности у детей.
Дозы же церебролизина, согласно инструкции, в среднем на человека составляют 10–30 мл, а в 1 мл содержится от 0,21 до 7,20 мг аминокислот, это означает, что пациент получает 2,1–216 мг аминокислот в сутки. Несложно сделать вывод, что эти почти гомеопатические дозы не окажут на организм вообще никакого терапевтического влияния. То есть можно взять 1 мл аминовена, развести в 20 мл воды, и у нас получится не парентеральное питание, а средство от инсульта и лекарство от умственной недостаточности у детей.
Далее мы сталкиваемся с другой ложной информацией от производителя. На официальном сайте написано следующее: «В США церебролизин имеет статус нового лекарственного препарата, разрешенного Администрацией по контролю за продуктами и лекарствами (Food and Drug Administration; FDA) для клинического применения в частном порядке». Если мы обратимся
на сайт FDA, то информации об этом, равно как и о церебролизине, не обнаружим.
А теперь обратимся к нашей науке, в частности к исследованиям.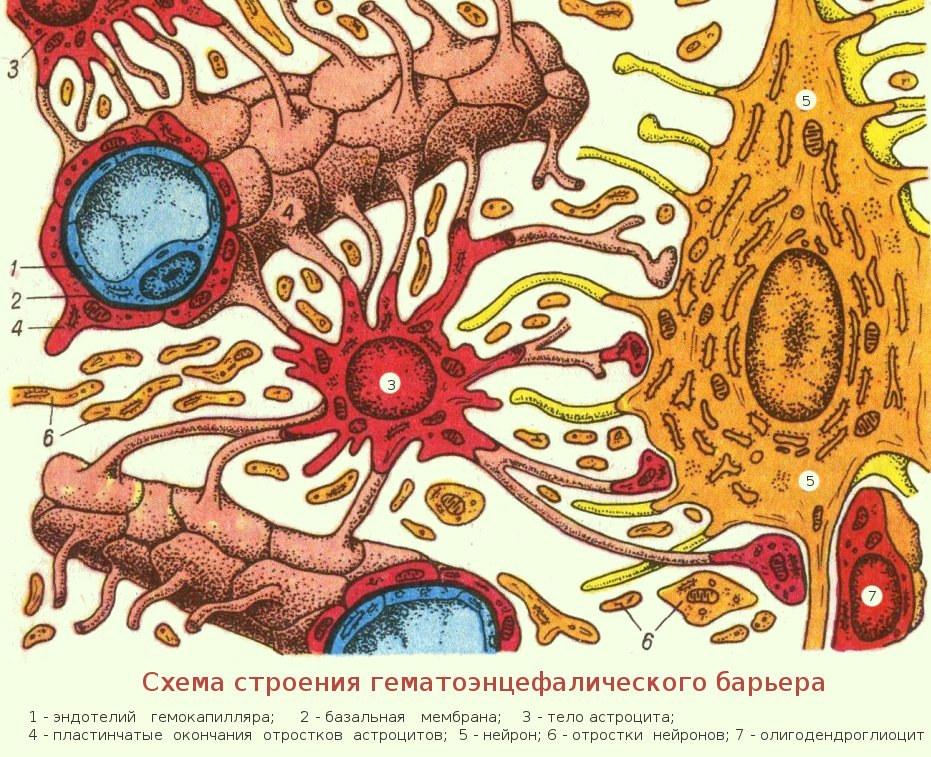 На самом деле, исследований церебролизина очень много, и, к счастью, мы имеем возможность сравнивать метаанализы этих исследований, которые публикует Кокрановское сотрудничество. Существует обзор 2010 года, в котором оценивали преимущества и риски церебролизина для лечения острого ишемического инсульта. Критерий выбора был четким – рандомизированные контролируемые исследования, сравнивающие церебролизин с плацебо, для выявления отсутствия лечения у пациентов с острым ишемическим инсультом. Выводы таковы, что имеется недостаточно доказательств для оценки влияния церебролизина на выживаемость и зависимость у людей с острым ишемическим инсультом (
На самом деле, исследований церебролизина очень много, и, к счастью, мы имеем возможность сравнивать метаанализы этих исследований, которые публикует Кокрановское сотрудничество. Существует обзор 2010 года, в котором оценивали преимущества и риски церебролизина для лечения острого ишемического инсульта. Критерий выбора был четким – рандомизированные контролируемые исследования, сравнивающие церебролизин с плацебо, для выявления отсутствия лечения у пациентов с острым ишемическим инсультом. Выводы таковы, что имеется недостаточно доказательств для оценки влияния церебролизина на выживаемость и зависимость у людей с острым ишемическим инсультом (
полная версию статьи).
А вот в обзор статьи 2017 года были включены 6 рандомизированных контролируемых исследований с общим числом участников – 1501, где сравнили церебролизин с плацебо (неактивное лекарство), которые использовались в дополнение к стандартной терапии, в том числе к тромболизису. Три из них были крупными многоцентровыми исследованиями, два – небольшими, и были признаны исследованиями неясного качества, а в одном не было числовых результатов. Исходя из основных результатов (доказательства актуальны по июнь 2016 год), этот обзор показал отсутствие пользы от применения церебролизина для снижения смертности при остром ишемическом инсульте. Не было разницы в общем числе людей с неблагоприятными событиями, но существуют опасения, что церебролизин может увеличить риск развития нефатальных серьезных неблагоприятных событий по сравнению с плацебо. Лекарство (церебролизин) и методология большинства включенных испытаний были предоставлены производителем церебролизина, создавая вероятный конфликт интересов. В настоящее время имеются доказательства среднего (умеренного) качества, позволяющие предполагать, что церебролизин работает не лучше, чем плацебо, в отношении смерти пациентов по любым причинам, когда он назначается людям с острым ишемическим инсультом в течение 48 часов после начала инсульта. Существуют доказательства среднего (умеренного) качества, которые вызывают обеспокоенность по поводу увеличения числа серьезных неблагоприятных событий при использовании церебролизина у людей с острым ишемическим инсультом.
Исходя из основных результатов (доказательства актуальны по июнь 2016 год), этот обзор показал отсутствие пользы от применения церебролизина для снижения смертности при остром ишемическом инсульте. Не было разницы в общем числе людей с неблагоприятными событиями, но существуют опасения, что церебролизин может увеличить риск развития нефатальных серьезных неблагоприятных событий по сравнению с плацебо. Лекарство (церебролизин) и методология большинства включенных испытаний были предоставлены производителем церебролизина, создавая вероятный конфликт интересов. В настоящее время имеются доказательства среднего (умеренного) качества, позволяющие предполагать, что церебролизин работает не лучше, чем плацебо, в отношении смерти пациентов по любым причинам, когда он назначается людям с острым ишемическим инсультом в течение 48 часов после начала инсульта. Существуют доказательства среднего (умеренного) качества, которые вызывают обеспокоенность по поводу увеличения числа серьезных неблагоприятных событий при использовании церебролизина у людей с острым ишемическим инсультом. Дальнейшие исследования, вероятно, окажут существенное влияние на нашу уверенность в оценке рисков серьезных неблагоприятных событий, связанных с применением церебролизина у людей с
Дальнейшие исследования, вероятно, окажут существенное влияние на нашу уверенность в оценке рисков серьезных неблагоприятных событий, связанных с применением церебролизина у людей с
острым инсультом.
Церебролизин в ветеринарии
Информации о применении препарата в ветеринарии практически нет, можно обнаружить лишь несколько статей, где описывают влияние церебролизина при когнитивных нарушениях у крыс, но эта информация не имеет никакого отношения к реальности (
например). Еще есть статья о том, как использовали церебролизин у собак для ускорения процесса восстановления
животных после операции. Данная информация также не имеет никакого отношения к науке, реальности, так как невозможно ни определить скорость восстановления пациента после операции, ни увеличить ее, поскольку единицы измерения данной скорости не существует, как и определенных цифр, показывающих, какой должна быть эта скорость и по сравнению с чем. Доз церебролизина для животных не существует.
Таким образом, препарат «Церебролизин» – это набор аминокислот в гомеопатических дозах. Действие «концентрата церебролизина» из неидентифицированных нейропептидов ничем не подтверждено, кроме слов из инструкции. Что это за пептиды – неизвестно. У препарата не описан механизм действия, не описаны также фармакокинетика и фармакодинамика. Информации о том, что набор неких нейропептидов проникает через ГЭБ и оказывает терапевтический эффект, нет. Сами аминокислоты, по сути, обладают лишь питательными функциями при их дефиците, а внутривенное введение аминокислот требуется только при невозможности принимать пищу внутрь. Терапевтической активностью аминокислоты не обладают, у них нет таких показаний к применению, как ишемический инсульт, задержка умственного развития у детей, ЧМТ и др. Современные научные обзоры подтверждают неэффективность церебролизина, а большинство из существующих статей о данном препарате предоставлены самим производителем, что создает конфликт интересов.
Особенности гипоталамической моноаминергической регуляции функции коры надпочечников у больных с гиперандрогенемией | Комаров
Аннотация
Поскольку большинство моноаминов не могут проникать через гематоэнцефалический барьер, трудно изучить их центральные метаболические нарушения. Тест с этимизолом был использован для изучения адренокортикальной функции у пациентов с гиперандрогенемией; этимизол может проникать через этот барьер, он стимулирует структуры мозга, которые регулируют адренокортикальную функцию посредством активации эндогенных моноаминов. У 10 пациенток с распространенной формой поликистоза яичников реакция АКТГ и гидрокортизона на введение этимизола была практически такой же, как и в контрольной группе. Этот ответ был увеличен в 13 пациентах с овариальным поликистозом и надпочечниковой гиперандрогенемией, и был купирован ципрогептадином. Этот факт свидетельствует о том, что надпочечниковый компонент гиперандрогенемии объясняется гиперактивностью гипоталамических серотонинергических систем, регулирующих секрецию кортикотропин-рилизинг-фактора. У 8 пациенток с поликистозом яичников и функциональной гиперпролактинемией реакция АКТГ и гидрокортизона на введение этимизола была снижена; такой тип ответа наблюдается при блокировании метоклопрамидом дофаминергических рецепторов.
Тест с этимизолом был использован для изучения адренокортикальной функции у пациентов с гиперандрогенемией; этимизол может проникать через этот барьер, он стимулирует структуры мозга, которые регулируют адренокортикальную функцию посредством активации эндогенных моноаминов. У 10 пациенток с распространенной формой поликистоза яичников реакция АКТГ и гидрокортизона на введение этимизола была практически такой же, как и в контрольной группе. Этот ответ был увеличен в 13 пациентах с овариальным поликистозом и надпочечниковой гиперандрогенемией, и был купирован ципрогептадином. Этот факт свидетельствует о том, что надпочечниковый компонент гиперандрогенемии объясняется гиперактивностью гипоталамических серотонинергических систем, регулирующих секрецию кортикотропин-рилизинг-фактора. У 8 пациенток с поликистозом яичников и функциональной гиперпролактинемией реакция АКТГ и гидрокортизона на введение этимизола была снижена; такой тип ответа наблюдается при блокировании метоклопрамидом дофаминергических рецепторов.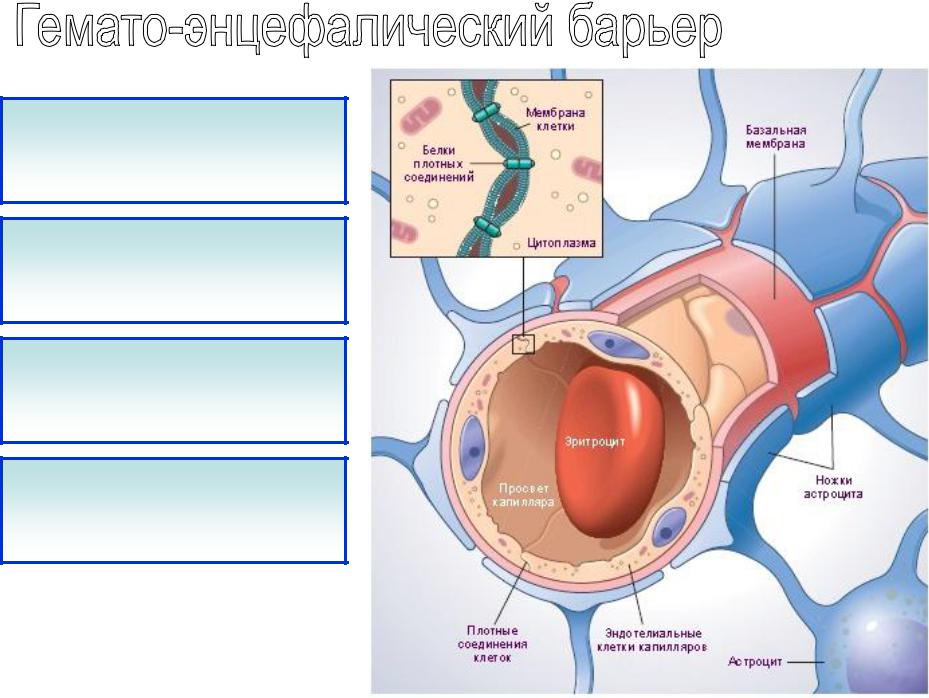 Этот факт указывает на наличие гипоталамической дофаминергической недостаточности у больных с поликистозом яичников и гиперпролактинемией, причем эта недостаточность затрагивает не только центры регуляции функции надпочечников, но, возможно, и структуры трансмиттерных систем гипоталамуса.
Этот факт указывает на наличие гипоталамической дофаминергической недостаточности у больных с поликистозом яичников и гиперпролактинемией, причем эта недостаточность затрагивает не только центры регуляции функции надпочечников, но, возможно, и структуры трансмиттерных систем гипоталамуса.
Повышенная секреция андрогенов надпочечниками является частой причиной гиперандрогене- мии у женщин и встречается приблизительно у 50 % больных с синдромом поликистозных яичников [9]. Патогенез этого нарушения остается недостаточно ясным. Одним из возможных механизмов предполагалось повышение функциональной активности коры надпочечников [10] вследствие нарушения центральной регуляции адренокортикальной функции [3]. Такое предположение согласуется с клиническими наблюдениями развития гирсутизма, аменореи и поликистозных изменений яичников в сочетании с типичными гормональными нарушениями после воздействия стрессорных факторов [1]. Одним из центральных механизмов, осуществляющих запуск стресс-реакции, является активность гипоталамической адренергической системы.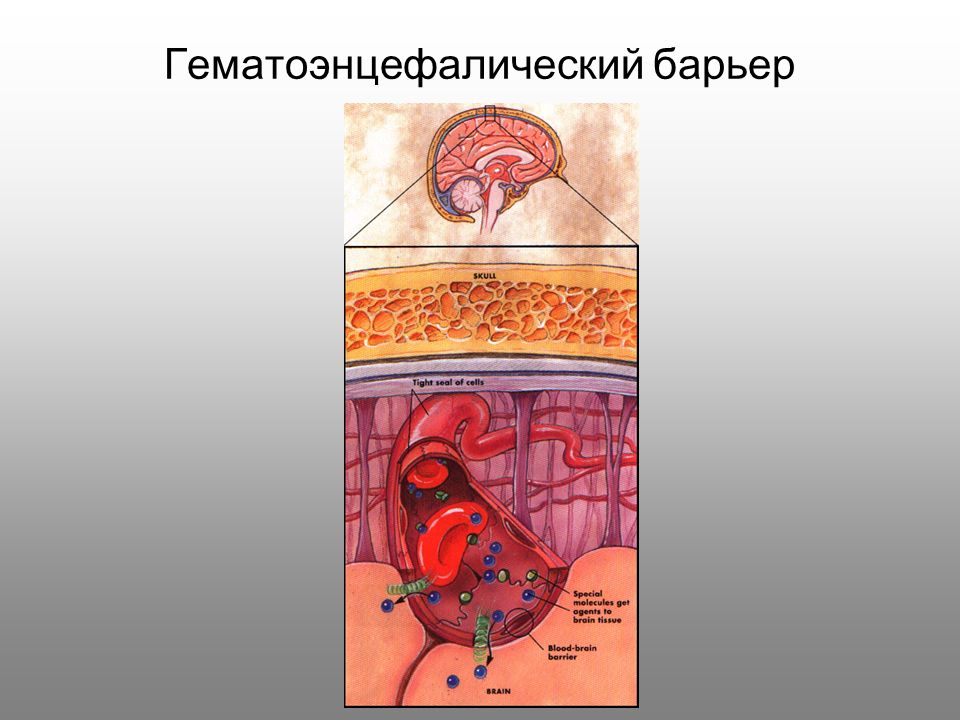 В ряде исследований приведены данные о нарушении моноаминергической регуляции у больных с поликистозом яичников. В частности, известны представления о недостаточной дофаминергической ингибиции гонадотропной функции гипофиза при поликистозе яичников [7, 14], о повышении серотониновой активности у больных с поликистозом яичников в сочетании с гиперфункцией надпочечников [4]. Особая трудность при изучении центральных нарушений обмена моноаминов состоит в том, что большинство из них не проникает через гематоэнцефалический барьер, поэтому содержание моноаминов в периферической циркуляции не отражает их активность в мозговых структурах. Отечественный препарат этими- зол, проникающий через гематоэнцефалический барьер, оказывает стимулирующее влияние на мозговые структуры, регулирующие функцию ги- поталамо-гипофизарно-надпочечниковой системы [5], путем активации эндогенных норадреналина, дофамина и серотонина [6]. Задачей настоящей работы явилось изучение гипоталамической моноаминергической регуляции функции коры надпочечников с помощью пробы с этимизолом у больных с гиперандрогенемией.
В ряде исследований приведены данные о нарушении моноаминергической регуляции у больных с поликистозом яичников. В частности, известны представления о недостаточной дофаминергической ингибиции гонадотропной функции гипофиза при поликистозе яичников [7, 14], о повышении серотониновой активности у больных с поликистозом яичников в сочетании с гиперфункцией надпочечников [4]. Особая трудность при изучении центральных нарушений обмена моноаминов состоит в том, что большинство из них не проникает через гематоэнцефалический барьер, поэтому содержание моноаминов в периферической циркуляции не отражает их активность в мозговых структурах. Отечественный препарат этими- зол, проникающий через гематоэнцефалический барьер, оказывает стимулирующее влияние на мозговые структуры, регулирующие функцию ги- поталамо-гипофизарно-надпочечниковой системы [5], путем активации эндогенных норадреналина, дофамина и серотонина [6]. Задачей настоящей работы явилось изучение гипоталамической моноаминергической регуляции функции коры надпочечников с помощью пробы с этимизолом у больных с гиперандрогенемией.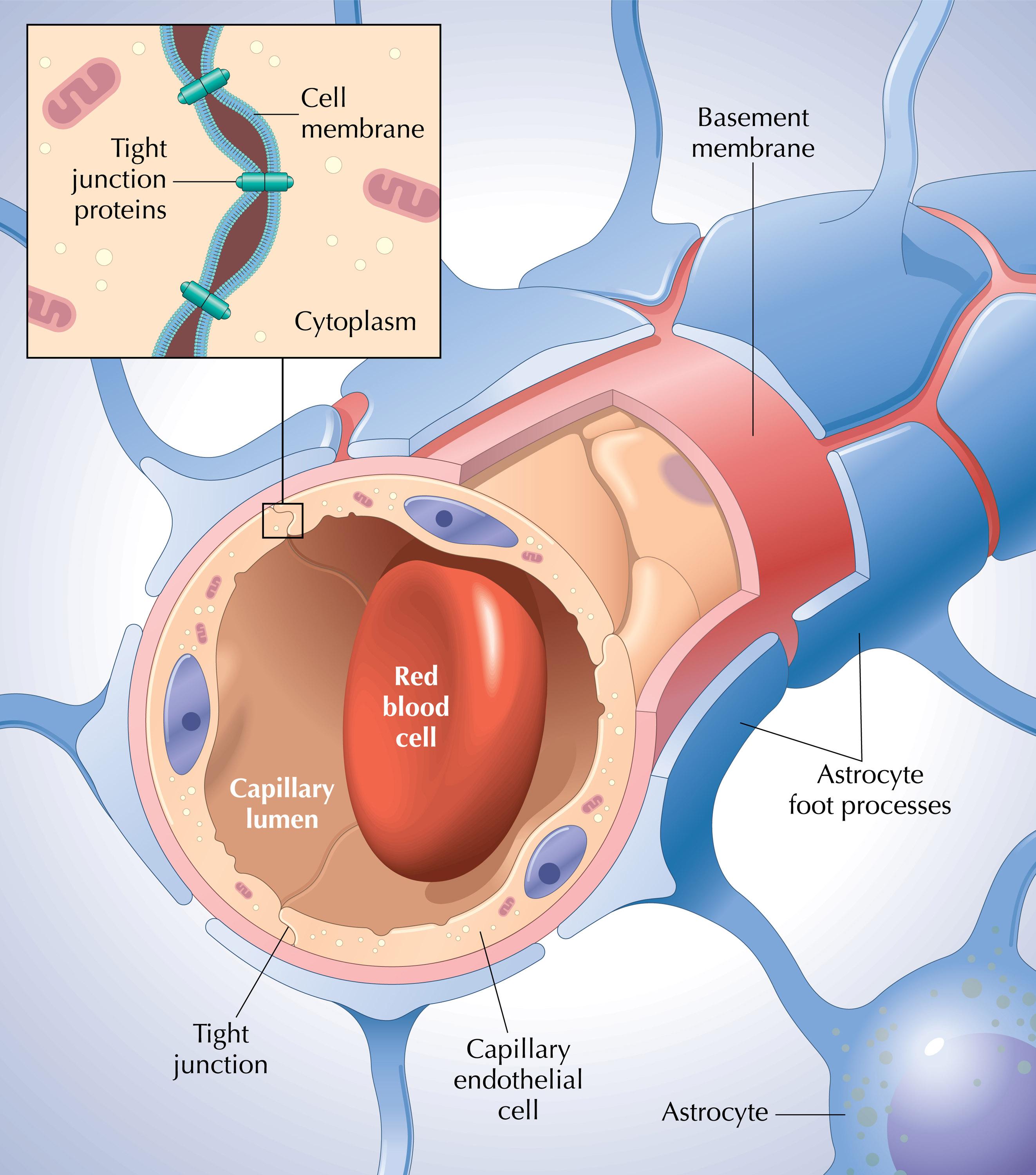
Материалы и методы
Обследована 31 больная в возрасте от 18 до 35 лет с гирсутизмом и различными нарушениями менструальной функции (аменорея — у 2 больных, олигоменорея — у 19, анову- ляция или недостаточность лютеиновой фазы цикла — у 10). Бесплодие наблюдалось у 11 женщин. У 6 больных имелось ожирение I—II степени. Контрольную группу составили 10 женщин в возрасте 18—37 лет без признаков гидрогенизации и нарушений менструальной и репродуктивной функций. Пробу с этимизолом проводили путем внутривенного введения 5 мл 1,5 % раствора непосредственно после взятия крови для гормональных исследований в базальных условиях. Повторное взятие крови производилось через 15, 30 и 90 мин после введения этимизола. Определяли содержание в крови пролактина (ПРЛ), кортикотропина (АКТЕ) и кортизола (К) радиоиммунологическим методом. Для выяснения роли отдельных моноаминергических систем у 23 больных проба с этимизолом была проведена повторно через 1 нед на фоне предварительного применения в течение 3 сут блокатора серотониновых рецепторов ципрогептадина (перитол) по 4 мг каждые 6 ч (7 больных), блокатора дофаминергических рецепторов метоклопрамида по 10 мг 4 раза в сутки (9 больных) и предшественника дофамина левопа по 250 мг каждые 6 ч (7 больных).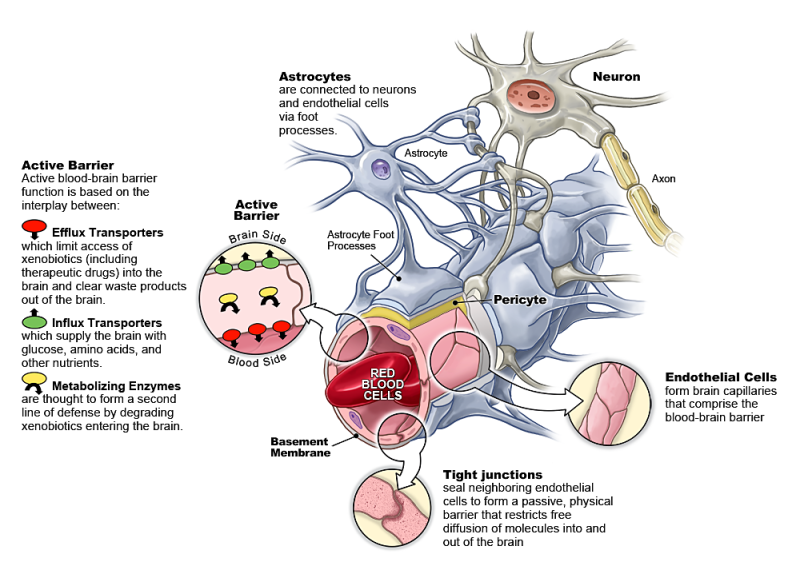 С интервалом не менее 1 нед от времени проведения пробы с этимизолом проводилась стандартная проба с 2 мг дексаметазона, во время которой исследовали содержание тестостерона (Т) в крови радиоиммунологическим методом. Всем больным проводилось ультразвуковое исследование матки и яичников. Полученные результаты обработаны статистически методом Стыодента.
С интервалом не менее 1 нед от времени проведения пробы с этимизолом проводилась стандартная проба с 2 мг дексаметазона, во время которой исследовали содержание тестостерона (Т) в крови радиоиммунологическим методом. Всем больным проводилось ультразвуковое исследование матки и яичников. Полученные результаты обработаны статистически методом Стыодента.
Результаты и их обсуждение
У 10 больных при исследовании установлена обычная форма синдрома поликистозных яичников (ПКЯ). У всех больных этой группы ультра-
Рис. 1. Динамика содержания К и АКТГ в крови (в % к исходному уровню) во время пробы с этимизолом у больных с обычной формой ПКЯ (/), ПКЯ с надпочечниковым компонентом гиперандрогенемйи (2), ПКЯ с гиперпролактинемией (3) и у здоровых женщин контрольной группы (заштрихованная область; М+т).
звуковым методом определялись увеличенные и поликистозно-измененные яичники. Экскреция 17-КС с мочой составляла 57,0+3,4 мкмоль/сут и достоверно превышала соответствующий показатель в контрольной группе (36,1 ±6,9 мкмоль/сут).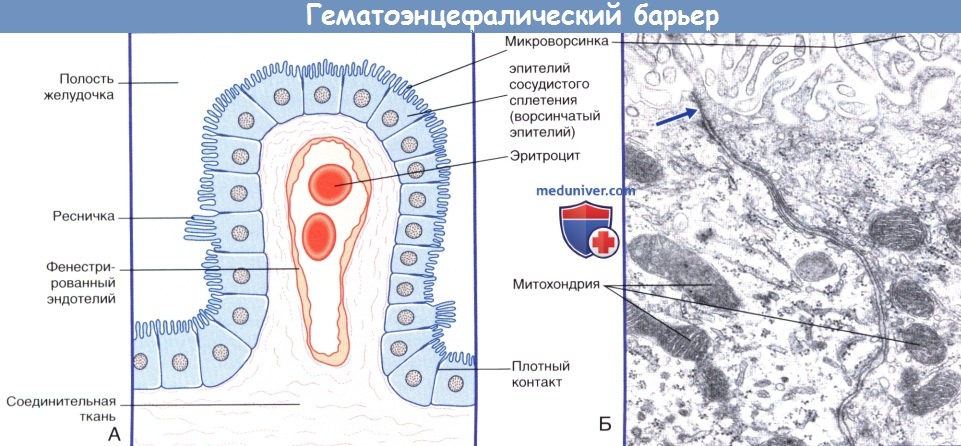 Содержание Т в крови 2,75±0,61 нмоль/л достоверно (р<0,02) превышало показатель в контрольной группе (1,42+0,15 нмоль/л), а снижение его при пробе с дексаметазоном достигало лишь 86,7+5,3 % от исходного уровня. Содержание ПРЛ в крови у больных этой группы составляло 269,5+49,5 мМЕ/л и соответствовало нормальным величинам (376,7+31,1 мМЕ/л). У 8 больных синдром ПКЯ сочетался с функциональной гиперпролактинемией, достигавшей в среднем уровня 1006,0 ±191,8 мМЕ/л. Экскреция 17-КС с мочой (50,9+9,2 мкмоль/сут) и содержание Т в крови (2,89+0,47 нмоль/л) у больных этой группы достоверно не отличались от соответствующих показателей в группе больных с обычной формой синдрома ПКЯ. У 13 больных синдром ПКЯ сочетался с надпочечниковым компонентом гиперандрогенемии. Экскреция 17-КС с мочой у больных этой группы составляла 94,8+ ±12,9 мкмоль/сут и достоверно превышала показатель в контрольной группе (р<0,01) и в группе больных с обычной формой ‘ ПКЯ (р<0,01). Содержание Т в крови (2,00±0,25 нмоль/л) не отличалось от уровня в контрольной группе, а подавление его дексаметазоном достигало 57,4 + ±9,7 % от исходного уровня, что достоверно (р<0,02) превышало степень подавления в группе больных с обычной формой ПКЯ.
Содержание Т в крови 2,75±0,61 нмоль/л достоверно (р<0,02) превышало показатель в контрольной группе (1,42+0,15 нмоль/л), а снижение его при пробе с дексаметазоном достигало лишь 86,7+5,3 % от исходного уровня. Содержание ПРЛ в крови у больных этой группы составляло 269,5+49,5 мМЕ/л и соответствовало нормальным величинам (376,7+31,1 мМЕ/л). У 8 больных синдром ПКЯ сочетался с функциональной гиперпролактинемией, достигавшей в среднем уровня 1006,0 ±191,8 мМЕ/л. Экскреция 17-КС с мочой (50,9+9,2 мкмоль/сут) и содержание Т в крови (2,89+0,47 нмоль/л) у больных этой группы достоверно не отличались от соответствующих показателей в группе больных с обычной формой синдрома ПКЯ. У 13 больных синдром ПКЯ сочетался с надпочечниковым компонентом гиперандрогенемии. Экскреция 17-КС с мочой у больных этой группы составляла 94,8+ ±12,9 мкмоль/сут и достоверно превышала показатель в контрольной группе (р<0,01) и в группе больных с обычной формой ‘ ПКЯ (р<0,01). Содержание Т в крови (2,00±0,25 нмоль/л) не отличалось от уровня в контрольной группе, а подавление его дексаметазоном достигало 57,4 + ±9,7 % от исходного уровня, что достоверно (р<0,02) превышало степень подавления в группе больных с обычной формой ПКЯ. Содержание ПРЛ в крови у больных этой группы (290,5± ±48,4 мМЕ/л) достоверно не отличалось от показателя в контрольной группе.
