3.1. Пирамидная система. Неврология и нейрохирургия
3.1. Пирамидная система
Различают два основных вида движений: непроизвольные и произвольные.
К непроизвольным относятся простые автоматические движения, осуществляемые за счет сегментарного аппарата спинного мозга и мозгового ствола по типу простого рефлекторного акта. Произвольные целенаправленные движения – акты двигательного поведения человека. Специальные произвольные движения (поведенческие, трудовые и др.) осуществляются при ведущем участии коры большого мозга, а также экстрапирамидной системы и сегментарного аппарата спинного мозга. У человека и высших животных осуществление произвольных движений связано с пирамидной системой. При этом проведение импульса из коры большого мозга к мышце происходит по цепи, состоящей из двух нейронов: центрального и периферического.