Содержание ПРЛ в крови у больных этой группы (290,5± ±48,4 мМЕ/л) достоверно не отличалось от показателя в контрольной группе.
Результаты пробы с этимизолом у больных с гиперандрогенемией и у здоровых женщин представлены на рис. 1. Как видно, введение этими-
4 Пробл. ээдооринооогии № 2 зола вызывает у здоровых лиц кратковременное повышение уровня АКТГ и К в крови, после чего он постепенно снижается до исходных величин и ниже. У больных с обычной формой синдрома ПКЯ реакция АКТГ и К на введение этимизола не обличалась от реакции в контрольной группе. (У больных с синдромом ПКЯ и сопутствующей надпочечниковой гипeраодоогеоемией в ответ на введение этимизола отмечался достоверно более высокий прирост содержания АКТГ и К в крови по сравнению с реакцией в контрольной группе и в группе больных с обычной формой синдрома ПКЯ. Поскольку этимизол потенцирует использование собственных эндогенных резервов мо11оамш1<)в, можно полагать, что у больных с ПКЯ и сопутствующей надпочечниковой гипераодрогенемией исходно повышена функциональная активность гипоталамических трансмиттерных систем, регулирующих функцию коры надпочечников. В отличие от контрольной группы содержание АКТГ в крови у больных с ПКЯ и надпочечниковой гипepaодоо- генемией оставалось повышенным до конца пробы, что не может рассматриваться как следствие воздействия этимизола, так как содержание К в крови начинает снижаться после 30 мин пробы. Длительно сохраняющийся повышенный уровень АКТГ в крови в этих условиях является результатом стимулирующего воздействия снижающимся уровнем К по механизму отрицательной обратной связи — его компонента, чувствительного к скорости изменения содержания К в крови. Этот компонент обратной связи опосредуется выделением кортиколибеоина из центров гипоталамуса [8]. Высокая чувствительность этого механизма, по-видимому, является специфической особенностью больных с надпочечниковой гиперандрогенемией.
В отличие от контрольной группы содержание АКТГ в крови у больных с ПКЯ и надпочечниковой гипepaодоо- генемией оставалось повышенным до конца пробы, что не может рассматриваться как следствие воздействия этимизола, так как содержание К в крови начинает снижаться после 30 мин пробы. Длительно сохраняющийся повышенный уровень АКТГ в крови в этих условиях является результатом стимулирующего воздействия снижающимся уровнем К по механизму отрицательной обратной связи — его компонента, чувствительного к скорости изменения содержания К в крови. Этот компонент обратной связи опосредуется выделением кортиколибеоина из центров гипоталамуса [8]. Высокая чувствительность этого механизма, по-видимому, является специфической особенностью больных с надпочечниковой гиперандрогенемией.
У больных с синдромом ПКЯ и сопутствующей гиперпролактинемией не наблюдалось увеличения
25
а
Рис. 2. Динамика содержания К и АКТГ в крови (в % к исходному уровню) во время пробы с этимизолом у больных с ПКЯ до (/) и на фоне приема (2) левопа (а),ципро- гептадина (б), метоклопрамида (в).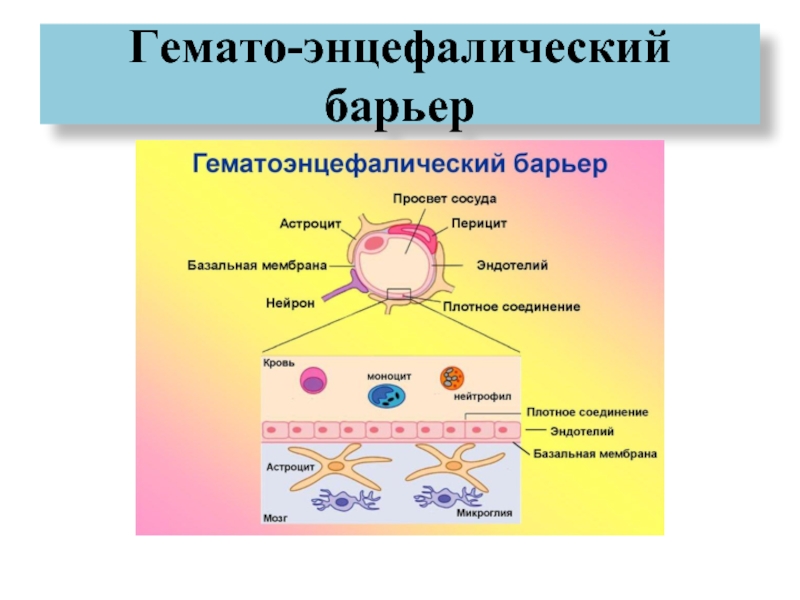
уровня АКТГ и К в крови в ответ на введение этимизола. Повышение содержания АКТГ отмечалось лишь после значительного (на 50 %) снижения уровня К, т. е. по механизму отрицательной обратной связи. Это указывает на определенную недостаточность гипоталамических моноами- нергических систем, регулирующих адренокортикальную функцию у больных с ПКЯ и гиперпролактинемией.
При применении левопа (рис. 2, а) наблюдалось лишь незначительное снижение прироста содержания АКТГ и К в крови в ответ на стимуляцию этимизолом. Это совпадает с представлением, что у здоровых лиц дофамин не играет существенной роли в стимуляции секреции корти- колиберина. Применение ципрогептадина (рис. 2,6) сопровождалось изменением реакции на этимизол, заключающимся в устранении прироста уровня АКТГ и К в крови. Эти изменения, вероятно, связаны с блокадой серотонинергических рецепторов и согласуются с полученными ранее данными об участии серотонинергических систем в стимуляции секреции кортиколиберина [13]. Ципрогептадин оказывает ингибирующее действие на вызванное серотонином выделение кортиколиберина, но не влияет на стимулируемую кортиколиберином секрецию АКТГ гипофизом [ 12].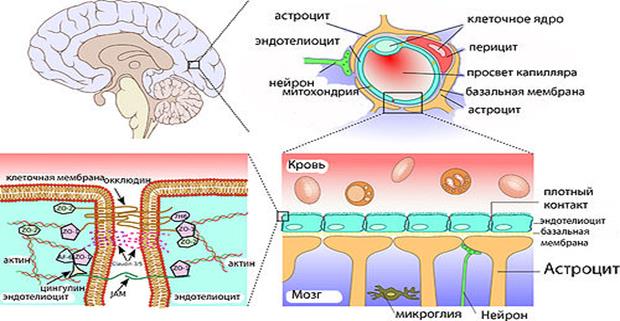
Итак, результаты пробы с этимизолом у больных с синдромом ПКЯ и надпочечниковой гиперандрогенемией могут рассматриваться как проявления повышенной активности гипоталамических серотонинергических систем, регулирующих секрецию кортиколиберина, и, таким образом, гиперандрогенемия у больных этой группы является следствием повышенной функциональной активности системы гипоталамус — гипофиз — кора надпочечников.
Известно, что серотонинергические механизмы опосредуют стимулирующее влияние стрессорных воздействий, в частности инсулиновой гипогликемии, на секрецию ПРЛ. Поэтому возможно, что часто наблюдаемая гиперпролактинемия у больных с надпочечниковым компонентом гиперандро- генемии является также следствием повышенной активности серотонинергических трансмиттерных систем гипоталамуса, регулирующих секрецию ПРЛ. Блокада дофаминергических рецепторов метоклопрамидом (рис. 2, в) сопровождалась устранением прироста уровня АКТГ и К в крови в ответ на введение этимизола. Имеются данные, что метоклопрамид при внутривенном введении оказывает стимулирующее действие на секрецию АКТГ и К, опосредуемое кортиколиберином [11].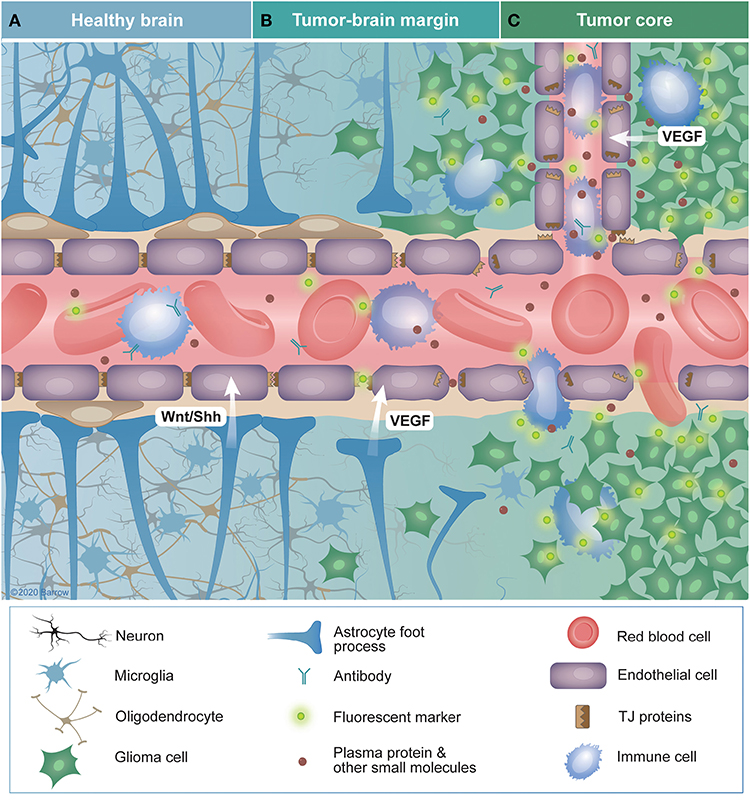 Повторная стимуляция кортиколиберином секреции АКТГ сопровождается угнетением чувствительности кортикотрофов к повторному введению кортиколиберина [15]. По-видимому, сходный механизм наблюдается при применении метоклопрамида перед пробой с этимизолом. Аналогичный тип реакции на этимизол наблюдался у больных с синдромом ПКЯ и гиперпролактинемией (см. рис. 1), на основании чего можно считать, что у больных этой группы имеется недостаточность дофаминергических систем гипоталамуса. Однако тип реакции на этимизол у больных с синдромом ПКЯ и гиперпролактинемией также совпадает с типом реакции на этимизол, проводимой на фоне применения ципрогептадина (см. рис. 2,6), т. е. при блокаде серотонинергических систем. Это позволяет допустить, что у больных данной группы имеется также недостаточность и серотонинергических систем гипоталамуса. Недостаточность трансмиттерных систем гипоталамуса может быть изначальным нарушением, присущим больным с синдромом ПКЯ и гиперпролактинемией, однако возможно, что развитие этой недостаточности является следствием повторных или длительно- действующих нейрогенных стрессорных воздействий.
Повторная стимуляция кортиколиберином секреции АКТГ сопровождается угнетением чувствительности кортикотрофов к повторному введению кортиколиберина [15]. По-видимому, сходный механизм наблюдается при применении метоклопрамида перед пробой с этимизолом. Аналогичный тип реакции на этимизол наблюдался у больных с синдромом ПКЯ и гиперпролактинемией (см. рис. 1), на основании чего можно считать, что у больных этой группы имеется недостаточность дофаминергических систем гипоталамуса. Однако тип реакции на этимизол у больных с синдромом ПКЯ и гиперпролактинемией также совпадает с типом реакции на этимизол, проводимой на фоне применения ципрогептадина (см. рис. 2,6), т. е. при блокаде серотонинергических систем. Это позволяет допустить, что у больных данной группы имеется также недостаточность и серотонинергических систем гипоталамуса. Недостаточность трансмиттерных систем гипоталамуса может быть изначальным нарушением, присущим больным с синдромом ПКЯ и гиперпролактинемией, однако возможно, что развитие этой недостаточности является следствием повторных или длительно- действующих нейрогенных стрессорных воздействий. Не исключено, что у больных с синдромом ПКЯ и гиперпролактинемией дофаминергическая недостаточность присуща также гипоталамическим структурам, входящим в состав тубероинфундибулярной системы. Развитие гиперпролактинемии у больных с синдромом ПКЯ обычно рассматривается как результат избыточной стимуляции лактотрофов гипофиза повышенным содержанием эстрогенов в крови, однако это положение убедительно не доказано [2]. Отсутствуют наблюдения повышенной частоты периферических пролиферативных процессов у больных с ПКЯ при сопутствующей гиперпролактинемии. Несмотря на то что гипер- эстрогенемия характерна для большинства больных с синдромом ПКЯ, гиперпролактинемия развивается лишь приблизительно у ‘/з больных. Полученные данные показывают, что развитие гиперпролактинемии у части больных с синдромом ПКЯ может быть опосредовано гипоталамиче — ской дофаминергической недостаточностью, снижением дофаминергического торможения лакто- тропной функции гипофиза и повышением чувствительности лактотрофов к циркулирующему уровню эстрогенов.
Не исключено, что у больных с синдромом ПКЯ и гиперпролактинемией дофаминергическая недостаточность присуща также гипоталамическим структурам, входящим в состав тубероинфундибулярной системы. Развитие гиперпролактинемии у больных с синдромом ПКЯ обычно рассматривается как результат избыточной стимуляции лактотрофов гипофиза повышенным содержанием эстрогенов в крови, однако это положение убедительно не доказано [2]. Отсутствуют наблюдения повышенной частоты периферических пролиферативных процессов у больных с ПКЯ при сопутствующей гиперпролактинемии. Несмотря на то что гипер- эстрогенемия характерна для большинства больных с синдромом ПКЯ, гиперпролактинемия развивается лишь приблизительно у ‘/з больных. Полученные данные показывают, что развитие гиперпролактинемии у части больных с синдромом ПКЯ может быть опосредовано гипоталамиче — ской дофаминергической недостаточностью, снижением дофаминергического торможения лакто- тропной функции гипофиза и повышением чувствительности лактотрофов к циркулирующему уровню эстрогенов.
Выводы
- У больных с обычной формой синдрома ПКЯ реакция АКТГ и К на введение этимизола не отличалась от типа реакции в контрольной группе.
- У больных с синдромом ПКЯ и надпочечниковой гиперандрогенемией наблюдается повышенная реакция АКТГ и К на введение этимизола, которая устраняется серотониновым антагонистом ципрогептадином. Это свидетельствует о том, что надпочечниковый компонент гиперандрогенемии обусловлен повышенной активностью серотонинергических гипоталамических систем, регулирующих секрецию кортиколиберииа,и,следовательно, гиперандрогенемия является результатом повышенной функциональной активности гипоталамо- гипофизарно-надпочечниковой системы.
- Наблюдаемая у больных с надпочечниковой гиперандрогенемией гиперпролактинемия, возможно, обусловлена повышенной активностью гипоталамических серотонинергических систем,регулирующих секрецию ПРЛ, что подтверждается результатами функциональных проб.
- У больных с синдромом ПКЯ и сопутствующей гиперпролактинемией реакция АКТГ и К на введение этимизола отсутствовала. Подобный тип реакции на введение этимизола наблюдался при блокаде серотониновой активности ципрогептадином и при блокаде дофаминергических рецепторов метоклопрамидом. Это свидетельствует о том, что у больных с синдромом ПКЯ и сопутствующей гиперпролактинемией имеется недостаточность серотонинергических и дофаминергических трансмиттерных систем гипоталамуса.
 Подобный тип реакции на введение этимизола наблюдался при блокаде серотониновой активности ципрогептадином и при блокаде дофаминергических рецепторов метоклопрамидом. Это свидетельствует о том, что у больных с синдромом ПКЯ и сопутствующей гиперпролактинемией имеется недостаточность серотонинергических и дофаминергических трансмиттерных систем гипоталамуса.
Подобный тип реакции на введение этимизола наблюдался при блокаде серотониновой активности ципрогептадином и при блокаде дофаминергических рецепторов метоклопрамидом. Это свидетельствует о том, что у больных с синдромом ПКЯ и сопутствующей гиперпролактинемией имеется недостаточность серотонинергических и дофаминергических трансмиттерных систем гипоталамуса.1. Богданова Е. А. // Акуш. и гин.— 1981.— № 7.— С. 6—8.
2. Дедов И. И.. Мельниченко Г. А., Шилин Д. Е. // Пробл. эндокринол.— 1988.— № 2.— С. 75—83.
3. Комаров Е. К. // Латвийское науч, о—во эндокринологов: Симпозиум: Тезисы докладов.— Рига, 1988.— С. 101—102.
4. Коренева Г. П., Шикаева Ф. В., Ефименко И. Ф. // Акуш. и гин.— 1982.— № 2.— С. 21—23.
Коренева Г. П., Шикаева Ф. В., Ефименко И. Ф. // Акуш. и гин.— 1982.— № 2.— С. 21—23.
5. Рыженков В. Е.. Бехтерева Н. П., Сапронов Н. С. // Фармакол. и токсикол.— 1971.— № 2.— С. 189—191.
6. Сапронов И. С. // Фармакол. и токсикол.— 1979.— № 3.— С. 216—221.
7. Cumming D. С., Reid R. L., Quigley М. Е. et al. // Clin. Endocr.— 1984,— Vol. 20, N 6,— P. 643—648.
8. Jones M. T. (1978). Цит. no: Feek С. M., Marante D. J., Edwards C. R. W. // Clin. Endocr. Metab.— 1983.— Vol. 12, N 3,— P. 597—618.
9. Lobo R. A. // Obstet. Gynec. Clin. N. Amer.— 1987.— Vol. 14, N 4,— P. 955—977.
14, N 4,— P. 955—977.
10. Maroulis G. B. // Fertil. and Steril.— 1981.— Vol. 36, N 3.— P. 273—305.
11. Nishida S., Matsuki M., Adachi N. et al. // J. clin. Endocr.— 1987,— Vol. 65, N 5.— P. 995—1001.
12. Peiletier G., Desy L. // Cell Tiss. Res.— 1979.— Vol. 196.— P. 525—528.
13. Petraglia F., Facchinetti E., Martignoni E. et al. // J. clin. Endocr.— 1984,— Vol. 59, N 6,— P. 1138—1142.
14. Quigley M. E., Rakoff J. S., Yen S. S. C. // Ibid.— 1981.— Vol. 52, N 2,— P. 231—234.
15. Schopohl J., Hauer A., Kaliebe T. et al. // Acta endocr (Kbh.).— 1986,— Vol. 112, N 2 — P. 157—165.
Schopohl J., Hauer A., Kaliebe T. et al. // Acta endocr (Kbh.).— 1986,— Vol. 112, N 2 — P. 157—165.
Мозговой барьер. Научные журналы. Наука и техника
Г. Кассиль, доктор медицинских наук
Энцефалит, менингит, столбняк, рассеянный склероз, дрожательный паралич… Эти заболевания центральной нервной системы пользуются печальной славой трудноизлечимых… В подобных случаях происходит нечто загадочное: инфекция проникает в мозг, а фармакологические препараты, применяемые при лечении нервных заболеваний, туда не попадают – не пропускает так называемый гематоэнцефалический барьер. Врачам приходится искать обходные пути, но поскольку механизм этот изучен еще далеко не достаточно, то медицина часто оказывается беспомощной в борьбе с этими болезнями. Этим проблемам посвящена предлагаемая читателям статья профессора Г.Н. Кассиля.
Возьмем для примера столбняк – тяжелое инфекционное заболевание, поражающее клетки мозга.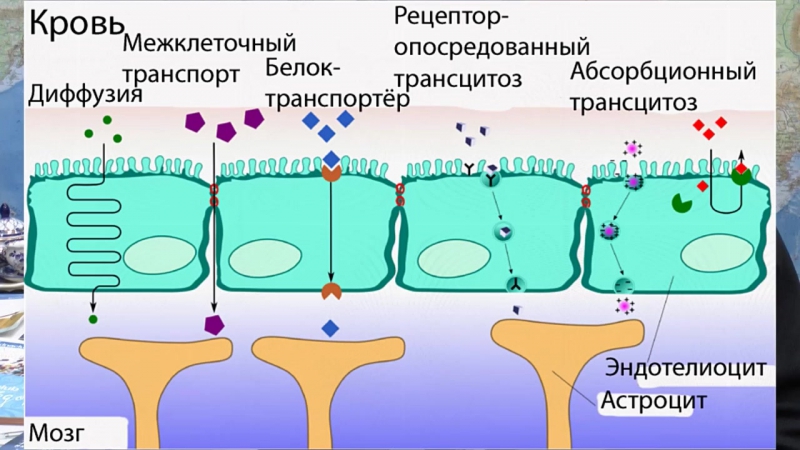 С тех пор, как была изготовлена противостолбнячная сыворотка, число заболеваний столбняком резко снизилось. Эта сыворотка, если она введена в самом начале заболевания, не только предохраняет, но и во многих случаях излечивает от столбняка. Если же недуг уже развился, если столбнячный токсин проник в клетки нервной системы, сыворотка даже в огромных количествах не помогает, так как она не доходит до пораженных ядом нервных клеток. На ее пути появляется какая-то преграда, и больной может погибнуть, несмотря на то, что организм его переполнен антителами, способными обезвредить с избытком весь токсин, накопившийся в нервных клетках. Что же это за преграда?
С тех пор, как была изготовлена противостолбнячная сыворотка, число заболеваний столбняком резко снизилось. Эта сыворотка, если она введена в самом начале заболевания, не только предохраняет, но и во многих случаях излечивает от столбняка. Если же недуг уже развился, если столбнячный токсин проник в клетки нервной системы, сыворотка даже в огромных количествах не помогает, так как она не доходит до пораженных ядом нервных клеток. На ее пути появляется какая-то преграда, и больной может погибнуть, несмотря на то, что организм его переполнен антителами, способными обезвредить с избытком весь токсин, накопившийся в нервных клетках. Что же это за преграда?
Еще в 1885 году выдающийся немецкий микробиолог П. Эрлих обнаружил, что кислые красители, введенные в кровь животного, в мозг не попадают. Прошло немало лет, и сотрудник Эрлиха – Э. Гольдман поставил два ставших знаменитыми опыта с полуколлоидной краской «трипановый синий». Оказалось, что если эту краску ввести в кровь, то она окрашивает все органы, кроме мозга. Если же краска вводится в подмозжечковую цистерну, то окрашивается и вещество мозга. Тогда-то и возникла мысль о существовании сосудистого барьера, как бы запирающего центральную нервную систему от веществ, циркулирующих в крови.
Если же краска вводится в подмозжечковую цистерну, то окрашивается и вещество мозга. Тогда-то и возникла мысль о существовании сосудистого барьера, как бы запирающего центральную нервную систему от веществ, циркулирующих в крови.
Схематическое изображение опытов Э. Гольдмана, в которых было установлено существование преграды для веществ, идущих из крови в мозг. В опыте А краситель вводили в кровь, и мозг оставался неокрашенным. В опыте Б краситель вводили в ликвор (через так называемую подмозжечковую цистерну), мозг окрашивался, но в кровь краситель не попадал. Неодолимая для него граница гематоэнцефалического барьера показана жирной чертой.
От опытов Эрлиха – Гольдмана до современных представлений о мозговом барьере наука прошла длинный и тернистый путь. В начале двадцатых годов нашего столетия фундаментальные работы академика Л.С. Штерн и ее сотрудников заложили учение о гематоэнцефалическом (кровемозговом) барьере.
Потом, как пишет английский ученый М.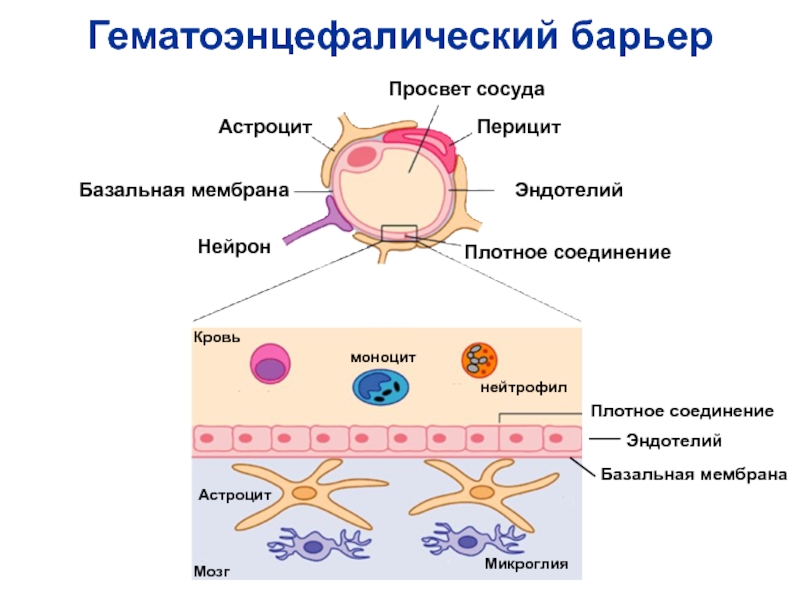 Бредбери в своей монографии «Концепция гематоэнцефалического барьера» (1983), «были времена, когда этот барьер пользовался дурной славой своего рода мифа, в который верили лишь отдельные одержимые физиологи и фармакологи. К счастью, в настоящее время положение изменилось, и я имею возможность объединить большое число экспериментальных данных, не только подтверждающих существование гематоэнцефалического барьера, но и проливающих яркий свет на его функции и ультраструктуру».
Бредбери в своей монографии «Концепция гематоэнцефалического барьера» (1983), «были времена, когда этот барьер пользовался дурной славой своего рода мифа, в который верили лишь отдельные одержимые физиологи и фармакологи. К счастью, в настоящее время положение изменилось, и я имею возможность объединить большое число экспериментальных данных, не только подтверждающих существование гематоэнцефалического барьера, но и проливающих яркий свет на его функции и ультраструктуру».
Мозговой барьер защищает центральную нервную систему от всевозможных чужеродных, ядовитых веществ, проникающих в кровь или образовавшихся в самом организме, способных повредить необычайно чувствительные нервные клетки головного и спинного мозга.
Конечно, не следует думать, что барьер является непреодолимой преградой, какой-то крепостной стеной, отделяющей центральную нервную систему от общей внутренней среды. Непроницаемость его относительна и зависит в значительной степени от количества и концентрации находящихся в крови веществ, от состояния организма, от длительности пребывания вещества в организме, от внешних воздействий и ряда других причин. Анатомические элементы, из которых складывается структура барьера, не только защищают мозг, но и регулируют его жизнедеятельность, питание, выведение продуктов обмена веществ и т.п.
Анатомические элементы, из которых складывается структура барьера, не только защищают мозг, но и регулируют его жизнедеятельность, питание, выведение продуктов обмена веществ и т.п.
Постоянство внутренней среды, в которой живет центральная нервная система человека и животных, является обязательным условием ее деятельности. Природа не случайно спрятала мозг в прочную костную коробку и защитила его от общей внутренней среды организма – крови – сложным, дифференцированным механизмом – мозговым барьером. Даже незначительные изменения в составе окружающей мозг цереброспинальной жидкости (или спинномозговая жидкость, или ликвор), небольшие колебания в поступлении кислорода либо питательных веществ к клеткам мозга оказывают подчас решающее влияние на их состояние. Отсюда и ведущее назначение гематоэнцефалического барьера – поддержание постоянства внутренней среды мозга, регуляция ее состава и биологических свойств. Он как бы оберегает мозг человека и животных от всевозможных случайностей, создает для нервных клеток постоянные условия. Поэтому точная и бесперебойная работа нейронов, а значит, умственная деятельность, психика, настроение, здоровье и болезнь во многом зависят от функционального состояния барьера.
Поэтому точная и бесперебойная работа нейронов, а значит, умственная деятельность, психика, настроение, здоровье и болезнь во многом зависят от функционального состояния барьера.
Какова же анатомия гематоэнцефалического барьера? Над решением этого вопроса уже десятки лет бьются многие поколения экспериментаторов и теоретиков – биологов, морфологов, физиологов, медиков. И то, что еще вчера казалось загадкой, сегодня решено или близко к решению. Конечно, мозговой барьер не орган тела, подобно печени, селезенке или легким. Это совокупность анатомических элементов, которые выполняют роль преграды наряду с другими своими функциями.
Так, основная функция мозговых капилляров – доставлять к мозгу кровь, через их стенки в ткань мозга поступает питание, через них же выводятся отработанные материалы. Обмен этот идет непрерывно, но не все вещества проникают через эти стенки.
Мозговой капилляр – первая линия обороны мозга, искусно построенная, проницаемая для одних веществ, полупроницаемая для других и непроницаемая для третьих.
Вообще строение капилляров, вернее, их внутреннего слоя – эндотелия, отличается в различных тканях и органах по форме ядра, структуре его оболочки и т.п. Вещества переходят из крови в околоклеточную жидкость через мельчайшие поры (щели) между эндотелиальными клетками и сквозь некоторые истонченные участки самих клеток – так называемые окошки, или фенестры.
Стенки мозговых капилляров не имеют ни пор, ни окошек. Отдельные клетки накладываются друг на друга подобно черепицам (гребенчатое строение), и места стыковок прикрыты особыми замыкательными пластинками. Щели между клетками необычайно узкие, поэтому движение жидкости из капилляра в ткань идет в основном сквозь его стенку. Строение самих эндотелиальных клеток мозгового капилляра также отличается некоторыми особенностями. Например, они содержат богатый набор митохондрий, что указывает на высокую активность в них энергетических и обменных процессов. В то же время в клетках капилляров мозга гораздо меньше транспортных пузырьков (вакуолей), причем особенно в той их стороне, что прилегает к просвету капилляра, но на границе с нервной тканью число их несколько выше.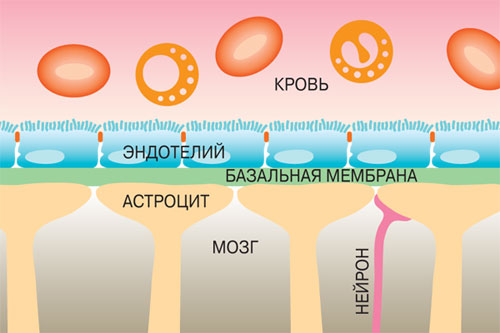 Это указывает, что проницаемость капилляра в направлении из крови в ткани мозга ниже, чем в обратном направлении. Цитоплазматические вакуоли в клетках различных органов обычно участвуют в эвакуации ненужных частиц вещества, перенося их во внеклеточную жидкость. Обратный же процесс, когда клеточная мембрана захватывает извне частицы и отправляет их внутрь клетки, в стенках мозговых капилляров почти полностью отсутствует.
Это указывает, что проницаемость капилляра в направлении из крови в ткани мозга ниже, чем в обратном направлении. Цитоплазматические вакуоли в клетках различных органов обычно участвуют в эвакуации ненужных частиц вещества, перенося их во внеклеточную жидкость. Обратный же процесс, когда клеточная мембрана захватывает извне частицы и отправляет их внутрь клетки, в стенках мозговых капилляров почти полностью отсутствует.
Определенную роль в осуществлении барьерной функции мозговых капилляров играет также расположенная под слоем эндотелиальных клеток прочная трехслойная базальная мембрана со слоем гликокаликса над ней. Составляющие этот слой нити и гранулы образуют своеобразную сеть, которая служит дополнительным препятствием для молекул различных веществ. Кроме того, исследования последних лет показали, что капилляры мозга содержат целый набор ферментов, способных снижать активность химических соединений, поступающих из крови в ткань мозга.
Однако одной лишь стенкой капилляров не ограничивается структура мозгового барьера. Вторая линия обороны расположена между стенкой капилляра и нейронами. Природа поставила здесь сложное сплетение звездчатых клеток (астроцитов) и их отростков (дендритов), образующих слой так называемой нейроглии. Она покрывает около 85 процентов наружной поверхности мозговых капилляров, к которой тесно прилегают присосковые ножки клеток нейроглии. Они способны растягивать просвет капилляра и суживать его. Основная их роль сводится к питанию нейронов. Присосковые ножки высасывают из крови необходимые нейронам питательные вещества и выводят обратно в кровь продукты их обмена веществ (не случайно астроциты получили название «питательных клеток» или «клеток-кормилиц»). При этом нейроглия может менять окислительный потенциал входящих в ее состав элементов, что вызывает изменение электрического заряда клеток и – соответственно – активности мозгового барьера: он становится менее проницаемым, если окислительный потенциал астроцитов повышен.
Вторая линия обороны расположена между стенкой капилляра и нейронами. Природа поставила здесь сложное сплетение звездчатых клеток (астроцитов) и их отростков (дендритов), образующих слой так называемой нейроглии. Она покрывает около 85 процентов наружной поверхности мозговых капилляров, к которой тесно прилегают присосковые ножки клеток нейроглии. Они способны растягивать просвет капилляра и суживать его. Основная их роль сводится к питанию нейронов. Присосковые ножки высасывают из крови необходимые нейронам питательные вещества и выводят обратно в кровь продукты их обмена веществ (не случайно астроциты получили название «питательных клеток» или «клеток-кормилиц»). При этом нейроглия может менять окислительный потенциал входящих в ее состав элементов, что вызывает изменение электрического заряда клеток и – соответственно – активности мозгового барьера: он становится менее проницаемым, если окислительный потенциал астроцитов повышен.
Мозговые капилляры опутаны сложным сплетением астроцитов (звездчатых клеток), их отростков-дендритов и присосковых ножек, которые проникают в эндотелий капилляров.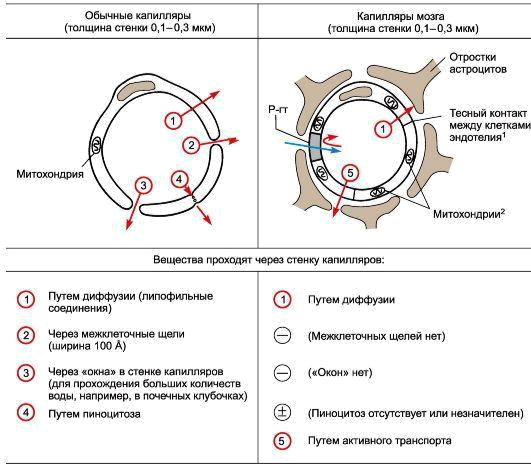 Все вместе они образуют одну из «линий обороны» гематоэнцефалического барьера. 1 – просвет капилляра, 2 – астроциты, 3 – базальная мембрана, 4 – присосковые ножки астроцитов.
Все вместе они образуют одну из «линий обороны» гематоэнцефалического барьера. 1 – просвет капилляра, 2 – астроциты, 3 – базальная мембрана, 4 – присосковые ножки астроцитов.
Но и нейроглией не исчерпывается заслон между кровью и мозгом. Барьерными функциями обладают также окутывающие мозг мягкие оболочки и сосудистые сплетения его боковых желудочков, принимающих активное участие в образовании цереброспинальной жидкости. Проницаемость капилляров сосудистых сплетений несколько выше, чем у капилляров мозга. Щели между эндотелиальными клетками в капиллярах сосудистых сплетений шире, хотя также замкнуты плотными контактами, обращаемыми в сторону полости мозговых желудочков. Здесь проходит третья линия обороны, третья ступень гематоэнцефалического барьера.
В целом же из этих линий образуется объединенный защитный и регуляторный механизм, подобно тому, как из отдельных органов с различным строением и назначением складываются дыхательная, пищеварительная, сердечно-сосудистая, эндокринная и другие системы. Гематоэнцефалический барьер – это мозаика приспособительных механизмов головного и спинного мозга, или, образно выражаясь, федерация автономных, но взаимосвязанных составных частей не только анатомического, но и физиологического механизма.
Гематоэнцефалический барьер – это мозаика приспособительных механизмов головного и спинного мозга, или, образно выражаясь, федерация автономных, но взаимосвязанных составных частей не только анатомического, но и физиологического механизма.
Боковые желудочки мозга, участвующие в образовании ликвора, – тоже часть гематоэнцефалического барьера. Здесь «линия обороны» проходит через капилляры сосудистых сплетений. 1, 2 – боковые желудочки, 3 – подмозжечковая цистерна, 4 – ликвороносные пространства головного мозга.
Как уже было отмечено, функцию мозгового барьера составляет не только защита мозга от чужеродных и ядовитых веществ, циркулирующих в крови, но и регуляция состава и свойств питательной среды, в которой живут нервные клетки головного и спинного мозга. Но каким образом получают они необходимые для жизнедеятельности химические вещества? Только ли прямым путем через присосковые ножки клеток-кормилиц? Или, как и в других органах, также и через тканевую, внеклеточную жидкость? Иными словами, является ли ликвор той непосредственной питательной средой, откуда нервные клетки получают все необходимые для жизни материалы и куда «сдают» свои отходы?
Созданная еще в 1917. ..1921 гг. теория К. Монакова – Л. Штерн утверждала, что питание мозга идет только через ликвор. Считалось, что эта прозрачная, бесцветная, как дистиллированная вода, жидкость окружает весь мозг, глубоко проникая в его толщу, и каждая его клетка как бы купается в этой питательной среде и находит в ней все необходимое для жизни и развития. Отметим, что эта теория – пример точного научного предвидения, поскольку тогда не существовало еще современных методов исследования мозга, и потому нельзя было точно сказать, какими же путями проникает ликвор в глубины мозга. Ничего не было известно о наличии в ткани мозга околоклеточных пространств, о существовании прямых связей между ними и теми пространствами под оболочками мозга, где циркулирует спинномозговая жидкость. И потому во всех монографиях, посвященных этой жидкости, и сейчас еще написано, что она не просачивается сквозь ткань мозга, не связана с внеклеточными пространствами в этой ткани, и вещества, попавшие в желудочки мозга или в ликворные резервуары, не доходят до тех нервных клеток, которые расположены глубже, чем на 2.
..1921 гг. теория К. Монакова – Л. Штерн утверждала, что питание мозга идет только через ликвор. Считалось, что эта прозрачная, бесцветная, как дистиллированная вода, жидкость окружает весь мозг, глубоко проникая в его толщу, и каждая его клетка как бы купается в этой питательной среде и находит в ней все необходимое для жизни и развития. Отметим, что эта теория – пример точного научного предвидения, поскольку тогда не существовало еще современных методов исследования мозга, и потому нельзя было точно сказать, какими же путями проникает ликвор в глубины мозга. Ничего не было известно о наличии в ткани мозга околоклеточных пространств, о существовании прямых связей между ними и теми пространствами под оболочками мозга, где циркулирует спинномозговая жидкость. И потому во всех монографиях, посвященных этой жидкости, и сейчас еще написано, что она не просачивается сквозь ткань мозга, не связана с внеклеточными пространствами в этой ткани, и вещества, попавшие в желудочки мозга или в ликворные резервуары, не доходят до тех нервных клеток, которые расположены глубже, чем на 2.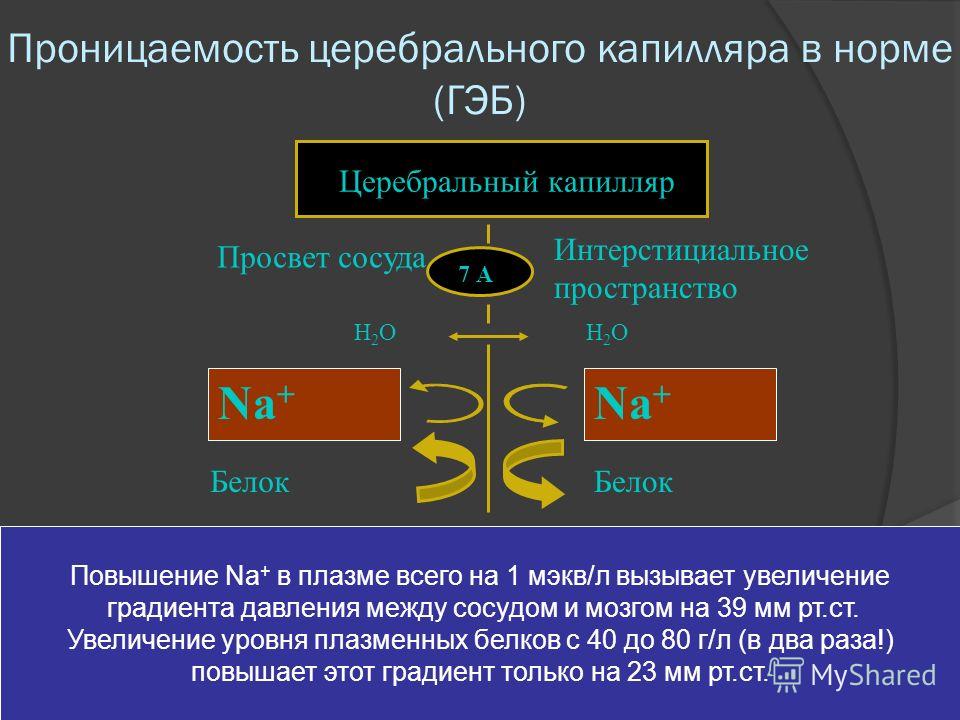 ..3 мм от поверхности мозга. Даже принято считать, что есть еще один барьер – между ликвором и мозгом (так называемый ликворо-церебральный барьер).
..3 мм от поверхности мозга. Даже принято считать, что есть еще один барьер – между ликвором и мозгом (так называемый ликворо-церебральный барьер).
Но исследования последних лет установили: в ткани мозга существуют внеклеточные пространства. А совсем недавно советский ученый В.А. Отеллин, изучая с помощью электронной микроскопии структуру участков, где ликвор соприкасается с тканью головного мозга, пришел к выводу, что на дне микроборозд мозга имеются проходы, открывающиеся в межклеточные промежутки.
Показан условный участок мозговой ткани – окружение одного из капилляров, расположенного, скажем, где-то в сером веществе. Капилляры мозга в отличие от других тканей не имеют пор, через которые различные вещества, например, кислород, могли бы проникать из крови в мозг. Те немногие, которым это удается (тот же кислород), проходят через стенки капилляра, используя специальный механизм. Для остальных стенка капилляра – это стенка, барьер. Но если между кровью и мозгом существует барьер, то как же доставляются тканям необходимые вещества? В значительной степени через ликвор. Однако до недавних пор не было известно, насколько глубоко в ткань мозга проникает ликвор, есть ли для этого необходимые пути. В последние годы установлено, что ликвор из зон его концентрации (это желудочки мозга, подпаутинное пространство) проходит в глубины мозга через отверстия в микробороздах и распределяется там по межклеточным пространствам (на схеме обозначены голубым цветом).
Однако до недавних пор не было известно, насколько глубоко в ткань мозга проникает ликвор, есть ли для этого необходимые пути. В последние годы установлено, что ликвор из зон его концентрации (это желудочки мозга, подпаутинное пространство) проходит в глубины мозга через отверстия в микробороздах и распределяется там по межклеточным пространствам (на схеме обозначены голубым цветом).
1 – кровеносный капилляр; 2 – нейроны; 3 – астроциты; 4 – межклеточные пространства; 5 – полость желудочков мозга, заполненная ликвором; 6 – слой эпителиальных клеток, выстилающих внутреннюю поверхность желудочков, между ними видны проходы, пропускающие ликвор; 7 – клетка нейроглии; 8 – подпаутинное пространство мозга, насыщенное ликвором; 9 – наружная мозговая мембрана с проходами для ликвора. (По В.А. Отеллину.)
Следовательно, цереброспинальная жидкость может проникнуть в глубины мозга и благодаря этому служить непосредственной питательной средой для нейронов.
Эта точка зрения подтверждается и с другой стороны. Экспериментаторам и врачам-невропатологам хорошо известно, что при нарушениях состава и свойств ликвора изменяются реактивность и возбудимость соприкасающихся с ним нервных клеток. Значит, ликвор влияет на их физиологическое состояние, активность и деятельность.
Экспериментаторам и врачам-невропатологам хорошо известно, что при нарушениях состава и свойств ликвора изменяются реактивность и возбудимость соприкасающихся с ним нервных клеток. Значит, ликвор влияет на их физиологическое состояние, активность и деятельность.
Несомненный интерес представляют в этом отношении сдвиги в содержании ионов калия и кальция. Обычно калия в цереброспинальной жидкости больше, чем кальция, и соотношение этих веществ равно примерно 2:1. Наши исследования показали, что при различных воздействиях на организм эта цифра иногда повышается до 3:1 (например, при сильном возбуждении), иногда падает до 1:1 и ниже. В большинстве случаев накопление калия в ликворе соответствует повышению возбудимости нервных центров, а уменьшение количества калия, наоборот, ее снижению. Так, при наркозе, при длительной бессоннице уровень калия в ликворе падает и нарастает содержание кальция.
Одно за другим открывают ученые все новые и новые биологически активные вещества (гормоны, медиаторы, модуляторы), передающие межклеточную информацию в головном мозге. Именно через ликвор они вступают в сложнейшие химические реакции, протекающие в нервной ткани мозга. Действуя на многочисленные рецепторы головного мозга, они передают гуморальную (то есть идущую не по нервным, а по сосудистым путям, вместе с жидкостью) информацию из одной мозговой точки в другую. В потоках, ручейках и «морях» церебророспинальной жидкости можно обнаружить десятки (по современным данным свыше 50) биологически активных веществ, как образовавшихся в мозгу, так и поступивших из крови.
Именно через ликвор они вступают в сложнейшие химические реакции, протекающие в нервной ткани мозга. Действуя на многочисленные рецепторы головного мозга, они передают гуморальную (то есть идущую не по нервным, а по сосудистым путям, вместе с жидкостью) информацию из одной мозговой точки в другую. В потоках, ручейках и «морях» церебророспинальной жидкости можно обнаружить десятки (по современным данным свыше 50) биологически активных веществ, как образовавшихся в мозгу, так и поступивших из крови.
Наши исследования показали, что химический состав, физико-химические и биологические свойства ликвора определяются проницаемостью мозгового барьера в направлении кровь – ликвор и обратно, то есть, иными словами, темпом перехода веществ из ликвора в мозг и скоростью выведения из него продуктов метаболизма в лимфу и кровь.
Таким образом, наряду с непосредственным поступлением питательных веществ из капилляров в нервные клетки, наряду с деятельностью клеток-кормилиц существует «путь через ликвор», который осуществляет дополнительное питание нейронов.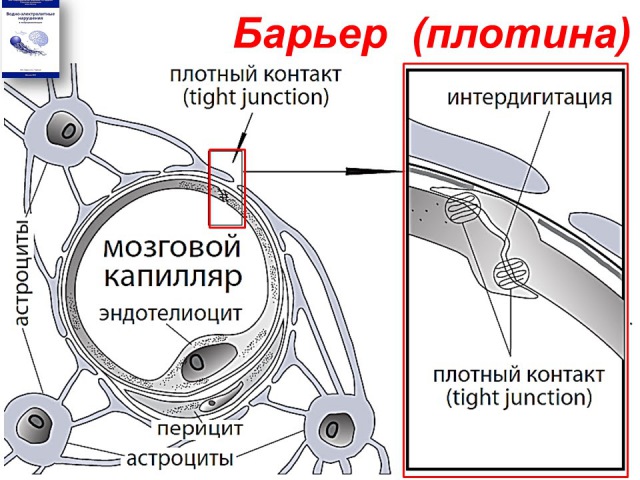
Но этим же путем, как мы видели, в мозг поступает и вся гуморальная информация из внутренней среды организма. Тем самым физиологическое значение гематоэнцефалического барьера значительно расширяется. Регулируя переход электролитов, гормонов, медиаторов, метаболитов из крови в мозг, то есть пропуская одни из них и задерживая другие, барьер регулирует тем самым деятельность всего организма. Поэтому нарушение нормального состояния барьера, его функций, что выражается обычно в увеличении или, наоборот, задержке поступления информации из внутренней среды, может в конечном счете привести к дезорганизации физиологических процессов. Нарушение барьерных функций в первую очередь отражается на составе и свойствах спинномозговой жидкости, в которой изменяется содержание биологически активных веществ. Это приводит к расстройству регуляторных механизмов и, как следствие, к перестройке биохимических и физиологических процессов.
Гематоэнцефалический барьер очень тонко реагирует на изменение условий среды и потребностей мозга. Проницаемость его увеличивается при голодании и гипоксии, под влиянием определенных фармакологических препаратов, при удалении некоторых эндокринных желез (щитовидной, гипофиза, поджелудочной), при повышении температуры тела до 41…42° или при снижении ее до 34…35°, при бессоннице, наркозе и утомлении. Многие инфекционные заболевания, беременность, черепно-мозговая травма, облучение, особенно рентгеновскими лучами, могут уменьшать проницаемость барьера и облегчать поступление в мозг как чужеродных, так и свойственных организму веществ.
Проницаемость его увеличивается при голодании и гипоксии, под влиянием определенных фармакологических препаратов, при удалении некоторых эндокринных желез (щитовидной, гипофиза, поджелудочной), при повышении температуры тела до 41…42° или при снижении ее до 34…35°, при бессоннице, наркозе и утомлении. Многие инфекционные заболевания, беременность, черепно-мозговая травма, облучение, особенно рентгеновскими лучами, могут уменьшать проницаемость барьера и облегчать поступление в мозг как чужеродных, так и свойственных организму веществ.
Но все это известно главным образом из наблюдений за больными, а объяснить, почему это так происходит, невозможно, потому что до сих пор мало что известно о механизмах перехода веществ из крови в мозг. Почему одни вещества легко проникают в головной и спинной мозг, в то время как другие, даже близкие к ним по химическому строению и биологическим свойствам, встречают на пути труднопреодолимую преграду? Высказывалось предположение, что здесь действует некий закон целесообразности: что-де существуют физиологически адекватные вещества, без которых нервные клетки не могут обойтись, и поэтому для них проход открыт, а другие вещества физиологически неадекватны, и барьер их задерживает «у входа» в центральную нервную систему.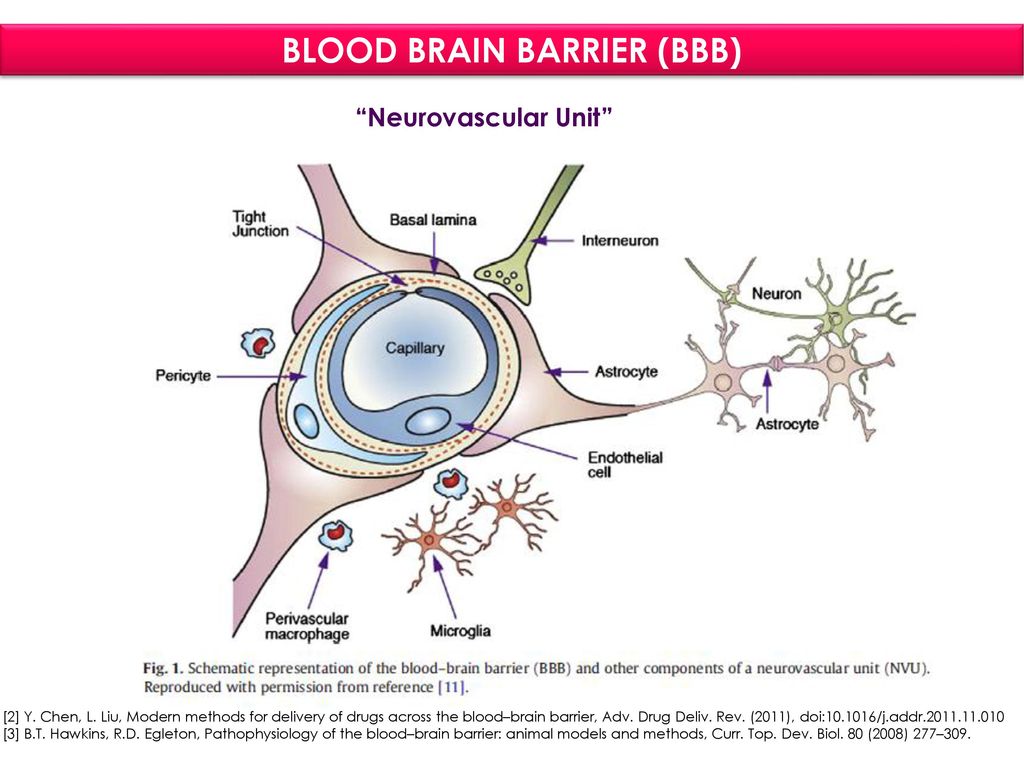 Но тогда почему он задерживает такие подчас жизненно необходимые мозгу вещества, как антитела, многие антибиотики, ряд гормонов, медиаторов, метаболитов? Почему пропускает одни фармакологические препараты и задерживает другие, совершенно необходимые для ликвидации патологических очагов в мозгу или нейтрализации накопившихся в нем токсинов? Ответов на эти вопросы пока нет, и специалистам не остается ничего иного, как идти на «прорыв» барьера: ученым – ради познания его природы, врачам – ради помощи больным в трудных случаях.
Но тогда почему он задерживает такие подчас жизненно необходимые мозгу вещества, как антитела, многие антибиотики, ряд гормонов, медиаторов, метаболитов? Почему пропускает одни фармакологические препараты и задерживает другие, совершенно необходимые для ликвидации патологических очагов в мозгу или нейтрализации накопившихся в нем токсинов? Ответов на эти вопросы пока нет, и специалистам не остается ничего иного, как идти на «прорыв» барьера: ученым – ради познания его природы, врачам – ради помощи больным в трудных случаях.
Однако искусственно изменить состояние мозгового барьера удается не всегда. Многие химические соединения и лекарственные препараты, именно те, которые чаще всего применяются для экспериментальных и терапевтических целей, не проникают в мозг даже при очень сильных воздействиях на организм. Иногда они проникают в незначительном количестве, недостаточном для того, чтобы получить нужный эффект – подавить рост бактерий или опухолевых клеток, нейтрализовать токсины. А при искусственном нарушении барьера в мозг нередко начинают поступать наряду с лечебными препаратами ядовитые шлаки тканевого обмена.
А при искусственном нарушении барьера в мозг нередко начинают поступать наряду с лечебными препаратами ядовитые шлаки тканевого обмена.
В лабораторном эксперименте применяют самые различные способы повышения проницаемости барьера: вдыхание углекислого газа, гипоксию, электрошок, перегревание организма, введение в кровь повышающих или понижающих давление растворов и т.п. В клинической практике наиболее эффективным методом является непосредственное введение лекарственных препаратов в желудочки мозга или в подмозжечковую цистерну – это уже не «прорыв», а «обход» барьера. Он с успехом был использован при лечении ряда инфекционных заболеваний головного мозга – таких, как столбняк, туберкулезный менингит, энцефалиты, воспаление мозговых оболочек, и др.
Нами предложен также метод «обхода» барьера путем введения некоторых витаминов, транквилизаторов, электролитов, биологически активных веществ в слизистую оболочку носа, откуда они попадают в цереброспинальную жидкость и определенные участки головного мозга.
Как видим, о гематоэнцефалическом барьере уже многое известно, но еще больше проблем остается нерешенными, многое требует дальнейшей разработки. Но то, что мы уже знаем, заслуживает самого пристального внимания как теоретиков, так и клиницистов, ибо помогает понять многие стороны деятельности центральной нервной системы человека и животных.
Ранее опубликовано:
Наука и жизнь. 1986. №11.
Дата публикации:
7 февраля 2003 года
Геб египетский бог | Геб Бог Земли
Геб — один из важнейших богов древнего Египта, известный как Бог земли. Считается, что он происходил из важной линии богов и был сыном Шу, который был богом Воздуха, и Тефнут, богиней влаги. У Геба и Нут было четверо детей: Осирис, Исида, Сет и Нефтида. Однако в различной древней литературе Геб также упоминается по-разному, как Себ, Кеб или Гебб. После Атума четыре божества Шу, Тефнут, Геб и Нут пребывали исключительно в Космосе, в то время как второй набор божеств Осирис, Исида, Сет и Нефтис заступался между людьми и космосом. Древние египтяне считали, что происходящие землетрясения вызывают смех Геба.
Древние египтяне считали, что происходящие землетрясения вызывают смех Геба.
Изображение Геба
Геб обычно изображался как человек, носящий комбинацию белой короны и короны Атефа, хотя он также изображался как гусь, его священное животное. В образе человека Геб изображен как олицетворение земли и показан зеленым цветом с растительностью, растущей из его тела. Как земля, его часто видят лежащим на боку, склонив одно колено к небу.В композициях и гимнах Геб — очень важный бог в Книге мертвых, который играет роль в наблюдении за взвешиванием сердца мертвых в Судном зале Осириса.
Происхождение Геба
Гелиополис считается местом рождения богов, которым поклонялись в Египте. Говорят, что здесь началась работа творения. Многие папирусы указывают на это и даже показывают, что после того, как бог Солнца появился в небе, он поднялся в небо и послал свои лучи на землю.Эти папирусы даже показывают, что Геб занимает видное положение, где он изображен лежащим на земле с одной рукой, протянутой на ней, а другой указывающей на небо. Во времена Птолемеев Геб стал отождествляться с богом Кроносом, которому поклонялись в греческой мифологии.
Во времена Птолемеев Геб стал отождествляться с богом Кроносом, которому поклонялись в греческой мифологии.
Считается, что поклонение Богу Гебу началось в Луну в додинастическую эпоху, когда ему в основном поклонялись как гусю, который является его священным животным. Эдфу назывался «Аат Геба», а Дендера также был известен как «дом детей Геба».Наличие бога земли мужского пола может показаться необычным, но некоторые говорят, что Геб был бисексуалом. Говорят, что в своем святилище в Бате он отложил большое яйцо, из которого восстал бог солнца в форме феникса или Бенбена. Геб дал прозвище «великий кудахтанье» из-за звука, который раздавался, когда яйцо откладывалось.
Множество ролей Геба
Говорят, что землетрясения были вызваны смехом Геба, и именно он предоставил драгоценные камни и минералы, которые можно было найти в пещерах и шахтах, поэтому он был признан богом мира шахты и пещеры.Как бог урожая, он также иногда считался супругом Рененутет, богини кобры.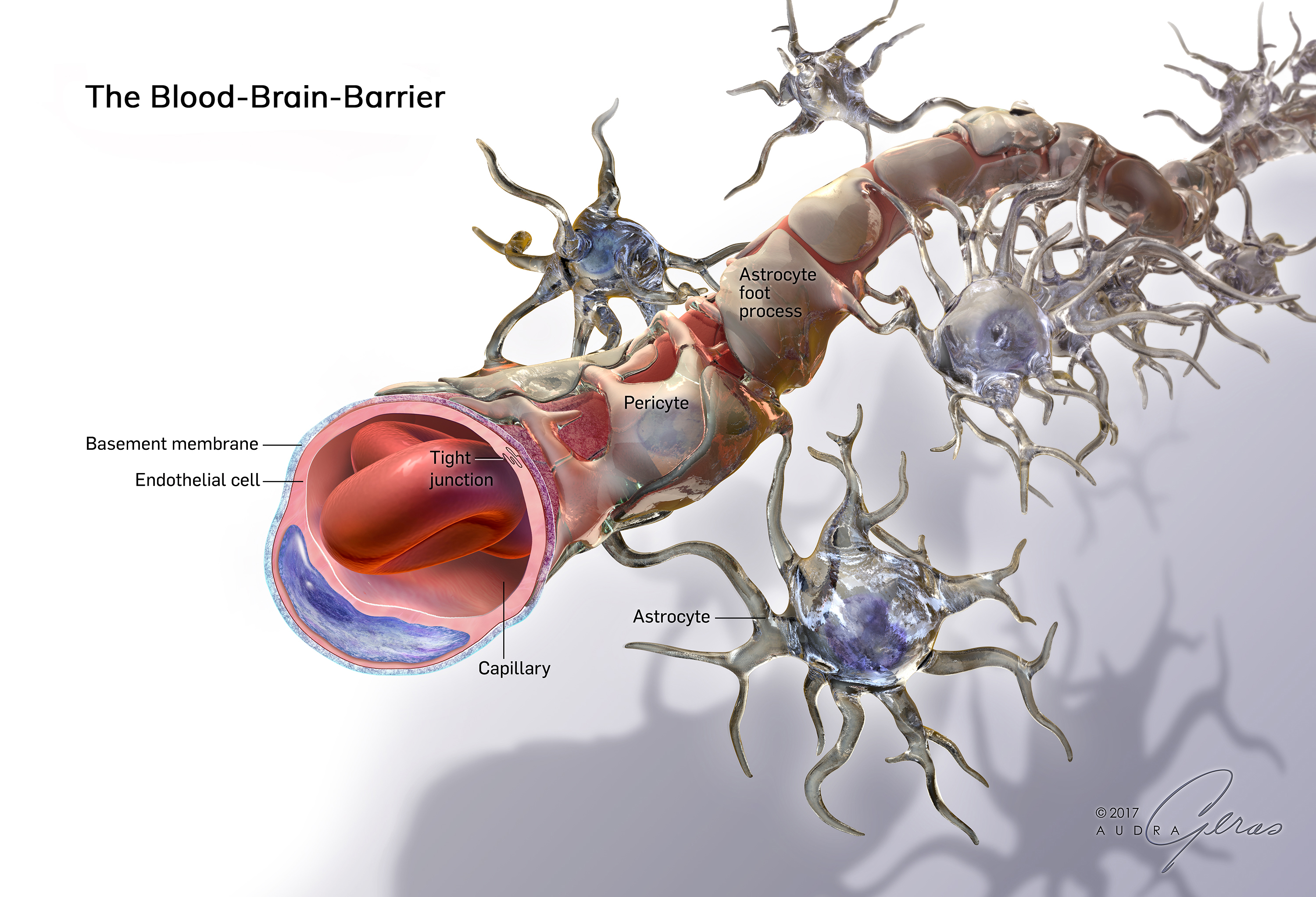
Снег является важным элементом контроля функционального состава растительных сообществ в ороарктической тундре.
Оже С., Шипли Б. (2013) Межвидовые и внутривидовые вариации признаков при небольших градиентах окружающей среды в старовозрастных лесах умеренного климата. J Veg Sci 24: 419–428. https://doi.org/10.1111/j.1654-1103.2012.01473.x
Артикул
Google ученый
Bates D, Mächler M, Bolker B, Walker S (2015) Подгонка линейных моделей смешанных эффектов с использованием lme4.J Stat Soft Artic 67: 1–48. https://doi.org/10.18637/jss.v067.i01
Артикул
Google ученый
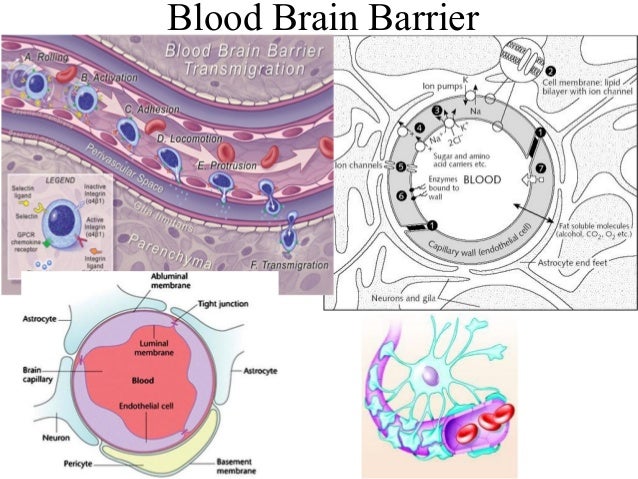 org/ScholarlyArticle»>
org/ScholarlyArticle»> Bjorkman AD, Myers-Smith IH, Elmendorf SC, Normand S, Rüger N, Beck PSA, Blach-Overgaard A, Blok D, Cornelissen JHC, Forbes BC, Georges D, Goetz SJ, Guay KC, Henry GHR, HilleRisLambers J, Hollister RD, Karger DN, Kattge J, Manning P, Prevéy JS, Rixen C, Schaepman-Strub G, Thomas HJD, Vellend M, Wilmking M, Wipf S, Carbognani M, Hermanutz L, Lévesque E, Molau U, Petraglia A, Soudzilovskaia NA, Spasojevic MJ, Tomaselli M, Vowles T, Alatalo JM, Alexander HD, Anadon-Rosell A, Angers-Blondin S, te Beest M, Berner L, Björk RG, Buchwal A, Buras A, Christie K, Купер Э.Дж., Дуллингер С., Эльберлинг Б., Эскелинен А., Фрей Э.Р., Грау О, Гроган П., Халлинджер М., Харпер К.А., Хейманс ММПД, Хадсон Дж., Хюльбер К., Итуррат-Гарсия М., Иверсен С.М., Ярошинска Ф., Джонстон Дж. Ф., Йоргенсен RH, Kaarlejärvi E, Klady R, Kuleza S, Kulonen A, Lamarque LJ, Lantz T, Little CJ, Speed JDM, Michelsen A, Milbau A, Nabe-Nielsen J, Nielsen SS, Ninot JM, Oberbauer SF, Olofsson J, Онипченко В.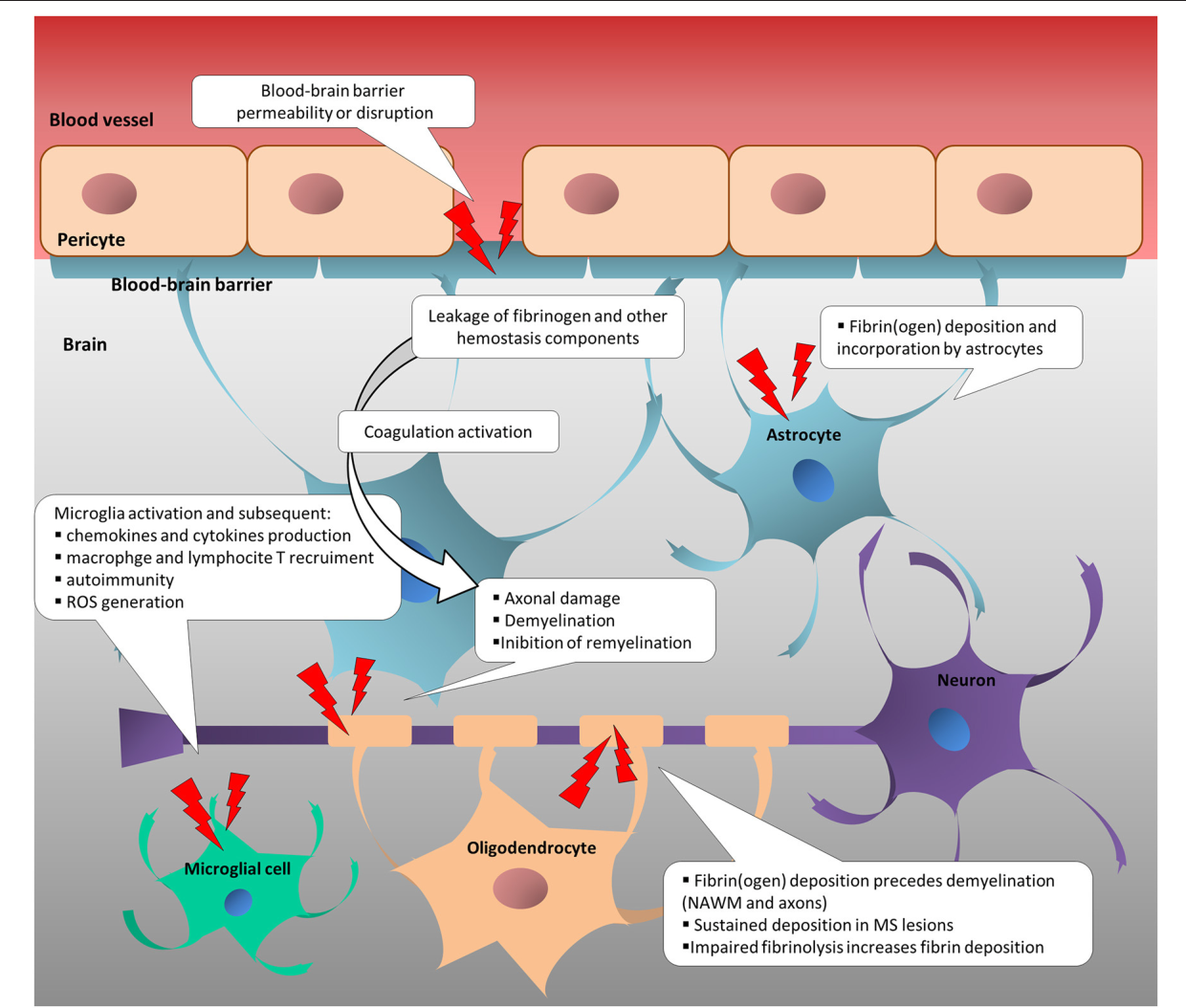 Г., Рампф С.Б., Сем Энчук П., Шетти Р., Кольер Л.С., Стрит ЛЭ, Судинг К.Н., Лента КД, Трант А, Трейер UA, Тремблей Дж. П., Тремблей М., Венн С., Вейерс С., Замин Т., Буланже-Лапойнт Н., Гулд В.А., Хик Д. Hofgaard A, Jónsdóttir IS, Jorgenson J, Klein J, Magnusson B, Tweedie C, Wookey PA, Bahn M, Blonder B, van Bodegom PM, Bond-Lamberty B, Campetella G, Cerabolini BEL, Chapin FS, Cornwell WK, Craine J , Dainese M, de Vries FT, Díaz S, Enquist BJ, Green W, Milla R, Niinemets Ü, Onoda Y, Ordoñez JC, Ozinga WA, Penuelas J, Poorter H, Poschlod P, Reich PB, Sandel B, Schamp B, Шереметев С., Вейхер Э. (2018a) Изменение функциональных характеристик растений в биоме теплой тундры.Природа 562: 57–62. https://doi.org/10.1038/s41586-018-0563-7
Г., Рампф С.Б., Сем Энчук П., Шетти Р., Кольер Л.С., Стрит ЛЭ, Судинг К.Н., Лента КД, Трант А, Трейер UA, Тремблей Дж. П., Тремблей М., Венн С., Вейерс С., Замин Т., Буланже-Лапойнт Н., Гулд В.А., Хик Д. Hofgaard A, Jónsdóttir IS, Jorgenson J, Klein J, Magnusson B, Tweedie C, Wookey PA, Bahn M, Blonder B, van Bodegom PM, Bond-Lamberty B, Campetella G, Cerabolini BEL, Chapin FS, Cornwell WK, Craine J , Dainese M, de Vries FT, Díaz S, Enquist BJ, Green W, Milla R, Niinemets Ü, Onoda Y, Ordoñez JC, Ozinga WA, Penuelas J, Poorter H, Poschlod P, Reich PB, Sandel B, Schamp B, Шереметев С., Вейхер Э. (2018a) Изменение функциональных характеристик растений в биоме теплой тундры.Природа 562: 57–62. https://doi.org/10.1038/s41586-018-0563-7
CAS
Статья
PubMed
Google ученый
Bjorkman AD, Myers-Smith IH, Elmendorf SC, Normand S, Thomas HJD, Alatalo JM, Alexander H, Anadon-Rosell A, Angers-Blondin S, Bai Y, Baruah G, te Beest M, Berner L, Björk RG, Blok D, Bruelheide H, Buchwal A, Buras A, Carbognani M, Christie K, Collier LS, Cooper EJ, Cornelissen JHC, Dickinson KJM, Dullinger S, Elberling B, Eskelinen A, Forbes BC, Frei ER, Iturrate- Гарсия М. , Хороший МК, Грау О, Грин П., Греве М., Гроган П., Хайдер С., Хайек Т., Халлинджер М., Хаппонен К., Харпер К.А., Хейманс MMPD, Генри GHR, Херманутц Л., Хьюитт RE, Холлистер Р. , Hülber K, Iversen CM, Jaroszynska F, Jiménez-Alfaro B, Johnstone J, Jorgensen RH, Kaarlejärvi E, Klady R, Klimešová J, Korsten A, Kuleza S, Kulonen A, Lamarque LJ, Lantz T, Lavalle JJ, , Lévesque E, Little CJ, Luoto M, Macek P, Mack MC, Mathakutha R, Michelsen A, Milbau A, Molau U, Morgan JW, Mörsdorf MA, Nabe-Nielsen J, Nielsen SS, Ninot JM, Oberbauer SF, Olofsson J , Онипченко В.Г., Петраглиа А., Пикеринг К., Prevéy JS, Rixen C, Rumpf SB, Schaepman-Strub G, Semenchuk P, Shetti R, Soudzilovskaia NA, Spasojevic MJ, Speed JDM, Street LE, Suding K, Tape KD, Tomaselli M, Trant A, Treier UA, Tremblay JP, Tremblay M, Venn S, Virkkala AM, Vowles T, Weijers S, Wilmking M, Wipf S, Zamin T (2018b) Tundra Trait Team: база данных характеристик растений, охватывающих биом тундры.Glob Ecol Biogeogr 27: 1402–1411. https://doi.
, Хороший МК, Грау О, Грин П., Греве М., Гроган П., Хайдер С., Хайек Т., Халлинджер М., Хаппонен К., Харпер К.А., Хейманс MMPD, Генри GHR, Херманутц Л., Хьюитт RE, Холлистер Р. , Hülber K, Iversen CM, Jaroszynska F, Jiménez-Alfaro B, Johnstone J, Jorgensen RH, Kaarlejärvi E, Klady R, Klimešová J, Korsten A, Kuleza S, Kulonen A, Lamarque LJ, Lantz T, Lavalle JJ, , Lévesque E, Little CJ, Luoto M, Macek P, Mack MC, Mathakutha R, Michelsen A, Milbau A, Molau U, Morgan JW, Mörsdorf MA, Nabe-Nielsen J, Nielsen SS, Ninot JM, Oberbauer SF, Olofsson J , Онипченко В.Г., Петраглиа А., Пикеринг К., Prevéy JS, Rixen C, Rumpf SB, Schaepman-Strub G, Semenchuk P, Shetti R, Soudzilovskaia NA, Spasojevic MJ, Speed JDM, Street LE, Suding K, Tape KD, Tomaselli M, Trant A, Treier UA, Tremblay JP, Tremblay M, Venn S, Virkkala AM, Vowles T, Weijers S, Wilmking M, Wipf S, Zamin T (2018b) Tundra Trait Team: база данных характеристик растений, охватывающих биом тундры.Glob Ecol Biogeogr 27: 1402–1411. https://doi.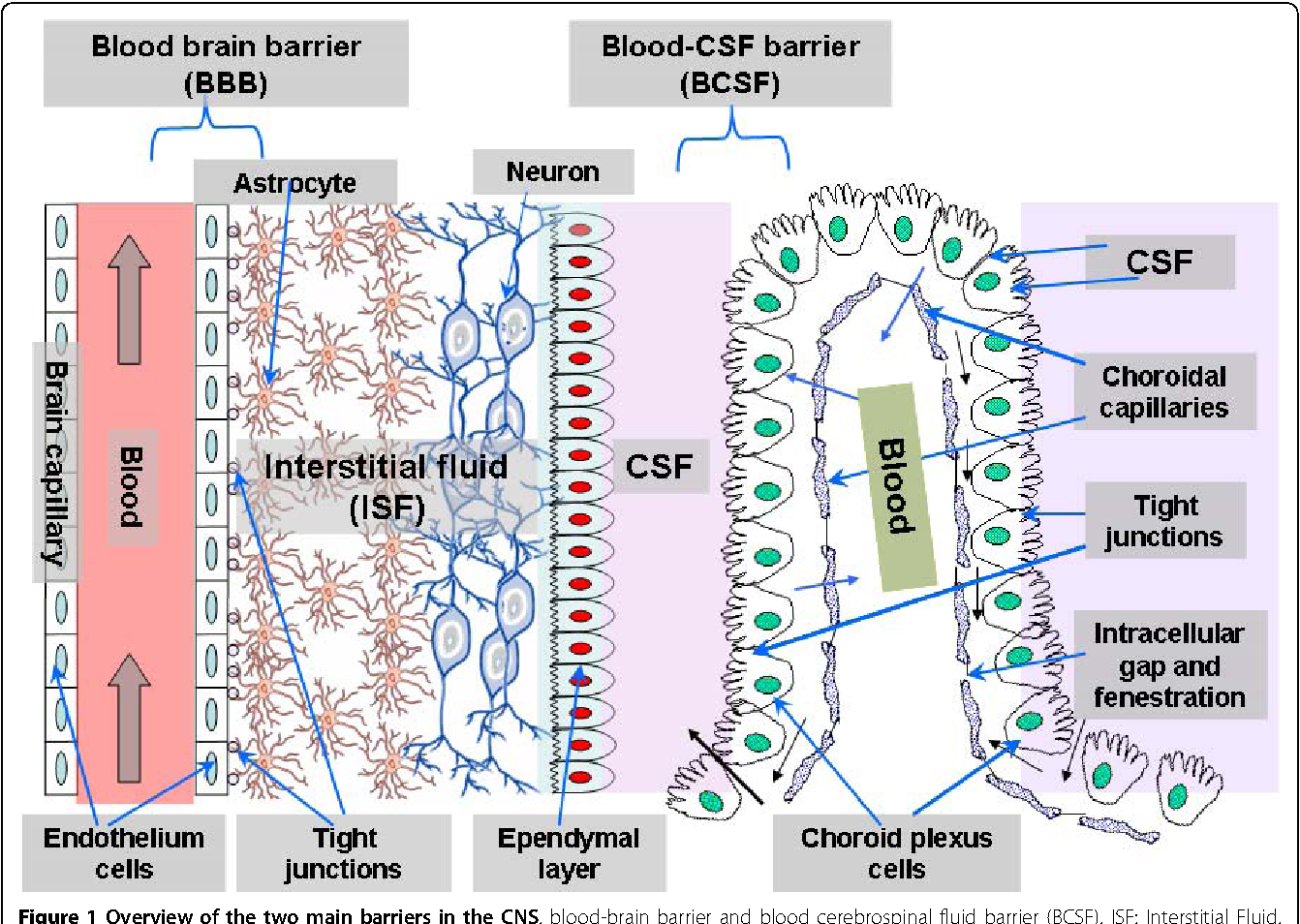 org/10.1111/geb.12821
org/10.1111/geb.12821
Артикул
Google ученый
Браун-Бланке Дж, Конард Х.С., Фуллер Г.Д. (1932) Социология растений; изучение растительных сообществ, 1-е изд. McGraw-Hill Book Company, Inc., Нью-Йорк
Google ученый
Брюльхайде Х, Денглер Дж., Пуршке О, Ленуар Дж., Хименес-Альфаро Б., Хеннекенс С.М., Ботта-Дукат З., Читри М., Филд Р, Янсен Ф., Каттге Дж., Пиллар В.Д., Шродт Ф., Махеха М.Д., Peet RK, Sandel B, van Bodegom P, Altman J, Alvarez-Dávila E, Khan MASA, Attorre F, Aubin I, Baraloto C, Barroso JG, Bauters M, Bergmeier E, Biurrun I, Bjorkman AD, Blonder B, Čarni A , Cayuela L, Černý T, Cornelissen JHC, Craven D, Dainese M, Derroire G, Sanctis MD, Díaz S, Doležal J, Farfan-Rios W, Feldpausch TR, Fenton NJ, Garnier E, Guerin GR, Gutierrez AG, Haider S , Hattab T, Henry G, Hérault B, Higuchi P, Hölzel N, Homeier J, Jentsch A, Jürgens N, Kącki Z, Karger DN, Kessler M, Kleyer M, Knollová I, Korolyuk AY, Kühn I, Laughlin DC, Lens F, Loos J, Louault F, Любенова М. И., Малхи Y, Марсено С., Mencuccini M, Müller JV, Munzinger J, Myers-Smith IH, Neill DA, Niinemets Ü, Orwin KH, Ozinga WA, Penuelas J, Pérez-Haase A , Петржик П., Филлипс О.Л., Пертель М., Райх П.Б., Рёмерманн К., Родригес А.В., Сабати ni FM, Sardans J, Schmidt M, Seidler G, Espejo JES, Silveira M, Smyth A, Sporbert M, Svenning JC, Tang Z, Thomas R, Tsiripidis I, Vassilev K, Violle C, Virtanen R, Weiher E, Welk E , Веше К., Винтер М., Вирт С., Джандт У. (2018) Глобальные взаимосвязи между характеристиками и окружающей средой растительных сообществ.Nat Ecol Evol 2: 1906. https://doi.org/10.1038/s41559-018-0699-8
И., Малхи Y, Марсено С., Mencuccini M, Müller JV, Munzinger J, Myers-Smith IH, Neill DA, Niinemets Ü, Orwin KH, Ozinga WA, Penuelas J, Pérez-Haase A , Петржик П., Филлипс О.Л., Пертель М., Райх П.Б., Рёмерманн К., Родригес А.В., Сабати ni FM, Sardans J, Schmidt M, Seidler G, Espejo JES, Silveira M, Smyth A, Sporbert M, Svenning JC, Tang Z, Thomas R, Tsiripidis I, Vassilev K, Violle C, Virtanen R, Weiher E, Welk E , Веше К., Винтер М., Вирт С., Джандт У. (2018) Глобальные взаимосвязи между характеристиками и окружающей средой растительных сообществ.Nat Ecol Evol 2: 1906. https://doi.org/10.1038/s41559-018-0699-8
Артикул
PubMed
Google ученый
Büntgen U, Bolze N, Hellmann L, Sittler B, Frauenberger B, Piermattei A, Kirdyanov A, Schweingruber FH, Ludemann T, Krusic PJ (2018) Долгосрочная динамика пополнения сообществ арктических карликовых кустарников в прибрежной восточной Гренландии . Дендрохронология 50: 70–80. https://doi.org/10.1016/j.dendro.2018.05.005
Дендрохронология 50: 70–80. https://doi.org/10.1016/j.dendro.2018.05.005
Артикул
Google ученый
Choler P (2005) Последовательные сдвиги в чертах альпийских растений по мезотопографическому градиенту. Arct Antarct Alpi Res 37: 444–453. https://doi.org/10.1657/1523-0430(2005)037[0444:CSIAPT impression2.0.CO;2
Артикул
Google ученый
Диас С., Ходжсон Дж. Г., Томпсон К., Кабидо М., Корнелиссен Дж. Х. К., Джалили А., Монтсеррат-Марти Дж., Грайм Дж. П., Зарринкамар Ф., Асри Й, Band SR, Басконсело С., Кастро-Диес П., Фунес Дж., Hamzehee B, Khoshnevi M, Pérez-Harguindeguy N, Pérez-Rontomé MC, Shirvany FA, Vendramini F, Yazdani S, Abbas-Azimi R, Bogaard A, Boustani S, Charles M, Dehghan M, de Torres-Espuny L, Falczuk , Guerrero-Campo J, Hynd A, Jones G, Kowsary E, Kazemi-Saeed F, Maestro-Martínez M, Romo-Díez A, Shaw S, Siavash B, Villar-Salvador P, Zak MR (2004). приводные экосистемы: данные с трех континентов.J Veg Sci 15: 295–304. https://doi.org/10.1111/j.1654-1103.2004.tb02266.x
приводные экосистемы: данные с трех континентов.J Veg Sci 15: 295–304. https://doi.org/10.1111/j.1654-1103.2004.tb02266.x
Артикул
Google ученый
Диас С., Каттге Дж., Корнелиссен Дж. Х. К., Райт И. Дж., Лаворель С., Дрей С., Реу Б., Клейер М., Вирт С., Колин Прентис И., Гарнье Е., Бениш Дж., Вестоби М., Портер Х, Райх П. Б., Moles AT, Dickie J, Gillison AN, Zanne AE, Chave J, Joseph Wright S, Sheremet’ev SN, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig JS, Günther A , Falczuk V, Rüger N, Mahecha MD, Gorné LD (2015) Глобальный спектр форм и функций растений.Природа 529: 167. https://doi.org/10.1038/nature16489
CAS
Статья
PubMed
Google ученый
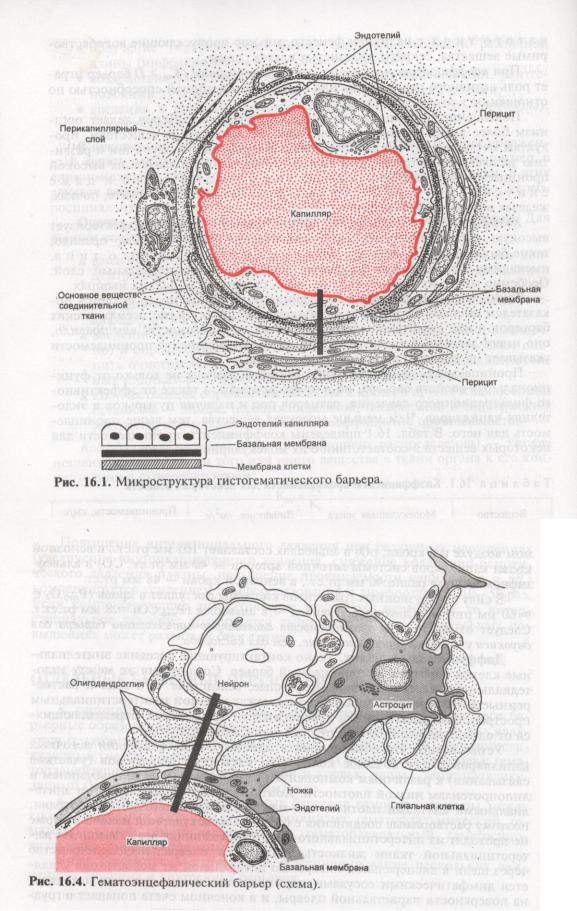 org/ScholarlyArticle»>
org/ScholarlyArticle»>Eskelinen A (2008) Воздействие травоядных и соседних животных на тундровые растения зависит от видовой принадлежности, наличия питательных веществ и местных условий окружающей среды. Дж. Экол 96: 155–165. https://doi.org/10.1111/j.1365-2745.2007.01322.x
Артикул
Google ученый
Gardner WR (1965) Динамические аспекты доступности воды и почвы для растений.Annu Rev Plant Physiol 16: 323–342. https://doi.org/10.1146/annurev.pp.16.060165.001543
CAS
Статья
Google ученый
Garnier E, Cortez J, Billès G, Navas ML, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint JP (2004) Функциональные маркеры растений фиксируют свойства экосистемы во время вторичного преемственность. Экология 85: 2630–2637. https://doi.org/10.1890/03-0799
Экология 85: 2630–2637. https://doi.org/10.1890/03-0799
Артикул
Google ученый
Kaarlejärvi E, Eskelinen A, Olofsson J (2013) Травоядность предотвращает положительную реакцию низинных растений на более теплые и более плодородные условия на больших высотах.Funct Ecol 27: 1244–1253. https://doi.org/10.1111/1365-2435.12113
Артикул
Google ученый
Lajoie G, Vellend M (2015) Понимание зависимости контекста во вкладе внутривидовой изменчивости в соответствие общности признака и окружающей среды. Экология 96: 2912–2922. https://doi.org/10.1890/15-0156.1
Артикул
PubMed
Google ученый
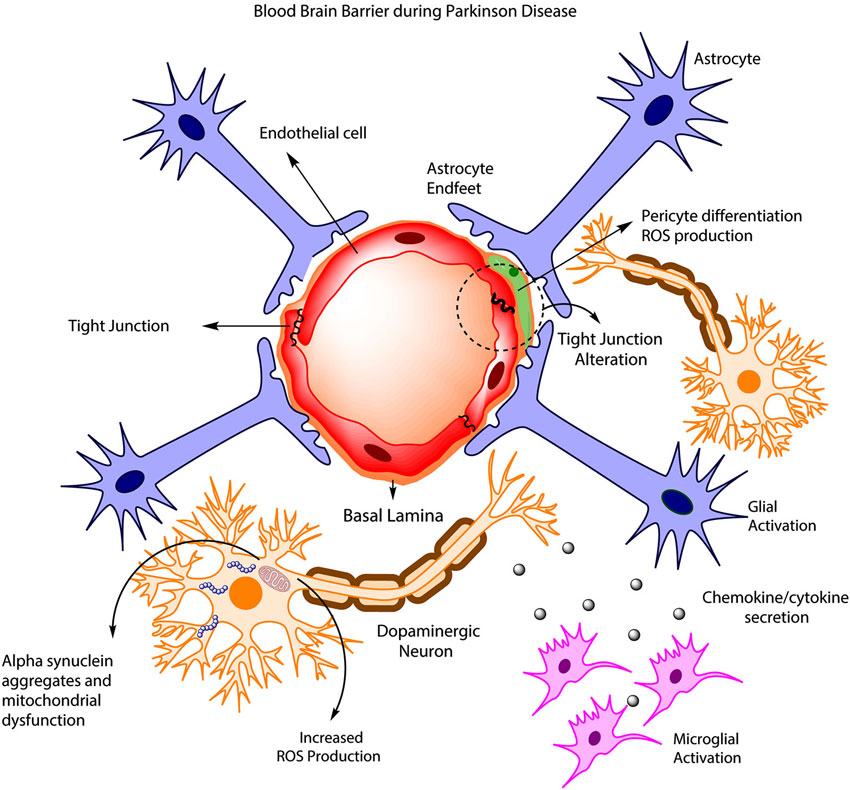 org/ScholarlyArticle»>
org/ScholarlyArticle»>Лепш Дж, де Белло Ф., Шмилауэр П., Долешал Дж. (2011) Реакция сообщества на окружающую среду: разделение круговорота видов и эффекты внутривидовой изменчивости признаков.Экография 34: 856–863. https://doi.org/10.1111/j.1600-0587.2010.06904.x
Артикул
Google ученый
Niittynen P, Heikkinen RK, Luoto M (2018) Снежный покров — это игнорируемый фактор потери арктического биоразнообразия. Nat Clim Change 8: 997. https://doi.org/10.1038/s41558-018-0311-x
Артикул
Google ученый
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2018) Vegan: сообщество экологический пакет.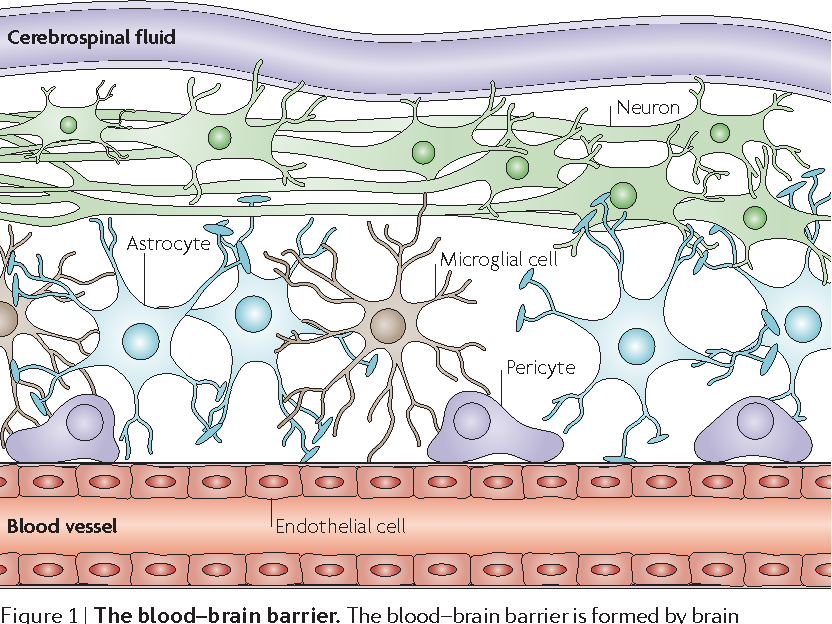 Пакет R версии 2.5-2. https://CRAN.R-project.org/package=vegan
Пакет R версии 2.5-2. https://CRAN.R-project.org/package=vegan
Паюнен А., Виртанен Р., Ройнинен Х. (2008) Влияние выпаса оленей на состав и видовое богатство растительности в экотоне лесотундры. Полярная биол 31: 1233–1244. https://doi.org/10.1007/s00300-008-0462-8
Артикул
Google ученый
Перес-Рамос И.М., Румет С., Круз П., Бланшар А., Отран П., Гарнье Э. (2012) Доказательства «экономического спектра растительных сообществ», обусловленного ограничениями питательных веществ и воды в средиземноморских пастбищах на юге Франции.J Ecol 100: 1315–1327. https://doi.org/10.1111/1365-2745.12000
Артикул
Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Пиринен П., Симола Х., Аалто Дж., Каукоранта Дж., Карлссон П., Руухела Р. (2012) Климатологическая статистика Финляндии за 1981–2010 годы. Finn Meteorol Inst Rep 1:83
Google ученый
Post E, Forchhammer MC, Bret-Harte MS, Callaghan TV, Christensen TR, Elberling B, Fox AD, Gilg O, Hik DS, Høye TT, Ims RA, Jeppesen E, Klein DR, Madsen J, McGuire AD , Rysgaard S, Schindler DE, Stirling I, Tamstorf MP, Tyler NJC, van der Wal R, Welker J, Wookey PA, Schmidt NM, Aastrup P (2009) Экологическая динамика 388 в Арктике, связанная с недавним изменением климата.Science 325: 1355–1358. https://doi.org/10.1126/science.1173113
CAS
Статья
Google ученый
R Core Team (2018) R: язык и среда для статистических вычислений.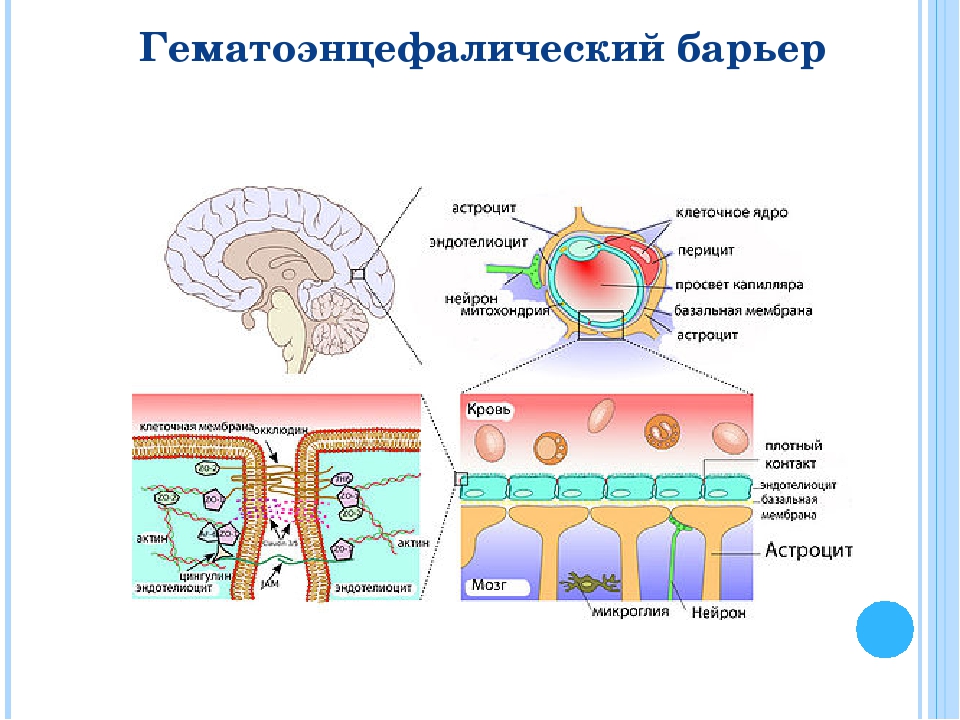 R Foundation for Statistical Computing, Вена
R Foundation for Statistical Computing, Вена
Schindelin J, Arganda-Carreras I, Frize E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Элисири К., Томанчак П., Кардона А. (2012) Фиджи: платформа с открытым исходным кодом для анализа биологических изображений.Нат Методы 9: 676–682. https://doi.org/10.1038/nmeth.2019
CAS
Статья
PubMed
PubMed Central
Google ученый
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 лет анализа изображений. Nat Методы 9: 671–675. https://doi.org/10.1038/nmeth.2089
CAS
Статья
PubMed
PubMed Central
Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Siefert A, Violle C, Chalmandrier L, Albert CH, Taudiere A, Fajardo A, Aarssen LW, Baraloto C, Carlucci MB, Cianciaruso MV, de L.Dantas V, de Bello F, Duarte LDS, Fonseca CR, Freschet GT, Gaucherand S, Gross N, Hikosaka K, Jackson B, Jung V, Kamiyama C, Katabuchi M, Kembel SW, Kichenin E, Kraft NJB, Lagerström A, Bagousse -Пингет Ю.Л., Ли Й., Мейсон Н., Мессье Дж., Накашизука Т., Овертон Дж. М., Пельцер Д. А., Перес-Рамос И. М., Пиллар В. Д., Прентис Х. С., Ричардсон С., Сасаки Т., Шамп Б. С., Шёб С., Шипли Б., Сундквист М. , Sykes MT, Vandewalle M, Wardle DA (2015) Глобальный метаанализ относительной степени внутривидовой изменчивости признаков в растительных сообществах.Ecol Lett 18: 1406–1419. https://doi.org/10.1111/ele.12508
Артикул
PubMed
Google ученый
Спасоевич М.Дж., Судинг К.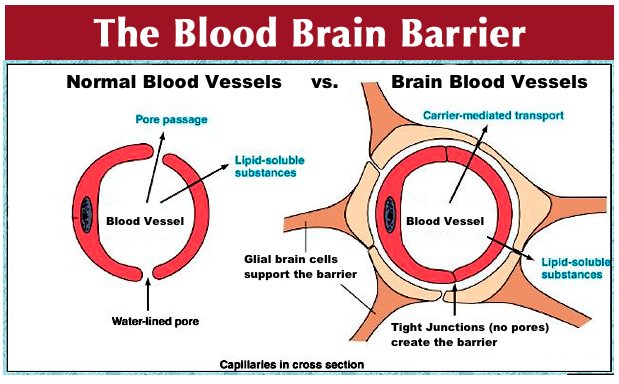 Н. (2012) Вывод механизмов сборки сообщества из моделей функционального разнообразия: важность множественных процессов сборки. J Ecol 100: 652–661. https://doi.org/10.1111/j.1365-2745.2011.01945.x
Н. (2012) Вывод механизмов сборки сообщества из моделей функционального разнообразия: важность множественных процессов сборки. J Ecol 100: 652–661. https://doi.org/10.1111/j.1365-2745.2011.01945.x
Артикул
Google ученый
Тилман Д. (1988) Стратегии растений, динамика и структура растительных сообществ.Princeton University Press, Princeton (ISBN: 978-0-691-08489-3)
Google ученый
Венн С.Е., Грин К., Пикеринг С.М., Морган Дж.В. (2011) Использование функциональных характеристик растений для объяснения состава сообщества через сильный фильтр окружающей среды в австралийских альпийских снежных пятнах. Завод Ecol 212: 1491–1499. https://doi.org/10. 1007/s11258-011-9923-1
1007/s11258-011-9923-1
Артикул
Google ученый
Виртанен Р., Оксанен Л., Оксанен Т., Коэн Дж., Форбс Б.К., Йохансен Б., Кайхкё Дж., Олофссон Дж., Пуллиайнен Дж., Тёммервик Х. (2016) Где безлесные тундровые районы северных высокогорья вписываются в глобальный биом система: к экологически естественному подразделению тундрового биома.Ecol Evolut 6: 143–158. https://doi.org/10.1002/ece3.1837
Артикул
Google ученый
Wood SN (2011) Быстрая стабильная оценка максимального и предельного правдоподобия полупараметрических обобщенных линейных моделей. J R Stat Soc (B) 73: 3–36. https://doi.org/10.1111/j.1467-9868.2010.00749.x
Артикул
Google ученый
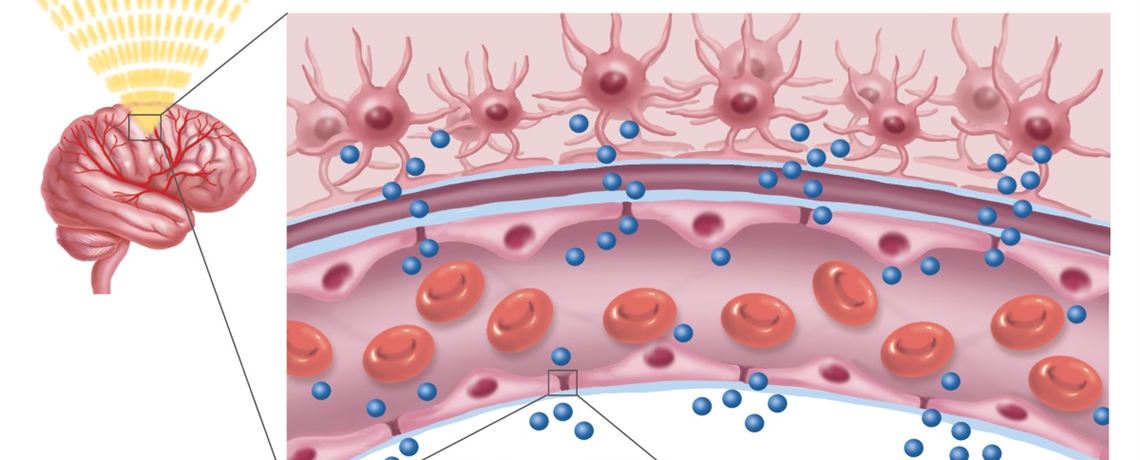 org/ScholarlyArticle»>
org/ScholarlyArticle»>Райт И.Дж., Рейх П.Б., Вестоби М., Акерли Д.Д., Барух З., Бонжерс Ф., Кавендер-Барес Дж., Чапин Т., Корнелиссен Дж.Х.С., Димер М., Флексас Дж., Гарнье Е., Грум П.К., Гулиас Дж., Хикосака К. , Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) Спектр экономики листа во всем мире.Природа 428: 821. https://doi.org/10.1038/nature02403
CAS
Статья
Google ученый
geb / 030-navigator.adoc at master · geb / geb · GitHub
geb / 030-navigator.adoc at master · geb / geb · GitHub
Постоянная ссылка
В настоящее время невозможно получить участников
1115 строк (792 слока)
38.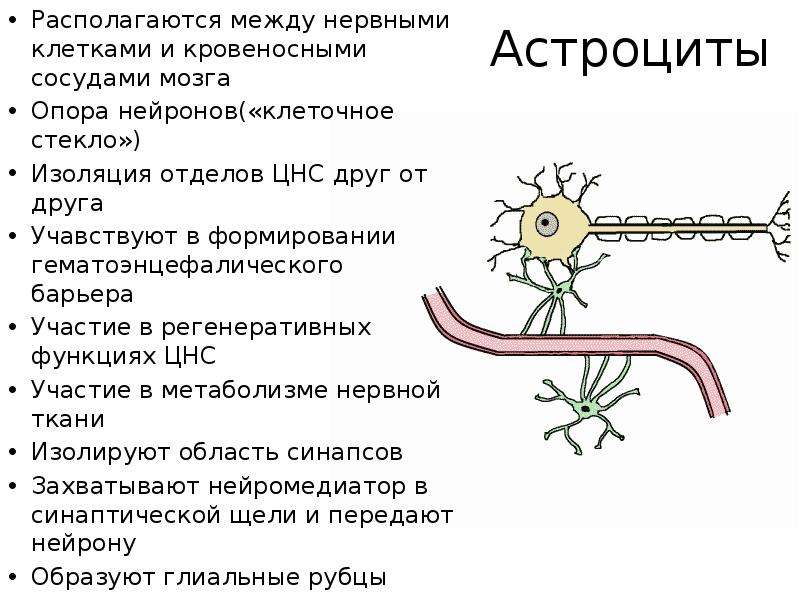 9 КБ
9 КБ
Geb предоставляет краткий и удобный интерфейс для содержимого и элементов управления в вашем браузере.
Это реализовано с помощью API-интерфейса Navigator , который представляет собой вдохновленный jQuery механизм для поиска, фильтрации и взаимодействия с элементами DOM.
Функция
$ ()
Функция $ () — это точка доступа к содержимому страницы браузера.
Он возвращает объект {navigator-api} , который примерно аналогичен объекту jQuery.Он аналогичен тому, что представляет один или несколько элементов на странице и может использоваться для уточнения сопоставленного содержимого или запроса сопоставленного содержимого.
Когда вызывается функция $ () , которая не соответствует никакому содержимому, возвращается «пустой» объект навигатора, не представляющий содержимого.
Операции с «пустыми» навигаторами возвращают null или другой «пустой» навигатор или другие значения, которые имеют смысл (например, метод size () возвращает 0 ).
Сигнатура функции $ () выглядит следующим образом…
$ («селектор css», «индекс или диапазон», «сопоставители атрибутов / текста»)
Ниже приводится конкретный пример…
ссылка: {test-dir} / navigator / DollarExamplesSpec.Groovy [] Будет найден 3-й (элементы с индексом 0) h2 элемент, чей атрибут class в точности соответствует « заголовок ».
Все аргументы являются необязательными, что означает, что все следующие вызовы действительны:
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Примечание | Существует псевдоним для функции доллара с именем «find», если метод с именем «$» вам не по вкусу. |
Селекторы CSS
Вы можете использовать любой селектор CSS, который поддерживает базовая реализация WebDriver …
ссылка: {test-dir} /navigator/DollarExamplesSpec. groovy []  groovy []
groovy [] Использование селекторов классов WebDriver
By
Для всех сигнатур функции $ () , которые принимают селектор css, существует эквивалентная сигнатура, в которой экземпляр класса {by-api} WebDriver может использоваться вместо строки .
Использование селекторов CSS - это идиоматический способ использования Geb, и его следует предпочесть использованию селекторов By .
Всегда можно выбрать те же элементы, используя селектор css, как и при использовании определенного селектора By , кроме определенных селекторов XPath, поэтому предоставляется этот удобный механизм.
Ниже приведены некоторые примеры использования селекторов By …
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Динамические навигаторы
По умолчанию, когда создается экземпляр Navigator , сохраняется постоянная ссылка на элементы, соответствующие селектору в этот момент времени. Во многих ситуациях это желаемое поведение, поскольку оно ограничивает количество команд WebDriver, выполняемых для поиска элементов, но во времена современных фреймворков javascript и одностраничных приложений это могло быть не так.
Во многих ситуациях это желаемое поведение, поскольку оно ограничивает количество команд WebDriver, выполняемых для поиска элементов, но во времена современных фреймворков javascript и одностраничных приложений это могло быть не так.
Это особенно актуально при работе с приложениями, которые полностью перерисовывают части DOM при изменении модели, а не обновляют ее.
Довольно часто тот факт, что элементы DOM, поддерживающие Navigator , были удалены из DOM и перерисованы, можно обойти, создав новый экземпляр этого Navigator , получив его из определения содержимого.К сожалению, этот обходной путь недоступен, когда действие, выполняемое как часть метода, определенного в классе модуля Geb, вызывает повторную визуализацию базового элемента модуля.
Это связано с тем, что ссылка на базовый Navigator модуля фиксируется во время создания модуля, что, в свою очередь, фиксирует ссылку на элементы, соответствующие селектору, используемому в определении этого Navigator .
Когда воссоздается часть DOM, которая содержит элементы этого Navigator , эти ссылки становятся устаревшими, в результате чего любые вызываемые ими методы генерируют {stale-element-reference-exception-api} , и их устаревание не может быть исправлено изнутри. код рассматриваемого метода модуля.Решение этой проблемы состоит в том, чтобы объявить базовый Navigator динамическим, что означает, что список элементов DOM, лежащих в основе этого Navigator , будет обновляться каждый раз, когда на нем вызывается метод доступа к элементам.
Любой метод, который возвращает Navigator и принимает карту атрибутов в качестве одного из параметров (большинство из этих методов описаны в разделах «Поиск и фильтрация» и «Перемещение»), можно использовать для создания динамического Navigator , просто передав {groovy -truth-link} [истинное] значение как динамический атрибут …
ссылка: {test-dir} / navigator / DynamicNavigatorSpec. Groovy []  Groovy []
Groovy [] Предупреждение | Вы должны иметь в виду, что динамические |
Давайте рассмотрим простое приложение Vue.js, которое отображает список элементов, которые можно изменить, используя следующий шаблон…
ссылка: {real-browser-test-dir} / navigator / DynamicModuleBaseSpec. Groovy [] 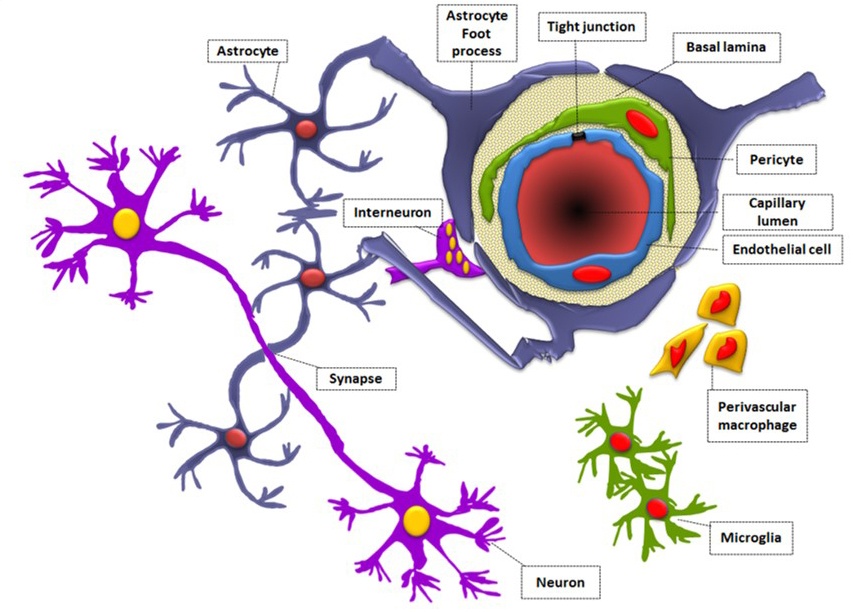 Groovy []
Groovy [] … и следующий код javascript…
ссылка: {real-browser-test-dir} /navigator/DynamicModuleBaseSpec.groovy [] Важно отметить, что элементы li повторно визуализируются при изменении порядка.
Давайте смоделируем каждый элемент в списке, используя следующий модуль…
ссылка: {real-browser-test-dir} /navigator/DynamicModuleBaseSpec.groovy [] Учитывая следующее определение содержания…
ссылка: {real-browser-test-dir} / navigator / DynamicModuleBaseSpec.Groovy [] Приведенный ниже код завершится ошибкой с StaleElementReferenceException …
ссылка: {real-browser-test-dir} /navigator/DynamicModuleBaseSpec.groovy [] Если мы изменим базовый навигатор модуля элемента списка на динамический, как в следующем классе страницы…
ссылка: {real-browser-test-dir} /navigator/DynamicModuleBaseSpec.groovy [] Тогда пройдет следующее .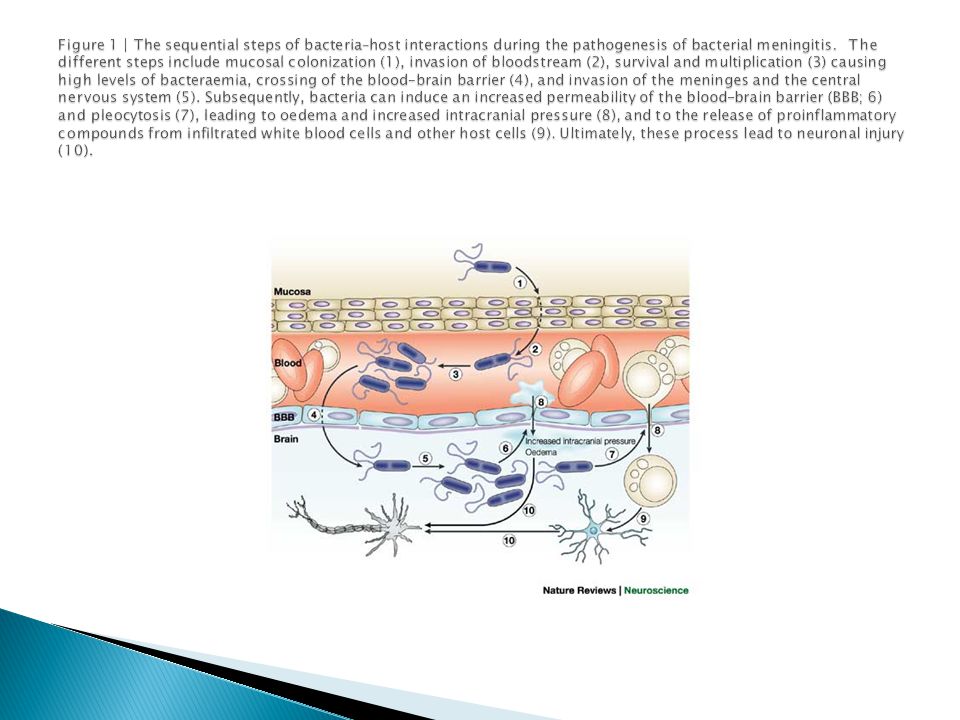 ..
..
ссылка: {real-browser-test-dir} / navigator / DynamicModuleBaseSpec.Groovy [] Предупреждение | Хотя использование динамических навигаторов может показаться способом полностью избежать |

Индексы и диапазоны
При сопоставлении можно указать одно положительное целое число или диапазон целых чисел для ограничения по индексу.
Рассмотрим следующий html…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Мы можем использовать индексы для сопоставления контента вот так…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] См. Ниже объяснение метода text () и использования оператора распространения.
Соответствие атрибутов и текста
Можно выполнять сопоставления атрибутов и текстовых значений узлов с помощью синтаксиса именованных параметров Groovy.Значение , текст обрабатывается специально как совпадение с текстом узла.
Все остальные значения сравниваются с соответствующими значениями атрибутов.
Рассмотрим следующий html…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Мы можем использовать сопоставители атрибутов вот так…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Значения атрибутов объединяются вместе…
ссылка: {test-dir} / navigator / DollarExamplesSpec.Groovy [] Мы можем использовать сопоставители текста вот так…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Вы можете смешивать атрибуты и сопоставления текста…
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] Использование выкроек
Чтобы сопоставить все значение атрибута или текст, используйте значение String . Также можно использовать шаблон для сопоставления регулярных выражений…
ссылка: {test-dir} / navigator / DollarExamplesSpec.Groovy [] Geb также поставляется с набором методов быстрого доступа…
ссылка: {test-dir} /navigator/DollarExamplesSpec. groovy []  groovy []
groovy [] Ниже приводится полный список:
| С учетом регистра | Нечувствительный к корпусу | Описание |
|---|---|---|
| | Соответствует значениям, начинающимся с заданного значения |
| | Соответствует значениям, которые содержат данное значение в любом месте |
| | Соответствует значениям, которые заканчиваются заданным значением |
| | Соответствует значениям, которые содержат заданное значение, окруженное либо пробелом, либо началом или концом значения |
| | Соответствует значениям, которые НЕ начинаются с заданного значения |
| | Соответствует значениям, которые нигде НЕ содержат данное значение |
| | Соответствует значениям, которые НЕ заканчиваются заданным значением |
| | Соответствует значениям, которые НЕ содержат заданное значение, окруженное либо пробелом, либо началом или концом значения |
Все эти методы сами по себе могут принимать строку или шаблон …
ссылка: {test-dir} / navigator / DollarExamplesSpec. Groovy []  Groovy []
Groovy [] Примечание | Вам может быть интересно, как работает эта магия, то есть откуда эти методы и где их можно использовать. |
Использование более одного шаблона
Иногда возникает необходимость поиска альтернативных шаблонов, например, если у вас есть мультибрендовая страница с разным текстом для каждой марки,
в этом случае вы можете использовать составные сопоставители allOf и anyOf .Оба принимают любые комбинации других сопоставителей.
ссылка: {test-dir} /navigator/DollarExamplesSpec.groovy [] | Составной сопоставитель | Описание |
|---|---|
| Совпадает, если все указанные сопоставители совпадают |
| Совпадает, если какой-либо из заданных совпадений соответствует |
Навигаторы повторяются
Объекты навигатора реализуют интерфейс Java Iterable , который позволяет вам делать множество вещей Groovy, например, использовать функцию max () …
Рассмотрим следующий html…
ссылка: {test-dir} / navigator / IterableSpec. Groovy []  Groovy []
Groovy [] Функцию max () можно использовать на экземплярах Navigator …
ссылка: {test-dir} /navigator/IterableSpec.groovy [] Это также означает, что объекты навигатора работают с оператором распространения Groovy…
ссылка: {test-dir} /navigator/IterableSpec.groovy [] При обработке навигатора как Iterable повторяемый контент всегда является точно совпадающими элементами (в отличие от включения дочерних элементов).
Прослушивание событий навигатора
Можно зарегистрировать слушателя, который будет получать уведомления каждый раз, когда происходят определенные события Navigator.
Лучшим справочником по событиям, которые можно прослушивать, является {navigator-event-listener-api} [документация для интерфейса NavigatorEventListener ].
Если вы хотите прослушивать только подмножество событий навигатора, то {navigator-event-listener-support-api} может пригодиться, поскольку он поставляется с пустыми реализациями по умолчанию всех методов {navigator-event-listener-api } [ NavigatorEventListener ].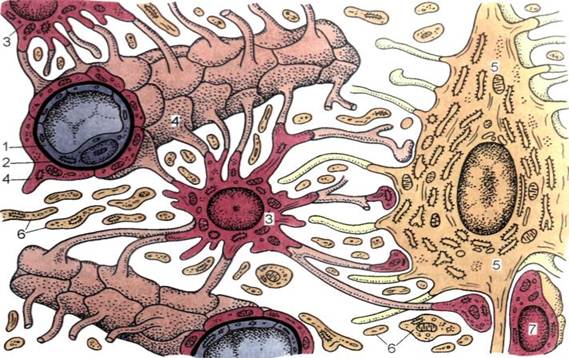
Одним из вариантов использования прослушивателя событий навигатора было бы улучшение отчетов путем написания отчета каждый раз, когда щелкают навигатор, его значение изменяется и т. Д.
В приведенном ниже примере показано, как зарегистрировать прослушиватель событий навигатора как часть сценария конфигурации, который просто печатает теги навигатора после нажатия на навигатор …
ссылка: {test-dir} /navigator/NavigatorEventListenerSpec.groovy [] равно () и hashCode ()
Можно проверить на равенство Navigator экземпляров.Правила просты — два пустых навигатора всегда равны, а два непустых навигатора равны только в том случае, если они содержат одни и те же элементы в одном и том же порядке.
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/EqualsSpec.groovy [] Вот несколько примеров одинаковых экземпляров навигатора…
ссылка: {test-dir} /navigator/EqualsSpec. groovy [] 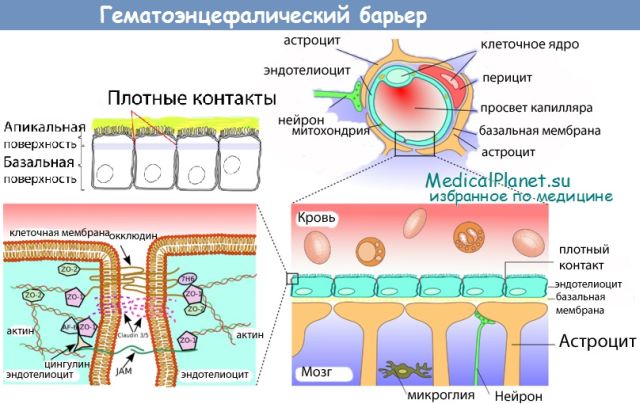 groovy []
groovy [] Два пустых штурмана
Два одноэлементных навигатора, содержащие один и тот же элемент
Два одноэлементных навигатора, содержащих один и тот же элемент, созданный разными методами
Два многоэлементных навигатора, содержащих одинаковые элементы
Два многоэлементных навигатора, содержащих одинаковые элементы, созданные разными методами
И не равные…
ссылка: {test-dir} / navigator / EqualsSpec.Groovy [] Пустые и непустые навигаторы
Одноэлементные навигаторы, содержащие разные элементы
Многоэлементные навигаторы, содержащие те же элементы, но в другом порядке
Поиск и фильтрация
Объекты
Navigator имеют методы find () и $ () для поиска потомков и методы filter () и not () для сокращения совпадающего содержимого.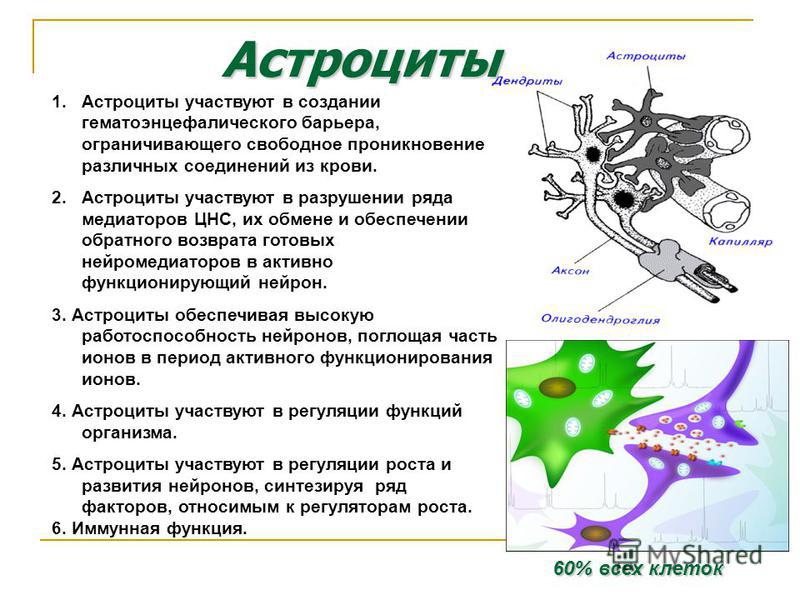
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy []
ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] Можно выбрать дел. B по…
ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] Мы можем выбрать div , содержащий p с…
ссылка: {test-dir} / navigator / FindingAndFilteringSpec.Groovy [] Или выберите div , содержащий input с атрибутом типа «текст», например…
ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] Мы можем выбрать div , который не содержит p , с помощью…
ссылка: {test-dir} /navigator/FindingAndFilteringSpec. groovy []  groovy []
groovy [] Или выберите div , который не содержит input с атрибутом типа «текст», например…
ссылка: {test-dir} / navigator / FindingAndFilteringSpec.Groovy [] Или выберите два div , которые не содержат input с атрибутом типа «submit», например…
ссылка: {test-dir} /navigator/FindingAndFilteringSpec.groovy [] Метод find () поддерживает те же типы аргументов , что и функция $ () .
Фильтр () , not () , имеет () и hasNot () методы имеют одинаковые сигнатуры — они принимают: строку селектора, карту предикатов или и то, и другое.
Эти методы возвращают новый объект навигатора, представляющий новое содержимое.
Композиция
Также возможно составить объекты навигатора из других объектов навигатора в ситуациях, когда вы не можете выразить набор содержимого в одном запросе.
Для этого просто вызовите функцию $ () и передайте навигаторы для создания.
Рассмотрим следующую разметку…
ссылка: {test-dir} /navigator/CompositionSpec.groovy [] Затем вы можете создать новый объект навигатора, который представляет абзацы a и b следующим образом:
ссылка: {test-dir} / navigator / CompositionSpec.Groovy [] Альтернативный способ — использовать метод add () из Navigator , который принимает либо String , либо селектор Webdriver’s By :
ссылка: {test-dir} /navigator/CompositionSpec.groovy [] Наконец, вы можете составлять объекты навигатора из содержимого. Итак, учитывая определение содержимого страницы:
ссылка: {test-dir} /navigator/CompositionSpec.groovy [] Вы можете объединить элементы содержимого в навигатор следующим образом:
ссылка: {test-dir} / navigator / CompositionSpec. Groovy []  Groovy []
Groovy [] Прохождение
Навигаторы также имеют методы для выбора контента вокруг совпадающего контента.
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Вы можете выбрать контент около p.d с помощью…
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Рассмотрим следующий HTML…
ссылка: {test-dir} / navigator / TraversingSpec.Groovy [] Следующий код выберет p.b и p.c …
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] предыдущий () , prevAll () , следующий () , nextAll () , родительский () , родительский () , ближайший () , братья и сестры () и Методы children () также могут принимать селекторы CSS и сопоставители атрибутов.
Используя тот же HTML, в следующих примерах будет выбрано p.c …
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Аналогичным образом рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] В следующих примерах будет выбрано дел. B …
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Метод closest () является особым случаем, поскольку он выбирает первого предка текущих элементов, который соответствует селектору.
Не существует версии метода closest () без аргументов.Например, будет выбрано дел. A …
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Эти методы не принимают индексы, поскольку они автоматически выбирают первое совпадающее содержимое.
Чтобы выбрать несколько элементов, вы можете использовать prevAll () , nextAll () и parent () , все из которых имеют версии без аргументов и версии, которые фильтруются с помощью селектора.
Методы nextUntil () , prevUntil () и parentUntil () возвращают все узлы вдоль соответствующей оси до первого, который соответствует селектору или атрибутам.Рассмотрим следующую разметку:
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] В следующих примерах будет выбрано div.b и div.c :
ссылка: {test-dir} /navigator/TraversingSpec.groovy [] Нажатие
Объекты
Navigator реализуют метод click () , который дает указание браузеру щелкнуть единственный элемент, сопоставленный навигатором.
Если этот метод вызывается для мультиэлемента Navigator , то создается исключение SingleElementNavigatorOnlyMethodException .
ссылка: {test-dir} /navigator/ClickingSpec.groovy [] Щелкнет элемент a.login , затем вызовет browser.page (LoginPage) и убедитесь, что браузер находится на ожидаемой странице.
Для всех классов страниц, переданных при использовании варианта списка, должна быть определена функция проверки «at», в противном случае будет выброшено исключение UndefinedAtCheckerException .
Определение видимости
Объекты навигатора имеют свойство отображено , которое указывает, виден ли элемент пользователю или нет.Отображаемое свойство объекта навигатора, которое ни с чем не соответствует, всегда является ложным .
Фокус
Geb предлагает простой способ поиска текущего элемента в виде метода Focus () , который возвращает Navigator .
Также можно проверить, удерживает ли данный объект Navigator текущий элемент с фокусом, используя его свойство с фокусом . с фокусом
Объекты Navigator , которые не соответствуют ни одному элементу, вернут false как значение свойства , а объекты, соответствующие нескольким элементам, вызовут исключение SingleElementNavigatorOnlyMethodException .
Учитывая следующий HTML…
ссылка: {test-dir} /navigator/FocusSpec.groovy [] focus () и свойство Navigator focus можно использовать следующим образом…
ссылка: {test-dir} /navigator/FocusSpec.groovy [] Размер и расположение
Вы можете получить размер и расположение содержимого на странице.
Все единицы указаны в пикселях.
Размер доступен через свойства height и width , а расположение доступно как свойства x и y , которые представляют расстояние от верхнего левого угла страницы.
(или родительский фрейм) до верхней левой точки содержимого.
Все эти свойства работают только с совпадающим элементом sole , а SingleElementNavigatorOnlyMethodException генерируется, если к ним обращаются на мультиэлементе Navigator .
Рассмотрим следующий html…
ссылка: {test-dir} /navigator/SizeAndLocationSpec.groovy [] Для него выполняются следующие условия…
ссылка: {test-dir} /navigator/SizeAndLocationSpec.groovy [] Чтобы получить какие-либо свойства для всех согласованных элементов, можно использовать оператор распространения Groovy.
ссылка: {test-dir} /navigator/SizeAndLocationSpec.groovy [] Доступ к имени тега, атрибутам, тексту и классам
Методы tag () , text () и classes () , а также доступ к атрибутам через нотацию @ или метод attr () возвращают запрошенное содержимое на единственном совпавшем элементе .
Если эти методы вызываются для мультиэлемента Navigator , то создается исключение SingleElementNavigatorOnlyMethodException .Метод classes () возвращает java.util.List имен классов, отсортированных по алфавиту.
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/TagTextClassesAttributesSpec.groovy [] Справедливы следующие утверждения…
ссылка: {test-dir} /navigator/TagTextClassesAttributesSpec.groovy [] Для получения информации обо всем совпадающем содержимом используйте оператор распространения Groovy …
ссылка: {test-dir} / navigator / TagTextClassesAttributesSpec.Groovy [] Свойства CSS
Доступ к
Css свойствам одноэлементного навигатора можно получить с помощью метода css () .
Если этот метод вызывается для мультиэлемента Navigator , то создается исключение SingleElementNavigatorOnlyMethodException .
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/CssPropertiesSpec.groovy [] Вы можете получить значение свойства float css следующим образом…
ссылка: {test-dir} / navigator / CssPropertiesSpec.Groovy [] Предупреждение | Существуют некоторые ограничения при получении свойств css объектов |
Отправка нажатий клавиш
Учитывая следующий HTML…
ссылка: {test-dir} /navigator/KeystrokesSpec.groovy [] Вы можете отправлять нажатия клавиш на ввод (и любое другое содержимое) с помощью оператора leftShift, который является ярлыком для метода {webdriver-send-keys-api} WebDriver.
ссылка: {test-dir} /navigator/KeystrokesSpec.groovy [] То, как контент реагирует на нажатия клавиш, зависит от того, что это за контент.
Не символы (например, клавиша удаления, ключевые аккорды и т. Д.)
Можно отправлять в контент нетекстовые символы с помощью перечисления WebDriver {webdriver-keys-api} …
ссылка: {real-browser-test-dir} /navigator/NonCharacterKeystrokesSpec.groovy []
ссылка: {real-browser-test-dir} /navigator/NonCharacterKeystrokesSpec.groovy [] Здесь мы отправляем «control-c» на вход.
См. Документацию по перечислению {webdriver-keys-api} для получения дополнительной информации о возможных ключах.
1. Доступ к входным значениям
Значение input , select и textarea Элементы могут быть получены и установлены с помощью метода value .
Вызов value () без аргументов вернет строковое значение , единственный элемент в навигаторе.
Если этот метод вызывается для мультиэлемента Navigator , то создается исключение SingleElementNavigatorOnlyMethodException .
Вызов value (value) установит текущее значение для всех элементов в навигаторе.Аргумент может быть любого типа и при необходимости будет преобразован в строку .
Исключениями являются то, что при установке значения флажка метод ожидает логическое значение (или существующее значение флажка или метку), а при установке нескольких значений выберите метод ожидает массив или
Коллекция ценностей.
2. Ярлыки управления формой
Взаимодействие с элементами управления формы ( ввод , выберите и т. Д.) - настолько распространенная задача в веб-функциональном тестировании, что Geb предоставляет удобные ярлыки для общих функций.
Geb поддерживает следующие сочетания клавиш для работы с элементами управления формы.
Рассмотрим следующий HTML…
ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] Значение может быть прочитано и записано с помощью обозначения свойств…
ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] Это буквально ярлыки для…
ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] Также есть ярлык для получения навигатора по имени элемента управления…
ссылка: {test-dir} / navigator / FormShortcutsSpec.Groovy [] Примечание | В приведенных выше и ниже примерах с элементами управления формой мы используем такой код, как |
Если ваше определение содержимого (страница или модуль) описывает содержимое, которое является входом , select или textarea , вы можете получить доступ и установить его значение так же, как описано выше для форм.Учитывая определения страницы и модуля для вышеупомянутого HTML:
ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] ссылка: {test-dir} /navigator/FormShortcutsSpec.groovy [] 3. Установка контрольных значений формы
Примечание | В следующих примерах описывается использование элементов управления формы только с использованием кода типа |
Предупреждение | Попытка установить значение для элемента, который не является одним из |
3.1. Выберите
Выбор значений устанавливается путем присвоения значения или текста требуемой опции. Присвоенные значения автоматически переводятся в String. Например…
ссылка: {test-dir} /navigator/SelectSpec.groovy [] Мы можем выбрать варианты с…
ссылка: {test-dir} /navigator/SelectSpec.groovy [] Первый вариант, выбранный его атрибутом значения.
Второй вариант, выбранный его атрибутом значения с приведением аргументов.
Третий вариант, выбранный по его тексту.
Если вы попытаетесь установить для выбора значение, которое не соответствует значению или тексту каких-либо параметров, будет выброшено исключение IllegalArgumentException .
3.2. Множественный выбор
Если для выбора есть атрибут , несколько , он устанавливается с помощью массива или Коллекция значений.
Все параметры, которых нет в значениях, не выбираются.
Например…
ссылка: {test-dir} / navigator / MultiSelectSpec.Groovy [] Мы можем выбрать варианты с…
ссылка: {test-dir} /navigator/MultiSelectSpec.groovy [] Второй и третий варианты выбираются по их атрибутам значения.
Первый, четвертый и пятый варианты, выбираемые по их атрибутам значения с приведением аргументов.
Первый и последний варианты, выбранные по их тексту.
Все параметры не выбраны.
Если назначаемая коллекция содержит значение, которое не соответствует значению или тексту каких-либо параметров, будет выброшено исключение IllegalArgumentException .
3.3. Флажок
Чекбоксы обычно устанавливаются / снимаются, устанавливая для них значение true или false .
Рассмотрим следующий html…
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] Вы можете установить отдельный флажок следующим образом…
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] Вызов value () для установленного флажка вернет значение атрибута value , т.е.e:
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] Вызов value () при снятом флажке вернет null , то есть:
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] В общем, вы должны использовать {groovy-true} при проверке, установлен ли флажок:
ссылка: {test-dir} / navigator / CheckboxSpec.Groovy [] ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] 3.4. Несколько флажков
Вы также можете установить флажок, явно установив для него значение или используя его метку. Это полезно, когда у вас есть несколько флажков с одинаковыми именами, например:
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] Вы можете выбрать собаку в качестве типа питомца следующим образом:
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] Если вы хотите установить несколько флажков вместо одного, вы можете использовать коллекцию в качестве значения:
ссылка: {test-dir} / navigator / CheckboxSpec.Groovy [] При проверке наличия флажка и наличия нескольких флажков с одинаковым именем убедитесь, что вы используете навигатор, который содержит только один из них, прежде чем вызывать на нем value () :
ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] ссылка: {test-dir} /navigator/CheckboxSpec.groovy [] 3,5. Радио
Значения радио устанавливаются назначением значения переключателя, который должен быть выбран, или текста метки, связанного с переключателем.
Например, со следующими переключателями…
ссылка: {test-dir} /navigator/RadioSpec.groovy [] Мы можем выбрать радиостанции по значению…
ссылка: {test-dir} /navigator/RadioSpec.groovy [] ссылка: {test-dir} /navigator/RadioSpec.groovy [] 3,6. Ввод текста и текстовые поля
В случае ввода текста или текстового поля присвоенное значение становится атрибутом value элемента.
ссылка: {test-dir} /navigator/TextInputAndAreaSpec.groovy [] ссылка: {test-dir} /navigator/TextInputAndAreaSpec.groovy [] Также возможно добавить текст, используя сокращение клавиш отправки…
ссылка: {test-dir} /navigator/TextInputAndAreaSpec.groovy [] Который также может использоваться для несимвольных клавиш…
ссылка: {real-browser-test-dir} /navigator/BackspaceSpec.groovy []
ссылка: {real-browser-test-dir} / navigator / BackspaceSpec.Groovy [] Предупреждение | WebDriver имеет некоторые проблемы с текстовыми полями и окружающими пробелами. |
3,7. Загрузка файла
В настоящее время с помощью WebDriver невозможно имитировать процесс, когда пользователь нажимает на элемент управления загрузкой файлов и выбирает файл для загрузки с помощью обычного средства выбора файлов.Однако вы можете напрямую установить значение элемента управления загрузкой на абсолютный путь к файлу в системе, в которой работает драйвер, и при отправке формы этот файл будет загружен.
Итак, если ваш HTML-код выглядит как…
ссылка: {test-dir} /navigator/FileUploadSpec.groovy [] А переменная uploadedFile содержит экземпляр File , указывающий на файл, который вы хотите выгрузить, вот как вы можете установить значение элемента управления выгрузкой…
ссылка: {test-dir} / navigator / FileUploadSpec.Groovy [] 4. Сложные взаимодействия
WebDriver поддерживает более сложные взаимодействия, чем простой щелчок или ввод элементов, например перетаскивание.
Вы можете использовать этот API напрямую при использовании Geb или использовать более удобный для Geb interact DSL.
4.1. Использование WebDriver
Actions API напрямую
Объект навигатора Geb построен на основе коллекции объектов WebDriver {webelement-api}.
Доступ к содержащимся в нем экземплярам WebElement можно получить с помощью следующих методов для объектов навигатора:
ссылка: {core-source-dir} / geb / navigator / Navigator.Groovy [] Используя методы класса WebDriver {actions-api} с экземплярами WebElement , можно эмулировать сложные пользовательские жесты.
Сначала вам нужно будет создать экземпляр Actions после получения драйвера WebDriver:
ссылка: {real-browser-test-dir} /navigator/InteractionsSpec.groovy [] Затем используйте методы Actions , чтобы составить серию действий пользовательского интерфейса, затем вызовите build () , чтобы создать конкретное действие Action :
ссылка: {real-browser-test-dir} / navigator / InteractionsSpec.Groovy [] Наконец, вызовите perform () , чтобы активировать желаемое поведение мыши или клавиатуры:
ссылка: {real-browser-test-dir} /navigator/InteractionsSpec.groovy [] 4.2. Использование
interact ()
Чтобы избежать необходимости управлять сборкой и выполнением Action через жизненный цикл экземпляра Actions , а также необходимости получать экземпляры WebElement из Navigator при эмуляции пользователя
жестов, Геб добавляет метод interact () .При использовании этого метода экземпляр Actions неявно создается, встроен в действие Action и выполняется.
Делегат закрытия, переданный в interact () , является экземпляром {interact-delegate-api} , который объявляет те же методы, что и Actions , но принимает Navigator в качестве аргументов для методов Actions
которые принимают WebElement .
ссылка: {real-browser-test-dir} / navigator / InteractionsSpec.Groovy [] Хотя обычно это не требуется, можно получить прямой доступ к экземпляру Actions , поддерживающему InteractDelegate :
ссылка: {real-browser-test-dir} /navigator/InteractionsSpec.groovy [] Полный список доступных методов, которые можно использовать для эмуляции жестов пользователя, см. В документации по классу {interact-delegate-api} .
4.3. Примеры взаимодействия
Вызовы на interact () могут использоваться для выполнения действий, которые более сложны, чем нажатие кнопок и якорей или ввод в поля ввода.
4.3.1. Перетащите
clickAndHold () , moveByOffset () , а затем release () перетащит элемент на страницу.
ссылка: {real-browser-test-dir} /navigator/DragAndDropSpec.groovy [] Перетаскивание также можно выполнить с помощью удобного метода dragAndDropBy () из API действий:
ссылка: {real-browser-test-dir} /navigator/DragAndDropSpec.groovy [] В этом конкретном примере элемент будет нажат, затем перетащен на 150 пикселей вправо и на 200 пикселей вниз перед тем, как отпустить.
Предупреждение | Перемещение в произвольные места с помощью мыши в настоящее время не поддерживается драйвером HTMLUnit, но перемещение непосредственно к элементам поддерживается. |
4.3.2. Нажатие клавиши Control
Щелчок по нескольким элементам, например по элементам в списке, удерживая клавишу Control, выполняется так же, как нажатие клавиши Shift.
ссылка: {real-browser-test-dir} /navigator/ControlClickSpec.groovy []
ссылка: {реальный-браузер-тест-каталог} / навигатор / ControlClickSpec.Groovy [] Вы не можете выполнить это действие в настоящее время.
Вы вошли в систему с другой вкладкой или окном. Перезагрузите, чтобы обновить сеанс.
Вы вышли из системы на другой вкладке или в другом окне. Перезагрузите, чтобы обновить сеанс.
Рекурсивная структура последовательностей Хофштадтера
Я медленно (но верно!) Пробираюсь через Годель, Эшер, Бах: вечная золотая коса Дугласа Хофштадтера.Как я уже упоминал ранее, до сих пор это было довольно классное чтение, заставляющее задуматься; изучение некоторых довольно глубоких идей как из математики, так и из информатики.
Одна из таких глубоких идей - это рекурсия . После представления идеи забавным диалогом «история в рассказе в рассказе» нам показывают несколько своеобразных последовательностей: начиная с последовательностей G (n), H (n), F (n) и M (n). Это примеры последовательности Хофштадтера . Уловка этих последовательностей не только в том, что они определены рекурсивно, но и в том, что они фактически являются нелинейно рекурсивными .G и H оба определены в терминах самих композиций. Еще более странно, что F и M определены в терминах вложенных композиций друг друга!
Упражнение, которое Хофштадтер дает читателю, состоит в том, чтобы определить рекурсивные структуры, которые могут быть созданы путем формирования графа каждой из последовательностей: где мы помечаем некоторые узлы от «1» до «n», и пусть «n» будет узлом непосредственно над узлом G (n) на графике. Как мы увидим, графы разветвляются вверх и вниз, как дерево, и мы сможем заметить некоторые рекурсивные шаблоны, которые определяют бесконечное построение всего графа.
Итак, приступим! Чтобы избавиться от необходимости вычислять значения последовательности вручную, мы закодируем некоторые рекурсивные функции в Clojure, которые мы можем использовать, чтобы легко находить значения для каждой из последовательностей Хофштадтера.
Последовательность G (n) Hofstadter
Первая показанная нам последовательность - это последовательность G (n), определяемая следующим образом.
Это переводится непосредственно в следующую рекурсивную функцию Clojure.
[Обратите внимание, что здесь мы больше озабочены удобочитаемостью, а не эффективностью, поэтому мы не будем беспокоиться о том, что мы легко взорвем наш стек для достаточно большого значения n, вычислив последовательность таким образом.Правильный способ сделать это в Clojure - использовать мемоизацию и ленивые последовательности - на самом деле Стюарт Хэллоуэй и Аарон Бедра даже используют точный пример последовательностей F / M Хофштадтера для демонстрации этих методов в своей книге Программирование Clojure !]
Затем мы можем использовать эту функцию для генерации диапазона значений для G (n), как показано ниже.
(диапазон 1 (инкр.21))
;> (1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21)
(карта g (диапазон 1 (инк 21)))
;> (1 1 2 3 3 4 4 5 6 6 7 8 9 9 10 11 11 12 12 13)
Теперь мы можем фактически использовать эту последовательность для создания графа, который визуализирует структуру рекурсии, как это делает Хофштадтер. в книге.Что мы делаем, так это начинаем с узла с меткой «1» внизу, и для каждого нового узла с меткой «n» мы помещаем этот узел непосредственно поверх узла с меткой «G (n)».
Так, например, если мы сведем в таблицу n и G (n) так, как мы их вычислили ...
n: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
G (n): 1 1 2 3 3 4 4 5 6 6 7 8 8 9 9 10 11 11 12 12 13
Тогда мы можем просто прочитать это прямо из таблицы. Таким образом, узел «2» располагается поверх узла «1», узел «3» - поверх узла «2», узлы «4» и «5» оба идут поверх «3» и так далее.
Когда мы это сделаем, мы сгенерируем следующий график.
В этом графике есть несколько очень хороших моментов. Во-первых, мы можем сразу заметить, как наши старые друзья, числа Фибоначчи, продвигаются вверх по самой правой ветви графика. Учитывая способ построения графика, это означает, что если F n является n-м числом Фибоначчи, то G (F n ) = F n-1 , что интересно.
Но более того, обратите внимание, что у нас, кажется, одна и та же форма повторяется снова и снова, когда мы ответвляемся.Начиная с узла «3», весь граф, кажется, полностью определяется повторением этой одной формы на двух ветвях (я закодировал разные вхождения разными цветами).
То есть, мы можем определить граф (назовем его G) рекурсивно. Мы начнем с выделения повторяющейся формы, которая будет исходной формой для G:
.
И у нас есть, что граф G определен в терминах самого себя: мы заменяем буквы 'G, которые появляются в верхнем левом и правом углах точными копиями графа G.И эти новые копии G, в свою очередь, будут определяться дополнительными копиями G и так далее. По мере того, как мы постоянно добавляем все больше и больше копий G, график будет ветвиться вверх и вниз, как показано.
И мы видим, что эта рекурсивная структура совпадает с той, которая была у нас для помеченных узлов ранее!
Итак, это рекурсивная структура для функции G. Но затем Хофштадтер просит нас рассмотреть зеркальное отображение графа G (узлы по-прежнему помечены слева направо, но мы перевернули структуру по горизонтали).
То есть мы должны рассмотреть следующий график (который мы назовем G flip ):
Теперь повторяющаяся «форма» рекурсивной структуры G flip очевидна: это просто зеркальное отображение рекурсивной структуры для нашего исходного графа G.
Однако предположим, что мы хотим получить рекурсивную функцию, которую мы, возможно, использовали для построения этого графика?
У нас было, что граф G построен из функции G (n) = n - G (G (n-1)). Если немного изменить это уравнение, мы можем получить n = G (n) + G (G (n-1)).Затем, учитывая, что мы определяем n как номер узла на вершине G (n) в графе, мы можем видеть, что это уравнение фактически сообщает нам инвариант для графа:
- Выберите любой узел n (например, n = 6).
- Обратите внимание на узел непосредственно под n (т.е. узел G (n), который в нашем примере равен G (6) = 4).
- Мы переходим к узлу n-1 (например, n = 5 в нашем примере: в нашем случае мы переместились за левую часть графика и вернулись к правой стороне.)
- Затем мы рассматриваем узел, который находится ровно на два узла ниже узла n-1. То есть, поскольку G (n-1) является узлом непосредственно ниже n-1, мы хотим G (G (n-1)), следующий узел вниз. (В нашем примере G (G (5)) = G (3) = 2.)
- И вот инвариант: этот узел при добавлении к узлу, который мы отметили на шаге 2, всегда должен давать нам n; т.е. G (n) + G (G (n-1)) = п. (В нашем примере 2 + 4 = 6, так что это верно.)
Таким образом, мы можем использовать эту процедуру, чтобы предложить способ, которым мы могли бы найти уравнение для инварианта графика G flip .Попробовав разные вещи и немного поработав, мы можем прийти к следующему «решению» для инварианта:
- Если мы выберем узел n,
- Затем узел непосредственно под узлом n-1, т.е. G перекидной (n-1) ...
- ... при добавлении к узлу, полученному перемещением двух узлов вниз от n, т.е. G flip (G flip (n)) ...
- ... должно быть в сумме n, т.е. G отражение (n-1) + G отражение (G отражение (n)) = n.
Итак, у нас есть G flip (n-1) + G flip (G flip (n)) = n для нашего инварианта.
Если мы сделаем быструю визуальную проверку на приведенной выше диаграмме, то окажется, что это действительный инвариант - по крайней мере, для узлов над узлом «3». И это вроде как симметрично исходной функции G (n), так что это приятно.
Но ведь проблема в нашей формуле? Мы не можем изменить это уравнение, чтобы получить выражение для G flip (n). Сдвиг нашего индекса n вперед нам не поможет, и чтобы добраться до этого вложенного члена G flip (n), нам потребуется разумная обратная функция G flip -1 (n) определено, о чем нам уже говорит график, невозможно.(Например, G flip (9) = 6 и G flip (10) = 6, так что такое G flip -1 (6)? Это 9 или 10? Может не то и другое.)
На самом деле кажется очень трудным найти правильно рекурсивную формулу для создания графика G flip , что в некотором роде удивительно, учитывая, что G имеет относительно хорошую рекурсивную формулу. Я определенно потратил несколько часов на попытки, но безуспешно. Может, это даже невозможно?
Последовательность H (n) Hofstadter
Далее нам дается последовательность H (n), которая выглядит так:
Это очень похоже на последовательность G (n), за исключением того, что у нас есть другой уровень рекурсивной вложенности.
Еще раз, перевод в рабочую функцию Clojure является механическим.
Мы можем легко получить первые 28 значений для H (n), сопоставив функцию в диапазоне [1, 28]:
(диапазон 1 (inc 28))
;> (1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
;> 23 24 25 26 27 28)
(карта h (диапазон 1 (инкремент 28)))
;> (1 1 2 3 4 4 5 5 6 7 7 8 9 10 10 11 12 13 13 14 14 15 16
;> 17 17 18 18 19)
И, как и в случае с функцией G (n), функцию H (n) можно использовать для генерации график.Мы делаем то же самое, что и раньше, убеждаясь, что узел с меткой «n» всегда находится прямо над узлом с меткой «H (n)».
Итак, мы сведем значения в таблицу, как и раньше:
n: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
H (n): 1 1 2 3 4 4 5 5 6 7 7 8 9 10 10 11 12 13 13 14 14 15
n: 23 24 25 26 27 28
H (n): 16 17 17 18 18 19
И тогда мы можем использовать эту таблицу, чтобы помочь нам начертите следующий график:
Здесь мы снова видим повторяющийся шаблон, который определяет весь граф, начиная с узла «4» и далее.(У нас также есть своего рода «удлиненная последовательность Фибоначчи», появляющаяся с правой стороны, где каждый узел представляет собой сумму узла непосредственно под ним вместе с узлом , двумя узлами под ним (так что мы идем вверх) из узла '4' внизу, мы имеем 4 = 3 + 1, 6 = 4 + 2, 9 = 6 + 3, 13 = 9 + 4 и т. д.))
Итак, мы можем определить график для H (n), который мы назовем H, используя следующий строительный блок:
Повторяющаяся форма для графика H (n) очень похожа на форму для G (n), но каждый раз у нас есть дополнительный узел в правой ветви.Наш граф будет расти и развиваться по мере того, как мы добавляем новые копии H, точно так же, как это делал граф G.
Но давайте рассмотрим зеркальное отображение H, которое мы назовем H flip :
Учитывая сходство между графиками G и H, можно ожидать, что будет соответствующее сходство между графиками G flip и H flip . Точнее, поскольку у нас был инвариант G flip (n-1) + G flip (G flip (n)) = n для графа G flip , мы могли бы сразу задаться вопросом, есть ли такой же вид симметричное уравнение для H flip работает, т.е.
H отражение (n-1) + H отражение (H отражение (H отражение (n))) = n.
Конечно, держится! Таким образом, это выражение действительно описывает инвариант для графа H flip . Однако у нас все еще есть та же проблема, что и раньше. Это рекуррентное уравнение не может сходиться к нашему базовому случаю как таковое, и мы не можем переписать его в выражение для H flip (n). К сожалению, это означает, что мы не можем использовать это уравнение для построения графика.
(Поскольку графики G flip и H flip очень похожи, мы также ожидаем, что поиск рекурсивного алгебраического уравнения для H flip вызовет такую же сложность, как мы видели, когда пытались найти выражение для G переверните . Так что продолжим спокойно; кроме того, у нас есть другие интересные последовательности, на которые стоит обратить внимание!)
Последовательности F (n) и M (n) Хофштадтера
Вот интересная пара последовательностей. Обратите внимание, что они рекурсивно переплетаются и определяются друг относительно друга:
Мы можем записать эти последовательности как взаимно рекурсивные функции Clojure (обратите внимание, что здесь мы должны использовать предварительное объявление.)
Затем мы можем использовать эти функции для получения первых 33-34 значений для каждой последовательности следующим образом.
Сначала посмотрим на график для F (n). Мы можем использовать нашу функцию Clojure f для генерации первых 34 значений путем сопоставления диапазона [1, 34]:
(диапазон 1 (inc 34))
;> (1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
;> 23 24 25 26 27 28 29 30 31 32 33 34)
(карта f (диапазон 1 (инкр. 34)))
;> (1 2 2 3 3 4 5 5 6 6 7 8 8 9 9 10 11 11 12 13 13 14 14 15
;> 16 16 17 17 18 19 19 20 21 21)
Затем мы можем свести их в таблицу и использовать таблицу для построения графика для F (n), которое мы обозначим F.
Таблица выглядит следующим образом:
n: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
F (n): 1 2 2 3 3 4 5 5 6 6 7 8 8 9 9 10 11 11 12 13 13 14
n: 23 24 25 26 27 28 29 30 31 32 33 34
F (n): 14 15 16 16 17 17 18 19 19 20 21 21
Используя значения из таблицы (с тем же условием размещения n над F (n), по крайней мере, для больших значений n (мы не будем слишком беспокоиться о том факте, что узлы '1', '2' и '3', похоже, не слишком хорошо работают с этой идеей)) , мы приходим к следующей диаграмме:
Мы можем заметить здесь своеобразную пару повторяющихся паттернов.У нас есть одна из форм, повторяющаяся снова и снова на правой ветви (которую я закрасил в оттенки , синий, на графике). Но обратите внимание на форму, которая каждый раз повторяется на левой ветке (которую я покрасила в красный цвет , ). Вам она кажется знакомой?
Ага! В левой ветви у нас есть тот же график G, с которым мы работали в начале этого сообщения в блоге!
Возможно, немного удивительно, что на графике появляется G для явно не связанной функции F (n).Но если мы посмотрим на функцию G (n) для сравнения, то увидим, что F (n) и G (n) похожи: единственная разница в том, что F (n) вызывает функцию M (n) для внешнего рекурсивного вызова.
Итак, рекурсивный строительный блок для графа F выглядит следующим образом:
Наконец, мы рассмотрим функцию M (n). Сопоставляя нашу функцию m с диапазоном [1, 33] в Clojure REPL, мы получаем следующее:
(диапазон 1 (inc 33))
;> (1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
;> 23 24 25 26 27 28 29 30 31 32 33)
(карта m (диапазон 1 (инкремент 33)))
;> (0 1 2 2 3 4 4 5 6 6 7 7 8 9 9 10 11 11 12 12 13 14 14 15
;> 16 16 17 17 18 19 19 20 20)
Эти значения можно свести в таблицу следующим образом.
n: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 9 1780 M (n): 0 1 2 2 3 4 4 5 6 6 7 7 8 9 9 10 11 11 12 12 13 14
n: 23 24 25 26 27 28 29 30 31 32 33
M (n): 14 15 16 16 17 17 18 19 19 20 20
Итак, график M для функции M (n) выглядит так:
Еще раз, у нас есть повторяющийся шаблон, который выстраивается на правой ветви каждый раз, начиная с узла 4 и далее (который я окрасил в синий цвет ), и в каждой точке у нас есть график для G появляется на левой ветви (в красный .)
Обратите внимание, что у нас также есть очень заметная башня чисел Фибоначчи, появляющаяся в левой части, что довольно круто. Это означает, что функция M (n) обладает тем же интересным свойством, что и функция G (n): если F n является n-м числом Фибоначчи, то M (F n ) = F n-1 .
Наконец, мы можем отметить, что граф M рекурсивно строится из следующей базы:
В некоторой степени мы можем думать о графике G, который появляется как на графиках F, так и на M, как намекающий на взаимодействие между ними из-за взаимной рекурсии между функциями F (n) и M (n).
В качестве последнего замечания, мы можем сделать замечание о простоте рекурсивных структур здесь, особенно с этими двумя последними переплетенными функциями F (n) и M (n). Рекурсивные строительные блоки довольно элегантны, учитывая, насколько запутанными были рекуррентные уравнения.
Итак, мы уже видели несколько довольно изящных рекурсивных структур. В своей книге Хофштадтер продолжает говорить о другой рекурсивной последовательности Q (n), которая обладает странным свойством: она хаотически перемещается по мере увеличения значения n.На самом деле о поведении этой последовательности известно немногое. Возможно, нам стоит изучить это в одном из будущих постов в блоге?
Сдвиги в составе сообществ и плотности населения существенно влияют на функцию экосистемы, несмотря на неизменное богатство, Ecology Letters
Изменения в составе сообществ и плотности населения существенно влияют на функцию экосистемы, несмотря на неизменное богатство.
Письма об экологии
(
ЕСЛИ
8.665
)
Дата публикации: 2017-09-17
, DOI:
10.1111 / эл.12828
Юрг В. Спаак; Ян М. Бэрт; Дональд Дж. Бэрд; Нико Эйзенхауэр; Лоррейн Малтби; Франческо Помати; Виктория Радчук; Джейсон Р.Рор; Пол Дж. Ван ден Бринк; Фредерик Де Лендер
Значительное внимание уделяется влиянию изменения окружающей среды на функцию экосистемы в результате изменения видового богатства.Однако изменение окружающей среды может повлиять на функцию экосистемы, не влияя на ее богатство, в первую очередь влияя на плотность населения и состав сообществ. Используя теоретическую модель, мы обнаруживаем, что, несмотря на инвариантное богатство, (1) небольшие воздействия окружающей среды могут уже привести к коллапсу функции; (2) конкурентоспособность может быть менее важным детерминантом изменения функции экосистемы, чем избирательность движущей силы изменения окружающей среды, и (3) влияние на функцию экосистемы возрастает, когда влияние на состав больше.Мы также представляем дополнительный статистический анализ 13 наборов данных о сообществах фитопланктона и перифитона, подвергшихся воздействию химических стрессоров, и показываем, что влияние на первичную продукцию при неизменном богатстве варьировалось от -75% до + 10%. Мы пришли к выводу, что цели защиты окружающей среды, основанные на показателях богатства, могут недооценивать экологические последствия экологических изменений.
更新 日期 : 2017-09-17
Функция ресничек и состав моторных белков фаллопиевых труб человека | Репродукция человека
Аннотация
ВОПРОС ДЛЯ ИССЛЕДОВАНИЯ
Каков состав моторных белков и функция ресничек фаллопиевых труб человека?
КРАТКИЙ ОТВЕТ
Хотя состав моторных белков и функция ресничек FT человека напоминают респираторные реснички, самки с первичной цилиарной дискинезией (PCD) не обязательно бесплодны.
ЧТО УЖЕ ИЗВЕСТНО
FT покрыты множеством подвижных ресничек, которые при просвечивающей электронной микроскопии показывают ультраструктуру 9 + 2. Сообщения о клинических случаях предполагают повышенную частоту субфертильности и внематочной беременности у женщин с PCD, заболеванием, характеризующимся дисфункцией подвижных ресничек и жгутиков.
ДИЗАЙН, РАЗМЕР, ПРОДОЛЖИТЕЛЬНОСТЬ
Это исследование состояло из наблюдательного лабораторного исследования образцов FT от пяти здоровых женщин, набранных с апреля 2012 года по декабрь 2013 года, и описательного наблюдательно-ретроспективного анализа клинической базы данных PCD.
УЧАСТНИКИ / МАТЕРИАЛЫ, УСТАНОВКИ, МЕТОДЫ
Ткань FT человека была получена от пяти здоровых женщин после перевязки маточных труб во время кесарева сечения. Состав моторных белков оценивали с помощью иммунофлуоресцентной микроскопии с использованием антител против динеиновых плеч и субъединиц регуляторного комплекса нексин-динеин. Подвижность ресничек исследовали с помощью высокоскоростной видеомикроскопии. Был проведен ретроспективный поиск информации о зачатии и родах в нашей базе данных лиц с PCD.
ОСНОВНЫЕ РЕЗУЛЬТАТЫ И РОЛЬ ШАНСА
Состав моторных белков ресничек FT человека был идентичен составу респираторных ресничек. Реснички FT показали скоординированное биение, что привело к направленному потоку жидкости в полость матки. Мы идентифицировали девять пациентов с PCD с тяжелой дисфункцией респираторных ресничек, которые родили детей после самопроизвольного зачатия. Это говорит о том, что биение ресничек не является ключевым двигателем транспорта яйцеклетки.
ОГРАНИЧЕНИЯ, ПРИЧИНА ПРЕДУПРЕЖДЕНИЯ
Реснички FT пораженных PCD женщин не были доступны для анализа.Таким образом, остается доказать, что реснички FT действительно обнаруживают те же дефекты, что и респираторные реснички у лиц с PCD. Необходимы комплексные эпидемиологические исследования, чтобы определить степень женской (суб) фертильности при PCD.
РАСШИРЕННОЕ ПРИМЕНЕНИЕ РЕЗУЛЬТАТОВ
Знание точного белкового состава и функции ресничек FT будет способствовать лучшему пониманию потока жидкости, генерируемого ресничками при размножении самок. Эти данные важны для последующих исследований функции и белкового состава ресничек FT у пациентов с PCD.
ФИНАНСИРОВАНИЕ ИССЛЕДОВАНИЙ / КОНКУРЕНЦИЯ ИНТЕРЕСОВ
Эта работа была поддержана Европейской комиссией FP7 (Седьмая рамочная программа исследований), гранты № 305404 (BESTCILIA) и 241955 (SYSCILIA) Х. Омрану, Deutsche Forschungsgemeinschaft (DFG OM 6 / 4, OM 6/5) и IZKF Muenster (Om2 / 009/12). Все авторы заявляют, что у них нет конкурирующих интересов.
Введение
Реснички и жгутики (хвосты сперматозоидов) представляют собой волосоподобные органеллы на основе микротрубочек, которые выступают с поверхности почти всех типов клеток человеческого тела (Fliegauf et al., 2007). Большинство типов клеток имеют единственную ресничку (моноцилию или первичную ресничку), которая может быть подвижной или неподвижной, тогда как некоторые клетки строят 200-300 множественных подвижных ресничек (MMC).
ММС выстилают верхние и нижние дыхательные пути, эпендимные клетки желудочков головного мозга и фаллопиевы трубы (FT) женской репродуктивной системы. Респираторные реснички отвечают за мукоцилиарный клиренс, важный механизм предотвращения хронического деструктивного заболевания дыхательных путей, а эпендимные реснички продвигают спинномозговую жидкость по водопроводу (Ibañez-Tallon et al., 2003). В нескольких исследованиях было документально подтверждено, что у женщин с ФТ присутствует ГМК (Афзелиус и Элиассон, 1983; Лион, и др., , 2006). ФТ человека представляют собой трубчатые серомукозные органы, соединяющие яичники с маткой. Анатомические структуры включают (от латерального к медиальному) фимбрию, воронку, ампулу и перешеек (рис. 1А). Ресничные и секреторные клетки представляют собой основные типы клеток слизистой оболочки. Ресничные клетки в основном обнаруживаются на вершине складок слизистой оболочки (Lyons et al. , 2006; Ezzati et al., 2014). Предполагается, что скоординированное биение ресничек вместе с мышечными сокращениями играет роль в воспроизводстве человека, направляя ооцит через FT в матку (Lyons et al. , 2006). Этот процесс изменяется в зависимости от уровня гормонов, а также от состава слизи в трубах и может быть нарушен инфекциями, курением и гинекологическими заболеваниями, такими как эндометриоз (Ezzati et al. , 2014).
Рисунок 1
( A ) Анатомия фаллопиевой трубы человека (FT), представляющая отдельные сегменты: (a) фимбрия, (b) воронка, (c) ампула и (d) перешеек.( B ) Мультицилифицированные клетки FT от здорового контрольного человека, окрашенные антителом, направленным против ацетилированного α-тубулина (зеленый) в качестве цилиарного маркера. Ядра окрашивали Hoechst33342 (синий). Масштабная линейка равна 10 мкм. ( C ) Поперечное сечение TEM реснички FT свиньи, показывающее состав аксонемы, напоминающий состав респираторных ресничек человека с микротрубочковой структурой 9 + 2, внешними плечами динеина, внутренними плечами динеина, регуляторным комплексом нексин-динеин и радиальными спицами. Масштабная шкала равна 200 нм.( D ) Схема поперечного сечения ресничек с указанием ультраструктурных компонентов.
Рисунок 1
( A ) Анатомия фаллопиевой трубы человека (FT), представляющая отдельные сегменты: (а) фимбрия, (б) воронка, (в) ампула и (г) перешеек. ( B ) Мультицилифицированные клетки FT от здорового контрольного человека, окрашенные антителом, направленным против ацетилированного α-тубулина (зеленый) в качестве цилиарного маркера. Ядра окрашивали Hoechst33342 (синий). Масштабная линейка равна 10 мкм.( C ) Поперечное сечение TEM реснички FT свиньи, показывающее состав аксонемы, напоминающий состав респираторных ресничек человека с микротрубочковой структурой 9 + 2, внешними плечами динеина, внутренними плечами динеина, регуляторным комплексом нексин-динеин и радиальными спицами. Масштабная шкала равна 200 нм. ( D ) Схема поперечного сечения ресничек с указанием ультраструктурных компонентов.
Первичная цилиарная дискинезия (ПЦД) - это аутосомно-рецессивное заболевание подвижных ресничек, ведущее к заболеванию верхних и нижних дыхательных путей из-за нарушения функции респираторных цилиар.Была выдвинута гипотеза, что самки с PCD могут быть затронуты субфертильностью, потому что генетический дефект также влияет на реснички FT (Afzelius and Eliasson, 1983; Halbert et al. , 1997; Lyons et al. , 2006).
FT реснички имеют длину ~ 10 мкм и диаметр 0,25 мкм (Satir, 1992). Анализ с помощью трансмиссионной электронной микроскопии (ПЭМ) показал, что поперечные срезы ресничек FT имеют ту же ультраструктуру аксонем с другими подвижными ресничками, такими как респираторные и эпендимные MMC или жгутики сперматозоидов (Dirksen and Satir, 1972; Afzelius, 1976, 2004; Halbert et al. ., 1976; Ibañez-Tallon et al. , 2003; Lyons et al. , 2006; Ezzati et al. , 2014). Девять периферических дублетных микротрубочек окружают две отдельные центральные микротрубочки, чтобы сформировать 9 + 2 микротрубочковую структуру (Fig. 1C и D). Внешнее и внутреннее плечо динеина (ODAs и IDA соответственно) содержат моторные белки, участвующие в генерации биений ресничек. Регуляторный комплекс нексин-динеин (N-DRC) соединяет внешние дуплеты, а радиальные спицы соединяют периферические дуплеты с центральным парным аппаратом (рис.1С и Г).
Хотя реснички обнаруживают одинаковую ультраструктуру с помощью TEM, состав моторных белков, которые вызывают генерацию биений ресничек, различается у разных видов. Например, ОПР Chlamydomonas reinhardtii представляет собой трехглавую структуру, содержащую три тяжелые цепи динеина, тогда как ОПР человека состоит только из двух голов (O'Toole et al. , 2012). У человека состав моторных белков аксонемы ресничек хвостов сперматозоидов отличается от состава подвижных респираторных ресничек (Fliegauf et al., 2005). В ресничках FT человека состав моторных белков ресничек еще не изучен. Это побудило нас подробно проанализировать состав моторных белков ресничек FT человека и, как следствие, функцию ресничек. Кроме того, мы провели поиск в нашей базе данных PCD для получения информации о зачатии и родах у женщин PCD с тяжелым респираторным заболеванием.
Материалы и методы
Субъектов
Ткань
FT была получена от пяти женщин, подвергшихся стерилизации во время кесарева сечения.Для сравнительного анализа образцы клеток респираторного эпителия были взяты у здоровых людей, как описано ранее (Raidt et al. , 2014). Средний возраст был одинаковым в обеих группах (FT: 34,6 года по сравнению с контрольным респираторным эпителием: 32,2 года). Письменное информированное согласие на участие в этом исследовании было получено от каждого человека до взятия образцов ткани FT или респираторных клеток. Исследование было одобрено этическим комитетом Вестфальского университета Вильгельма, Мюнстер, Германия.
Иммунофлуоресцентный микроскопический анализ высокого разрешения
Криосрезы FT человека распределяли на предметные стекла, сушили на воздухе и хранили при -80 ° C перед использованием. Репрезентативные срезы были приготовлены из каждого сегмента FT, чтобы визуализировать распределение и состав подвижных ресничек. Кроме того, мы изучали одиночные ресничные клетки FT, полученные при щеточной биопсии фимбрии и воронки, а также респираторных ресничек после взятия пробы из носа.
Клетки и ткани обрабатывали и инкубировали первичными и вторичными антителами, как сообщалось ранее (Omran and Loges, 2009; Hjeij et al., 2014). В качестве маркеров цилиарной аксонемы используются моноклональные мышиные антитела против ацетилированного α-тубулина (Sigma-Aldrich, Taufkirchen, Германия) или поликлональные кроличьи антитела против α / ß тубулина (Cell Signaling Technology, New England Biolabs, Франкфурт-на-Майне, Германия). использовался. Ранее сообщалось о поликлональных кроличьих антителах, направленных против DNAH5, DNALI1, DNAH9, CCDC39 и LRRC48, а также о моноклональных мышиных антителах против DNAI2 (Fliegauf et al. , 2005; Zariwala et al. , 2006; Loges et al. al., 2008, 2009; Merveille et al. , 2011; Wirschell et al. , 2013). Поликлональные кроличьи антитела против DNAI1 были получены от Sigma-Aldrich. В качестве вторичных антител использовали антитела против мышиного Alexa Fluor 488 и против кроличьего Alexa Fluor 546 (Life Technologies, Дармштадт, Германия). Ядра окрашивали Hoechst 33 342 (Sigma-Aldrich). Иммунофлуоресцентные изображения (IF) получали с помощью Zeiss Apotome Axiovert 200 и обрабатывали с помощью AxioVision v.4.8 (Carl Zeiss AG, Геттинген, Германия) и Adobe Creative Suite 4 (Adobe Systems, Сан-Хосе, Калифорния, США).
Анализ с помощью высокоскоростной видеомикроскопии
Непосредственно после хирургического удаления ткани FT человека суспендировали в среде RPMI 1640. Анализ с помощью высокоскоростной видеомикроскопии (HVMA) проводился как в полосах реснитчатых клеток, которые были получены путем чистки фимбрии и инфундибулума цитологической щеткой (Engelbrecht Medicine and Laboratory Technology, Edermünde, Германия), так и на целых срезах тканей всех сегментов. продольно разрезанных яйцеводов. HVMA в клетках респираторного эпителия выполняли, как описано ранее (Raidt et al., 2014). Респираторные клетки получали от здоровых небеременных добровольцев в произвольное время менструального цикла. Видеоанализ Sisson-Ammons (SAVA) был использован для анализа паттерна и частоты биений ресничек (Sisson et al. , 2003).
Анализ транспорта с помощью ресничек
Транспортную функцию ресничек FT человека визуализировали с использованием частиц размером 2 мкм (2% флуоресцентные микросферы FluoSpheres, Life Technologies). Видео были записаны с помощью Hokawo 2.6 при 10-кратном увеличении с помощью цифровой камеры ORCA.flash 4.0 (Hamamatsu Photonics, Herrsching, Германия), подключенной к микроскопу PolyVar MET (Reichert-Jung, Вена, Австрия). Перенос частиц был проанализирован с использованием Фиджи (Schindelin et al. , 2012).
Результаты
Пространственное распределение MMC в FT человека
Распределение реснитчатых клеток вдоль FT человека анализировали с помощью IF. Окрашивание срезов фимбрии, воронки, ампулы и перешейка с использованием антител против DNAH5 и ацетилированного α-тубулина показало, что все сегменты были заселены MMC, что указывает на то, что биение ресничек происходит по всей длине FT (рис.2). Плотность мультицилиатных клеток была наибольшей в фимбриях и снижалась к перешейку. Как и в респираторных ресничках, оба антитела локализованы совместно по всей длине аксонем ресничек во всех сегментах FT (Fig. 3).
Рисунок 2
Криосрезы человеческих фаллопиевых труб (FT) от здоровых контрольных лиц были дважды помечены антителами, направленными против ацетилированного тубулина (зеленый) и DNAH5 (красный). Оба белка совместно локализуются (желтые) вдоль аксонемы ресничек.Ядра окрашивали Hoechst33342 (синий). Окрашивали отдельные сегменты FT человека: ( A ) фимбрия, ( B ) infundibulum, ( C ) ампулла и ( D ) перешеек. Мультицилиндрические клетки присутствовали во всех сегментах FT. ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 50 мкм.
Рисунок 2
Криосрезы человеческих фаллопиевых труб (FT) от здоровых контрольных лиц были дважды помечены антителами, направленными против ацетилированного тубулина (зеленый) и DNAH5 (красный).Оба белка совместно локализуются (желтые) вдоль аксонемы ресничек. Ядра окрашивали Hoechst33342 (синий). Окрашивали отдельные сегменты FT человека: ( A ) фимбрия, ( B ) infundibulum, ( C ) ампулла и ( D ) перешеек. Мультицилиндрические клетки присутствовали во всех сегментах FT. ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 50 мкм.
Рисунок 3
Клетки респираторного эпителия (респ) и фаллопиевых труб (FT) здоровых контрольных лиц после биопсии щеткой были дважды помечены антителами, направленными против ацетилированного α-тубулина (зеленый) и DNAH5 (красный).Оба белка совместно локализуются (желтый цвет) по всей длине аксонемы ресничек в обоих типах клеток. Ядра окрашивали Hoechst33342 (синий). ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 10 мкм.
Рисунок 3
Клетки респираторного эпителия (соответственно) и фаллопиевых труб (FT) здоровых контрольных лиц после биопсии кистью были дважды помечены антителами, направленными против ацетилированного α-тубулина (зеленый) и DNAH5 (красный). Оба белка совместно локализуются (желтый цвет) по всей длине аксонемы ресничек в обоих типах клеток.Ядра окрашивали Hoechst33342 (синий). ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 10 мкм.
Состав моторных белков ресничек FT человека
Для дальнейшего анализа состава моторных белков ресничек FT, криосрезы, а также полоски эпителиальных клеток из вороночного сегмента использовали для дополнительного окрашивания IF. Субклеточная локализация нескольких компонентов ODA, тяжелых цепей аксонемного динеина DNAH5 (рис.3) и DNAH9 (рис.4A), а также промежуточных цепей аксонемного динеина DNAI1 и DNAI2 (Fig. 4B and C), показали идентичное распределение в ресничках FT человека, как и в респираторных ресничках (Supplementary data, Fig. S1). Точно так же компоненты IDA, DNALI1 (фиг. 4D) и N-DRC, LRRC48 (фиг. 4E), и связанный с N-DRC белок, CCDC39 (фиг. 4F), показали одинаковый паттерн окрашивания аксонем у человека. FT как респираторные реснички (дополнительные данные, рис. S1).
Рисунок 4
Субклеточная локализация моторных белков ресничек в мультицилилированных клетках FT человека здоровых людей из контрольной группы.Антитела, направленные против ацетилированного тубулина ( A и B , D - F ; зеленый) или α / ß-тубулина ( C ; красный) использовали в качестве маркеров аксонемы ресничек. Антитела против моторных белков ресничек DNAH9 (A; красный; компонент тяжелой цепи плеча динеина, который, как известно, локализуется в дистальной части аксонемы ресничек в респираторных ресничках), DNAI1 (B; красный; промежуточная цепь внешнего плеча динеина), DNAI2 (C; зеленый; внешняя промежуточная цепь плеча динеина; зеленый), DNALI1 (D; красный; компонент внутреннего плеча динеина), LRRC48 (E; красный; субъединица регуляторного комплекса нексин-динеин) и CCDC39 (F; красный; субъединица регуляторного комплекса нексин-динеин) были использованы.Ядра окрашивали Hoechst33342 (синий). ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 10 мкм (A, E и F) и 50 мкм (B – D) соответственно. Все белки, кроме DNAH9, локализуются по всей длине аксонем ресничек. Окрашивание в желтый цвет (слияние) указывает на совместную локализацию обоих белков в аксонеме ресничек. Как и в респираторных клетках, DNAH9 локализуется в дистальной части аксонемы ресничек, что приводит к желтому окрашиванию дистальной части аксонемы ресничек, тогда как проксимальная часть остается зеленой (A; слияние).
Рисунок 4
Субклеточная локализация моторных белков ресничек в мультицилилированных клетках FT человека здоровых людей из контрольной группы. Антитела, направленные против ацетилированного тубулина ( A и B , D - F ; зеленый) или α / ß-тубулина ( C ; красный) использовали в качестве маркеров аксонемы ресничек. Антитела против моторных белков ресничек DNAH9 (A; красный; компонент тяжелой цепи плеча динеина, который, как известно, локализуется в дистальной части аксонемы ресничек в респираторных ресничках), DNAI1 (B; красный; промежуточная цепь внешнего плеча динеина), DNAI2 (C; зеленый; внешняя промежуточная цепь плеча динеина; зеленый), DNALI1 (D; красный; компонент внутреннего плеча динеина), LRRC48 (E; красный; субъединица регуляторного комплекса нексин-динеин) и CCDC39 (F; красный; субъединица регуляторного комплекса нексин-динеин) были использованы.Ядра окрашивали Hoechst33342 (синий). ДИК представляет собой дифференциальный интерференционный контраст. Масштабная линейка равна 10 мкм (A, E и F) и 50 мкм (B – D) соответственно. Все белки, кроме DNAH9, локализуются по всей длине аксонем ресничек. Окрашивание в желтый цвет (слияние) указывает на совместную локализацию обоих белков в аксонеме ресничек. Как и в респираторных клетках, DNAH9 локализуется в дистальной части аксонемы ресничек, что приводит к желтому окрашиванию дистальной части аксонемы ресничек, тогда как проксимальная часть остается зеленой (A; слияние).
Характер и частота биения ресничек в фаллопиевых трубах человека
Анализ
HVMA 25 видео полосок клеток FT при 25 ° C выявил среднюю частоту биений ресничек 3,48 Гц, что немного ниже нормального диапазона (4,25–11,63 Гц) респираторных ресничек (Raidt et al. , 2014) . HVMA полосок реснитчатых клеток обнаруживает сходный паттерн биения ресничек с таковым в респираторных клетках с отчетливыми движениями вперед и восстановления (дополнительные данные, видео S1 и S2).Анализ вида сверху показал регулярное плоское биение вдоль продольной оси без значительного отклонения в сторону. Биение ресничек соседних ресничек было скоординировано (Дополнительные данные, Видео S3). Мы не наблюдали каких-либо значительных различий в частоте или характере биений в ресничках разных анатомических сегментов FT.
Транспорт с помощью ресничек в фаллопиевых трубах человека
Затем мы оценили с помощью HVMA, могут ли реснички FT генерировать направленный поток жидкости.В самом деле, быстрое и синхронизированное движение ресничек FT вызывает ламинарный поток жидкости на поверхности клеток FT. Этот ламинарный поток привел к быстрой транспортировке флуоресцентных шариков от латерального края FT к матке со средней скоростью 32,43 мкм / с (рис. 5; дополнительные данные, видео S4).
Рисунок 5
Цветные следы флуоресцентных шариков, образованные синхронным движением ресничек маточных труб. Флуоресцентные шарики перемещались от латерального края маточной трубы к матке со средней скоростью 32.43 мкм / с. Изображения были сняты с интервалом 0 мс ( A ), 100 мс ( B ), 200 мс ( C ) и 300 мс ( D ).
Рисунок 5
Цветные следы флуоресцентных шариков, образованные синхронным движением ресничек маточных труб. Флуоресцентные шарики транспортировались от латерального края маточной трубы к матке со средней скоростью 32,43 мкм / с. Изображения были сняты с интервалом 0 мс ( A ), 100 мс ( B ), 200 мс ( C ) и 300 мс ( D ).
Вынашивание потомства у женщин с PCD
Основываясь на нашем анализе того, что FT и респираторные подвижные реснички имеют идентичный состав моторных белков и сходный паттерн и частоту биений ресничек, мы провели поиск в нашей базе данных PCD для информации о беременности и родах у женщин с PCD с тяжелыми дефектами моторных белков ресничек. Мы идентифицировали девять женщин с ПЦД, которые родили детей без какой-либо репродуктивной помощи. У каждого из этих людей были классические признаки PCD, включая хронический риносинусит, хронический влажный кашель, рецидивирующую пневмонию и бронхоэктазы.У трех самок (OP-118, OP-527 II1, OP-515) генетический анализ выявил двуаллельные мутации, предсказывающие полное отсутствие моторного белка DNAH5 в аксонеме ресничек. В OP-1795 II3 гомозиготная мутация в гене RSPh5A приводит к полному отсутствию белков RSPh2, RSPh5A и RSPH9 радиальной головки. У семи из девяти пациентов респираторные MMC были доступны для IF, что показало отсутствие основных моторных белков у всех этих людей. Сводка диагностических деталей приведена в таблице I.
Таблица I
Диагностические данные у женщин с ПЦД, родивших после самопроизвольного зачатия.
| Тема . | HVMA респираторной MMC . | ТЕМ / ИФ респираторных MMC . | Мутация . | Кол-во детей . | Состав тела . |
|---|---|---|---|---|---|
| OP-118 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие ДНКH5 в аксонемах | DNAH5 : c.8029C> T [p.Arg2677 *] + c.10813G> A [p.Asp3605Asn] | 1 | Обратное положение |
| OP-527 II1 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме компонента ODA DNAH5 | DNAH5 : c.10815delT [p.Pro3606Hisfs22 *] + c.8642C> G [p.Ala2881Gly] | 1 | |
| F-725 II3 | Жесткий, нескоординированный CBP; сильно сниженный CBF <1 Гц | n.а. / отсутствие DNAH5 в дистальной аксонеме ресничек | DNAI1 : c.463delA [p.Lys155Argfs2 *] + n.a. | 3 | Situs inversus |
| F-649 II2 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 и компонента IDA DNALI1 | n.a. | 1 | Situs inversus |
| OP-515 | n.a. | нет данных | DNAH5 : c.376delG [p.Val126Tyrfs22 *] + c.13194_13197delCAGA [p.Asp4398Glufs16 *] | 1 | Situs inversus |
| OP-439 | Дефект ДНК | DA 902 | Дефект ДНК | DA 902 Дефектные цилии O 902 : c.3598 + 2T> C + c.10.616G> A [p.R3539H] | 2 | Situs inversus |
| OP-1795 II3 | Аномальный круговой CBP; нормальный CBF | Дефект канальцевой транспозиции в подмножестве ресничек / аксонемы отсутствие белков головки радиальной спицы RSPh2, RSPh5A и RSPH9 | RSPh5A: c.1391G> A [p.Gly464Glu] (hom) | 1 | Situs solitus |
| OP-2049 | Жесткий, нескоординированный CBP; нормальный CBF | n.a. | нет данных | 2 | Situs inversus |
| OP-1728 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 | n.a. | 1 | Situs solitus |
| Тема . | HVMA респираторной MMC . | ТЕМ / ИФ респираторных MMC . | Мутация . | Кол-во детей . | Состав тела . |
|---|---|---|---|---|---|
| OP-118 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме ДНКH5 | DNAH5 : c.8029C> T [p.Arg2677 *] + c.10813G> A [p.Asp3605Asn] | 1 | Обратное положение |
| OP-527 II1 | Некоординированный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме компонента ODA DNAH5 | DNAH5 : c.10815delT [p.Pro3606Hisfs22 *] + c.8642C> G [p.Ala2881Gly] | 1 | |
| F-725 II3 | Жесткий, нескоординированный CBP; сильно сниженный CBF <1 Гц | n.а. / отсутствие DNAH5 в дистальной аксонеме ресничек | DNAI1 : c.463delA [p.Lys155Argfs2 *] + n.a. | 3 | Situs inversus |
| F-649 II2 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 и компонента IDA DNALI1 | n.a. | 1 | Situs inversus |
| OP-515 | n.a. | нет данных | DNAH5 : c.376delG [p.Val126Tyrfs22 *] + c.13194_13197delCAGA [p.Asp4398Glufs16 *] | 1 | Situs inversus |
| OP-439 | Дефект ДНК | DA 902 | Дефект ДНК | DA 902 Дефектные цилии O 902 : c.3598 + 2T> C + c.10.616G> A [p.R3539H] | 2 | Situs inversus |
| OP-1795 II3 | Аномальный круговой CBP; нормальный CBF | Дефект канальцевой транспозиции в подмножестве ресничек / аксонемы отсутствие белков головки радиальной спицы RSPh2, RSPh5A и RSPH9 | RSPh5A: c.1391G> A [p.Gly464Glu] (hom) | 1 | Situs solitus |
| OP-2049 | Жесткий, нескоординированный CBP; нормальный CBF | n.a. | нет данных | 2 | Situs inversus |
| OP-1728 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 | n.a. | 1 | Situs solitus |
Таблица I
Диагностические данные у женщин с ПЦД, родивших после спонтанного зачатия.
| Тема . | HVMA респираторной MMC . | ТЕМ / ИФ респираторных MMC . | Мутация . | Кол-во детей . | Состав тела . |
|---|---|---|---|---|---|
| OP-118 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие ДНКH5 в аксонемах | DNAH5 : c.8029C> T [p.Arg2677 *] + c.10813G> A [p.Asp3605Asn] | 1 | Обратное положение |
| OP-527 II1 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме компонента ODA DNAH5 | DNAH5 : c.10815delT [p.Pro3606Hisfs22 *] + c.8642C> G [p.Ala2881Gly] | 1 | |
| F-725 II3 | Жесткий, нескоординированный CBP; сильно сниженный CBF <1 Гц | n.а. / отсутствие DNAH5 в дистальной аксонеме ресничек | DNAI1 : c.463delA [p.Lys155Argfs2 *] + n.a. | 3 | Situs inversus |
| F-649 II2 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 и компонента IDA DNALI1 | n.a. | 1 | Situs inversus |
| OP-515 | n.a. | нет данных | DNAH5 : c.376delG [p.Val126Tyrfs22 *] + c.13194_13197delCAGA [p.Asp4398Glufs16 *] | 1 | Situs inversus |
| OP-439 | Дефект ДНК | DA 902 | Дефект ДНК | DA 902 Дефектные цилии O 902 : c.3598 + 2T> C + c.10.616G> A [p.R3539H] | 2 | Situs inversus |
| OP-1795 II3 | Аномальный круговой CBP; нормальный CBF | Дефект канальцевой транспозиции в подмножестве ресничек / аксонемы отсутствие белков головки радиальной спицы RSPh2, RSPh5A и RSPH9 | RSPh5A: c.1391G> A [p.Gly464Glu] (hom) | 1 | Situs solitus |
| OP-2049 | Жесткий, нескоординированный CBP; нормальный CBF | n.a. | нет данных | 2 | Situs inversus |
| OP-1728 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 | n.a. | 1 | Situs solitus |
| Тема . | HVMA респираторной MMC . | ТЕМ / ИФ респираторных MMC . | Мутация . | Кол-во детей . | Состав тела . |
|---|---|---|---|---|---|
| OP-118 | Несогласованный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме ДНКH5 | DNAH5 : c.8029C> T [p.Arg2677 *] + c.10813G> A [p.Asp3605Asn] | 1 | Обратное положение |
| OP-527 II1 | Некоординированный CBP; минимальная остаточная подвижность | Дефект ODA / отсутствие в аксонеме компонента ODA DNAH5 | DNAH5 : c.10815delT [p.Pro3606Hisfs22 *] + c.8642C> G [p.Ala2881Gly] | 1 | |
| F-725 II3 | Жесткий, нескоординированный CBP; сильно сниженный CBF <1 Гц | n.а. / отсутствие DNAH5 в дистальной аксонеме ресничек | DNAI1 : c.463delA [p.Lys155Argfs2 *] + n.a. | 3 | Situs inversus |
| F-649 II2 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 и компонента IDA DNALI1 | n.a. | 1 | Situs inversus |
| OP-515 | n.a. | нет данных | DNAH5 : c.376delG [p.Val126Tyrfs22 *] + c.13194_13197delCAGA [p.Asp4398Glufs16 *] | 1 | Situs inversus |
| OP-439 | Дефект ДНК | DA 902 | Дефект ДНК | DA 902 Дефектные цилии O 902 : c.3598 + 2T> C + c.10.616G> A [p.R3539H] | 2 | Situs inversus |
| OP-1795 II3 | Аномальный круговой CBP; нормальный CBF | Дефект канальцевой транспозиции в подмножестве ресничек / аксонемы отсутствие белков головки радиальной спицы RSPh2, RSPh5A и RSPH9 | RSPh5A: c.1391G> A [p.Gly464Glu] (hom) | 1 | Situs solitus |
| OP-2049 | Жесткий, нескоординированный CBP; нормальный CBF | n.a. | нет данных | 2 | Situs inversus |
| OP-1728 | Неподвижные реснички | Дефект ODA / отсутствие аксонемы DNAH5 | n.a. | 1 | Situs solitus |
Обсуждение
Здесь мы сообщаем подробный анализ состава моторных белков и функций FT MMC человека.Мы идентифицировали ГМК по всему яйцеводу (рис. 2). Плотность мультицилиндрических клеток была самой высокой в фимбриях и снижалась по направлению к матке, что согласуется с данными, описанными с помощью ТЕА (Crow et al. , 1994). Подробный анализ состава моторных белков ресничек с помощью IF выявил идентичную локализацию белков, которые, как известно, играют важную роль в управлении движением ресничек (компоненты ODA, DNAH5, DNAI1, DNAI2 и DNAH9) и регулируют паттерн биения ресничек (компонент IDA, DNALI1 и N- DRC-связанные компоненты, LRRC48 и CCDC39) по сравнению с мультицилилированными респираторными клетками (рис.4). Это включает присутствие двух разных типов ODA, локализующихся в разных проксимальных (тип 1, содержащий DNAH5) и дистальных компартментах (тип 2, содержащий DNAH5 и DNAH9) цилиарных аксонем (Fliegauf et al. , 2005).
Прямой анализ цилиарного транспорта с использованием HVMA выявил быстрый транспорт частиц к матке (дополнительные данные, видео S4 и рис. 5). В соответствии с нашими данными IF, показывающими идентичный состав моторных белков, мы наблюдали идентичный паттерн биения ресничек в ресничках FT, как и в респираторных ресничках по HVMA (дополнительные данные, Videos S1-S3).Мы наблюдали немного меньшую частоту биений ресничек в ресничках FT, чем в респираторных ресничках и в образцах FT от небеременных индивидуумов (Lyons et al. , 2002). Возможно, это вызвано высоким уровнем прогестерона в конце беременности. Было показано, что прогестерон снижает частоту биений ресничек (Mahmood et al. , 1998; Bylander et al. , 2013). В другом исследовании измерения, проведенные в истмических частях яйцеводов у женщин, перенесших гистерэктомию по поводу доброкачественных заболеваний, выявили гораздо более высокую частоту биений ресничек> 10 Гц (Liao et al., 2012). Положительная корреляция между частотой биений ресничек и температурой хорошо известна (Sisson et al. , 2003). Таким образом, это расхождение с нашими выводами можно объяснить, по крайней мере, частично, более высокой температурой 37 ° C, использованной во время измерений в этом исследовании. Подобно предыдущим находкам (Lyons et al. , 2002), мы не наблюдали вариаций частоты или паттерна биений ресничек в разных анатомических участках яйцевода.
PCD - это заболевание подвижных респираторных ресничек, проявляющее значительную генетическую гетерогенность.В настоящее время мутации более чем в 30 различных генах связаны с PCD (Werner et al. , 2015). Фенотипически биение ресничек при PCD можно разделить на варианты со статическими ресничками, варианты с нормальной или повышенной частотой биений, но с аномальным паттерном биения ресничек, и варианты с неспособностью генерировать MMC (Boon et al., , 2014; Raidt et al. , 2014; Wallmeier et al. , 2014). Следует отметить, что биение ресничек всегда очень дискинетично при PCD даже в вариантах с нормальной или повышенной частотой, и поэтому люди с этими подтипами страдают тяжелыми респираторными заболеваниями из-за значительного нарушения мукоцилиарного клиренса (Schwabe et al., 2008; Olbrich et al. , 2012; Wirschell et al. , 2013). Помимо респираторных ресничек, PCD поражает подвижные реснички и в других органах. Дисфункция ресничек эмбрионального узла приводит к рандомизации распределения органов слева и справа, что приводит к situs inversus totalis у 40-50% людей (Lucas et al. , 2014; Werner et al. , 2015) и гетеротаксия у небольшой группы пациентов (Kennedy et al. , 2007; Nakhleh et al., 2012). У мужчин с поражением PCD дискинезия жгутиков сперматозоидов обычно приводит к бесплодию (Fliegauf et al. , 2005, 2007). Хотя хорошо известно, что яйцеводы самок выстланы MMC, роль цилиарной функции FT в репродукции самок плохо изучена. Основываясь на выводе по аналогии, женщины с PCD подозреваются в нарушении фертильности (Afzelius and Eliasson, 1983; McComb et al. , 1986; Lurie et al. , 1989; Halbert et al. , 1997; Lyons ). и другие., 2006). В отличие от этого, мы сообщаем здесь о девяти женщинах с PCD, родивших детей после самопроизвольного зачатия. Это согласуется с предыдущими сообщениями о случаях (Jean et al. , 1979; Eliyahu and Shalev, 1996). У шести из этих девяти самок респираторные реснички лишены компонента ODA, DNAH5. Двуаллельные мутации в DNAH5 , предсказывающие полное отсутствие этого моторного белка, присутствовали у трех человек (OP-118, OP-515 и OP-527 II1). У пациента OP-1795 II3 гомозиготная мутация в гене RSPh5A приводит к отсутствию белков RSPh2, RSPh5A и RSPH9 головки лучевой спицы.Только у пациента OP-2049, женщины с situs inversus totalis, хроническим риносинуситом и бронхоэктатической болезнью легких, мы не смогли продемонстрировать ультраструктурный или генетический дефект из-за отсутствия материала. Взятые вместе, мы здесь показываем, что спонтанное зачатие может происходить у женщин с отсутствием важных моторных белков в аксонеме подвижных ресничек и сильно нарушенным респираторным мукоцилиарным клиренсом. Это указывает на то, что транспорт яйцеклетки к матке, по-видимому, не полностью зависит от транспорта, генерируемого ресничками.
Наше исследование демонстрирует ограничения, которые требуют дальнейшего обсуждения. Поскольку образцы ткани FT от женщин, пораженных PCD, не были доступны для анализа HVMA или IF, мы не смогли продемонстрировать, что реснички FT у женщин PCD действительно имеют тот же дефект, что и респираторные реснички. Таким образом, прямые доказательства того, что реснички FT имеют тот же патогенный дефект, что и респираторные реснички, отсутствуют, даже если наши результаты весьма наводят на размышления. Кроме того, мы не исследовали роль механизмов транспорта, не зависящих от ресничек, таких как мышечные сокращения или состав слизи (Lyons et al., 2006). Биение ресничек обусловлено скоординированной активностью моторных белков. Этот процесс снова регулируется множеством факторов, таких как возраст (Ho et al. , 2001), гормоны (Mahmood et al. , 1998; Jain et al. , 2012; Bylander et al. , 2013 ; O et al. , 2013), кальциевые и циклические нуклеотидные пути (Li et al. , 2000; Schmid and Salathe, 2011), хемокины (Allen-Gipson et al. , 2004; Papathanasiou et al. ., 2008) и раздражителей (Zhou et al. , 2009; Wyatt et al. , 2013). Целью нашего исследования было проанализировать состав моторных белков ресничек FT и результирующее биение ресничек. Мы обнаружили идентичный состав моторных белков по сравнению с респираторными ресничками и, следовательно, очень похожее биение ресничек. Детальный анализ регуляции биений ресничек разными агентами не входил в рамки исследования. Кроме того, наша серия из девяти самок PCD с явно нормальной фертильностью не исключает возможности увеличения субфертильности в этой группе пациентов.Поскольку основная цель нашей базы данных PCD не заключалась в сборе информации о фертильности, полноты данных было недостаточно для более подробного статистического анализа. В конечном итоге потребуются крупные эпидемиологические исследования для дальнейшего определения точной степени женской (суб) фертильности при PCD. Для этой цели и решения других вопросов, касающихся проявления, течения и сопутствующих заболеваний PCD, мы создали международный регистр PCD, который в настоящее время набирает пациентов (www.pcdregistry.eu).
В заключение, мы продемонстрировали, что яйцеводы самок человека выстланы MMC и демонстрируют тот же состав моторных белков ресничек и паттерн биения, что и дыхательные MMC.Активность MMC в FT приводит к скоординированному транспорту частиц к матке. Однако, по крайней мере, часть людей с тяжелой дисфункцией подвижных ресничек может зачать ребенка спонтанно, что указывает на то, что нарушение функции ресничек может быть компенсировано другими механизмами. Необходимы эпидемиологические исследования, чтобы выяснить точную степень снижения фертильности у этих женщин.
Авторские роли
Х. Омран и К.В. разработали исследование. C.W. и J.R. составили рукопись и подготовили рисунки.Р.С. предоставил образцы FT. C.W. и J.R. предоставили клинические данные. J.R. и C.W. провели эксперименты HVMA. J.R., N.T.L. и П. провели эксперименты IF. G.W.D. и N.T.L. участвовал в интерпретации данных и написании статьи. J.R., P.P. и Т. провели и проанализировали эксперименты с функцией транспорта ресничек. Х. Ольбрих оказал помощь в составлении рукописи и рисунков. Все авторы прочитали и критически отредактировали рукопись и предоставили окончательное одобрение для публикации.
Финансирование
Эта работа была поддержана грантами Европейской комиссии FP7 (Седьмая рамочная программа исследований) 305404 (BESTCILIA) и 241955 (SYSCILIA) Х. Омрану, Deutsche Forschungsgemeinschaft (DFG OM 6/4, OM 6/5) и IZKF Muenster (Om2 / 009/12).
Конфликт интересов
Не объявлено.
Благодарности
Мы благодарны людям, страдающим PCD, за их участие, и мы благодарим немецкую группу поддержки пациентов «Kartagener Syndrom und Primaere Ciliaere Dyskinesie e.V. ’. Мы благодарим M. Herting, L. Overkamp, K. Vorspohl и S. Helms за отличную техническую помощь.
Список литературы
Афзелиус
BA
.
Человеческий синдром, вызванный неподвижными ресничками
.
Наука
1976
;
193
:
317
-
319
.
Афзелиус
BA
.
Заболевания, связанные с ресничками
.
Дж Патол
2004
;
204
:
470
-
477
.
Афзелиус
В
,
Элиассон
R
.
Проблемы мужского и женского бесплодия при синдроме неподвижных ресничек
.
евро J Respir Dis Suppl
1983
;
127
:
144
-
147
.
Аллен-Гипсон
DS
,
Romberger
DJ
,
Forget
MA
,
May
KL
,
Sisson
JH
,
Wyatt
TA
.
ИЛ-8 подавляет стимулированную изопротеренолом частоту биений ресничек в эпителиальных клетках бронхов крупного рогатого скота
.
J Aerosol Med
2004
;
17
:
107
-
115
.
Boon
м
,
Wallmeier
J
,
Ma
L
,
Loges
NT
,
Jaspers
M
,
Olbrich
H
,
Dougherty
000
0003
Вернер
C
,
Amirav
I
et al. .
Мутации MCIDAS приводят к нарушению мукоцилиарного клиренса с уменьшением образования множественных подвижных ресничек
.
Нац Коммуна
2014
;
5
:
4418
.
Биландер
А
,
Lind
K
,
Goksör
M
,
Billig
H
,
Larsson
DJ
.
Классический рецептор прогестерона опосредует быстрое снижение частоты биений ресничек маточных труб с помощью прогестерона
.
Репрод Биол Эндокринол
2013
;
11
:
33
.
Ворона
Дж
,
Amso
NN
,
Lewin
J
,
Shaw
RW
.
Морфология и ультраструктура эпителия маточных труб на разных стадиях менструального цикла и менопаузы
.
Hum Reprod
1994
;
9
:
2224
-
2233
.
Дирксен
ER
,
Сатир
П
.
Активность ресничек в яйцеводе мыши по данным трансмиссионной и сканирующей электронной микроскопии
.
Тканевая клетка
1972
;
4
:
389
-
403
.
Элиягу
S
,
Шалев
Е
.
Фертильная женщина с синдромом Картагенера и тремя беременностями подряд
.
Hum Reprod
1996
;
11
:
683
.
Ezzati
м
,
Джаханбахч
О
,
Ариан
S
,
Карр
BR
.
Трубный транспорт гамет и эмбрионов: обзор физиологии и патофизиологии
.
J Assist Reprod Genet
2014
;
31
:
1337
-
1347
.
Fliegauf
м
,
Olbrich
H
,
Horvath
J
,
Wildhaber
JH
,
Zariwala
MA
,
Kennedy
M
,
000 MR
Неправильная локализация ДНКН5 и ДНКН9 в респираторных клетках пациентов с первичной цилиарной дискинезией
.
Am J Respir Crit Care Med
2005
;
171
:
1343
-
1349
.
Fliegauf
м
,
Benzing
T
,
Omran
H
.
Когда портятся реснички: дефекты ресничек и цилиопатии
.
Nat Rev Mol Cell Biol
2007
;
8
:
880
-
893
.
Халберт
SA
,
Tam
PY
,
Blandau
RJ
.
Транспортировка яиц в яйцеводе кролика: роль ресничек и мышц
.
Наука
1976
;
191
:
1052
-
1053
.
Халберт
SA
,
Patton
DL
,
Zarutskie
PW
,
Soules
MR
.
Функция и строение ресничек маточной трубы бесплодной женщины с синдромом Картагенера
.
Hum Reprod
1997
;
12
:
55
-
58
.
Hjeij
R
,
Onoufriadis
A
,
Watson
CM
,
Slagle
CE
,
Klena
NT
,
Dougherty
GW
9000 9000 9000 M000 Loges3
Diggle
CP
,
Morante
NFC
et al..
Мутации CCDC151 вызывают первичную дискинезию ресничек за счет нарушения образования стыковочного комплекса внешнего плеча динеина
.
Am J Hum Genet
2014
;
95
:
257
-
274
.
Ho
JC
,
Chan
KN
,
Hu
WH
,
Lam
WK
,
Zheng
L
,
Tipoe
GL
,
Sun
J
Цанг
кВт
.
Влияние старения на мукоцилиарный клиренс носа, частоту ударов и ультраструктуру респираторных ресничек
.
Am J Respir Crit Care Med
2001
;
163
:
983
-
988
.
Ибаньес-Таллон
Я
,
Heintz
N
,
Omran
H
.
Бить или не бить: роль ресничек в развитии и болезни
.
Hum Mol Genet
2003
;
12
:
R27
-
R35
.
Джайн
R
,
Ray
JM
,
Pan
JH
,
Brody
SL
.
Зависимая от половых гормонов регуляция частоты биений ресничек в эпителии дыхательных путей
.
Am J Respir Cell Mol Biol
2012
;
46
:
446
-
453
.
Жан
Я
,
Langlais
J
,
Roberts
KD
,
Chapdelaine
A
,
Bleau
G
.
Фертильность женщины с нефункциональными ресничными клетками маточных труб
.
Fertil Steril
1979
;
31
:
349
-
350
.
Кеннеди
MP
,
Omran
H
,
Leigh
MW
,
Dell
S
,
Morgan
L
,
Molina
PL
,
Robinson
BV
Ольбрих
H
,
Северин
T
et al. .
Врожденные пороки сердца и другие гетеротаксические дефекты в большой группе пациентов с первичной цилиарной дискинезией
.
В обращении
2007
;
115
:
2814
-
2821
.
Ли
D
,
Шираками
G
,
Жан
X
,
Johns
RA
.
Регулирование частоты биений ресничек с помощью сигнального пути оксида азота и циклического гуанозинмонофосфата в эпителиальных клетках дыхательных путей крысы
.
Am J Respir Cell Mol Biol
2000
;
23
:
175
-
181
.
Ляо
SB
,
Li
HWR
,
Ho
JC
,
Yeung
WSB
,
Ng
EHY
,
Cheunga
NY
,
Tang
Tang
Возможная роль адреномедуллина в патогенезе трубной внематочной беременности
.
J Clin Endocrinol Metab
2012
;
97
:
2105
-
2112
.
Логи
NT
,
Olbrich
H
,
Fenske
L
,
Mussaffi
H
,
Horvath
J
,
Fliegauf
M
,
Kuhl
Петерффи
E
,
Chodhari
R
et al..
Мутации DNAI2 вызывают первичную цилиарную дискинезию с дефектами внешнего плеча динеина
.
Am J Hum Genet
2008
;
83
:
547
-
558
.
Логи
NT
,
Olbrich
H
,
Becker-Heck
A
,
Häffner
K
,
Heer
A
,
Reinhard
C
,
000 M0002000 M0003
000 M0003
,
Заривала
MA
,
Leigh
МВт
et al..
Делеции и точечные мутации LRRC50 вызывают первичную цилиарную дискинезию из-за дефектов динеинового плеча
.
Am J Hum Genet
2009
;
85
:
883
-
889
.
Лукас
JS
,
Burgess
A
,
Mitchison
HM
,
Moya
E
,
Williamson
M
,
Hogg
C
.
Диагностика и лечение первичной цилиарной дискинезии
.
Arch Dis Child
2014
;
99
:
850
-
856
.
Лурье
м
,
Тур-Каспа
I
,
Weill
S
,
Katz
I
,
Rabinovici
J
,
Goldenberg
S
.
Цилиарная ультраструктура эпителия дыхательных путей и маточных труб у бесплодной женщины с синдромом Картагенера. Количественная оценка
.
Комод
1989
;
95
:
578
-
581
.
Лион
RA
,
Джаханбахч
O
,
Махмуд
T
,
Саридоган
E
,
Sattar
S
,
Sheaff
MT
000
oy
oy
oy
Частота биений ресничек маточной трубы в зависимости от стадии менструального цикла и анатомического участка
.
Hum Reprod
2002
;
17
:
584
-
588
.
Лион
RA
,
Саридоган
E
,
Джаханбахч
O
.
Репродуктивное значение ресничек фаллопиевых труб человека
.
Обновление Hum Reprod
2006
;
12
:
363
-
372
.
Махмуд
т
,
Саридоган
E
,
Смутна
S
,
Хабиб
AM
,
Джаханбахч
O
.
Влияние стероидов яичников на частоту сердцебиения эпителия фаллопиевых труб человека
.
Hum Reprod
1998
;
13
:
2991
-
2994
.
МакКомб
-П
,
Лэнгли
L
,
Villalon
M
,
Verdugo
P
.
Реснички яйцевода и синдром Картагенера
.
Fertil Steril
1986
;
46
:
412
-
416
.
Merveille
A-C
Ул.
,
Billen
F
,
Clément
A
et al. .
CCDC39 необходим для сборки внутренних плеч динеина и регуляторного комплекса динеина, а также для нормальной перистальтики ресничек у людей и собак
.
Нат Генет
2011
;
43
:
72
-
78
.
Nakhleh
N
,
Francis
R
,
Giese
RA
,
Tian
X
,
Li
Y
,
Zariwala
MA
,
Yagi
hal
Hagi
Куреши
S
,
Chatterjee
B
et al..
Высокая распространенность респираторной цилиарной дисфункции у пациентов с врожденными пороками сердца и гетеротаксией
.
В обращении
2012
;
125
:
2232
-
2242
.
O
WS
,
Li
HWR
,
Liao
SB
,
Cheung
ANY
,
Ng
EHY
,
Yeung
WSB
,
Ho
9000 Tang
Снижение экспрессии адреномедуллина и частоты биений ресничек в носовом эпителии при трубной беременности
.
Fertil Steril
2013
;
100
:
459
-
463.e1
. .
О'Тул
ET
,
Гиддингс
TH
,
Портер
ME
,
Островски
LE
.
Компьютерный анализ изображений ресничек человека и жгутиков хламидомонады выявляет как сходства, так и различия в структуре аксонемы
.
Цитоскелет (Хобокен)
2012
;
69
:
577
-
590
.
Ольбрих
H
,
Schmidts
M
,
Werner
C
,
Onoufriadis
A
,
Loges
NT
,
Raidt
J
,
Banki
,
Banki
,
Banki
Burgoyne
T
,
Al Turki
S
et al..
Рецессивные мутации HYDIN вызывают первичную цилиарную дискинезию без рандомизации лево-правой асимметрии тела
.
Am J Hum Genet
2012
;
91
:
672
-
684
.
Омран
H
,
Логи
NT
.
Иммунофлуоресцентное окрашивание мерцательных клеток респираторного эпителия
.
Методы Cell Biol
2009
;
91
:
123
-
133
.
Папатанасиу
А
,
Джаханбахч
O
,
Саридоган
E
,
Lyons
RA
.
Влияние интерлейкина-6 на частоту биений ресничек в фаллопиевых трубах человека
.
Fertil Steril
2008
;
90
:
391
-
394
.
Рейдт
Дж
,
Wallmeier
J
,
Hjeij
R
,
Onnebrink
G
,
Pennekamp
P
,
Loges
NT
,
000 Hjeij
0003
Догерти
GW
,
Омран
H
et al. .
Характер и частота биений ресничек в генетических вариантах первичной дискинезии ресничек
.
евро Респир J
2014
;
44
:
1579
-
1588
.
Сатир
-П
.
Механизмы движения ресничек: материалы электронной микроскопии
.
Scanning Microsc
1992
;
6
:
573
-
579
.
Шинделин
Дж
Ул.
,
Saalfeld
S
,
Schmid
B
et al..
Фиджи: платформа с открытым исходным кодом для анализа биологических изображений
.
Nat Methods
2012
;
9
:
676
-
682
.
Шмид
А
,
Салате
М
.
Координация биений ресничек кальцием
.
Biol Cell
2011
;
103
:
159
-
169
.
Швабе
GC
,
Hoffmann
K
,
Loges
NT
,
Birker
D
,
Rossier
C
,
de Santi
MM
,
Flaubrich
,
Не удалось
M
,
Liebers
U
et al. .
Первичная дискинезия ресничек, связанная с нормальной ультраструктурой аксонемы, вызывается мутациями ДНКh21
.
Хум Мутат
2008
;
29
:
289
-
298
.
Сиссон
JH
,
Stoner
JA
,
Ammons
BA
,
Wyatt
TA
.
Захват полностью цифровых изображений и анализ частоты биений ресничек по всему полю
.
J Microsc
2003
;
211
:
103
-
111
.
Валмайер
Дж
,
Аль-Мутаири
DA
,
Чен
CT
,
Loges
NT
,
Pennekamp
P
,
Menchen
T
,
HE
Ma
Ma
Ma
,
Olbrich
H
,
Dougherty
GW
et al..
Мутации в CCNO приводят к врожденному нарушению мукоцилиарного клиренса с уменьшением образования множественных подвижных ресничек
.
Нат Генет
2014
;
46
:
646
-
651
. .
Вернер
С
,
Große Onnebrink
J
,
Omran
H
.
Диагностика и лечение первичной цилиарной дискинезии
.
Реснички
2015
;
4
:
1
-
9
.
Wirschell
м
,
Olbrich
H
,
Werner
C
,
Tritschler
D
,
Bower
R
,
Продажа
amp WS
,
Loges
NT2000 Пен
NT2000
Lindberg
S
,
Stenram
U
et al..
Субъединица DRC1 регуляторного комплекса нексин-динеин важна для функции подвижных ресничек у водорослей и людей
.
Нат Генет
2013
;
45
:
262
-
268
.
Вятт
TA
,
Wells
SM
,
Alsaidi
ZA
,
Devasure
JM
,
Klein
EB
,
Bailey
KL
,
000
.
Асимметричный диметиларгинин блокирует опосредованное оксидом азота стимулированное спиртом биение ресничек
.
Медиаторы воспаления
2013
;
2013
:
1
-
9
.
Заривала
MA
,
Leigh
MW
,
Ceppa
F
,
Kennedy
MP
,
Noone
PG
,
Carson
JL
,
Hazucha
000 Mori
Хорват
J
,
Ольбрих
H
et al..
Мутации ДНКI1 при первичной цилиарной дискинезии: свидетельство эффекта основателя в общей мутации
.
Am J Respir Crit Care Med
2006
;
174
:
858
-
866
.
Чжоу
H
,
Wang
X
,
Brighton
L
,
Hazucha
M
,
Jaspers
I
,
Carson
JL
.
Повышенная частота пульсации цилиарного эпителия носа, связанная с воздействием табачного дыма в образе жизни
.
Токсикол для вдыхания
2009
;
21
:
875
-
881
.
Заметки автора
© Автор, 2015. Опубликовано Oxford University Press от имени Европейского общества репродукции человека и эмбриологии. Все права защищены. Для получения разрешений, пожалуйста, напишите в журналы[email protected]
курсов английского языка | Общественный колледж округа Джонсон
Четкое и лаконичное общение - ключ к успеху в любой карьере. Отточите свои навыки редактирования и редактирования на курсах английского языка JCCC.
Что я узнаю на курсах английского языка JCCC?
Пройдите курсы английского языка в JCCC, чтобы научиться лучше общаться, стать более сильным писателем и разносторонним читателем, размышляющим о человеческом опыте.
Последовательность составления материалов Департамента английского языка JCCC закладывает основу для письменного изложения аргументов.Вы откроете для себя дисциплину, которая является наследницей почти 3000 лет литературы и искусства риторики. У вас будет возможность заявить о себе в письменной форме в JCCC.
Успешное завершение Композиций I и II требуется для многих программ на получение степени и сертификата в JCCC. Вы узнаете все этапы процесса написания, от поиска идей и обоснования своих позиций кратким английским языком до редактирования, проверки и защиты вашего окончательного документа.
Разделы доступны в соответствии с вашим расписанием, с временем начала уже с 7 часов утра.м. и уже в 18:00. Мы предлагаем как полные семестровые, так и краткосрочные уроки композиции. Онлайн-курсы английского языка предлагают еще большую гибкость.
От Библии как литературы до литературы американской поп-музыки, от Гомера и Шекспира до научной фантастики и детской литературы - вы научитесь понимать точки зрения со всех сторон.
Вводный курс литературы для первокурсников дает вам обзор поэзии, художественной литературы и драмы. Курсы уровня 200 рассматривают литературу с различных точек зрения, в том числе:
- Жанр - с такими курсами, как «Введение в художественную литературу», «Введение в поэзию», «Драма как литература» и «Шедевры кино».
- География - с американской литературой I и II, британской литературой I и II и мировыми шедеврами
- Поп-культура - через литературу научной фантастики и литературу американской поп-музыки
- Разнообразие - с литературой женщин, U.С. Латино и Латинская литература и Библия как литература
[Каталог] Курсы английского
Почему курсы английского языка при JCCC?
Неотъемлемая часть обучения в колледже - это четкое письмо и включение исследований в ваши письменные задания. Курсы английского языка JCCC охватывают весь спектр письменных работ на уровне колледжа, от составления эффективных предложений до создания новаторских исследовательских проектов, как технических, так и творческих.
The Mind’s Eye, студенческий литературный журнал, дает вам возможность публиковать художественную литературу, стихи и эссе, а также произведения искусства и фотографии.Для получения дополнительной информации свяжитесь с профессором Томом Рейнольдсом, консультантом факультета The Mind’s Eye, по телефону 913-469-8500, доб. 3935.
Преподаватели кафедры английского языка
JCCC отличаются интеллектуальным любопытством, творческими усилиями, приверженностью студентам и дисциплине, а также непрекращающимся стремлением соединить местное с универсальным, субъективное с объективным, меня с нами и нас. им.
Совокупный опыт преподавания на нашем факультете в колледжах буквально простирается на века.Нам посчастливилось собрать сообщество экспертов не только по композиции и риторике, но и по широкому кругу литературных, писательских и кинематографических интересов.
Мы очень гордимся тем, что являемся неотъемлемой частью успеха учащихся еще долгое время после того, как учащийся прошел свой последний урок Comp.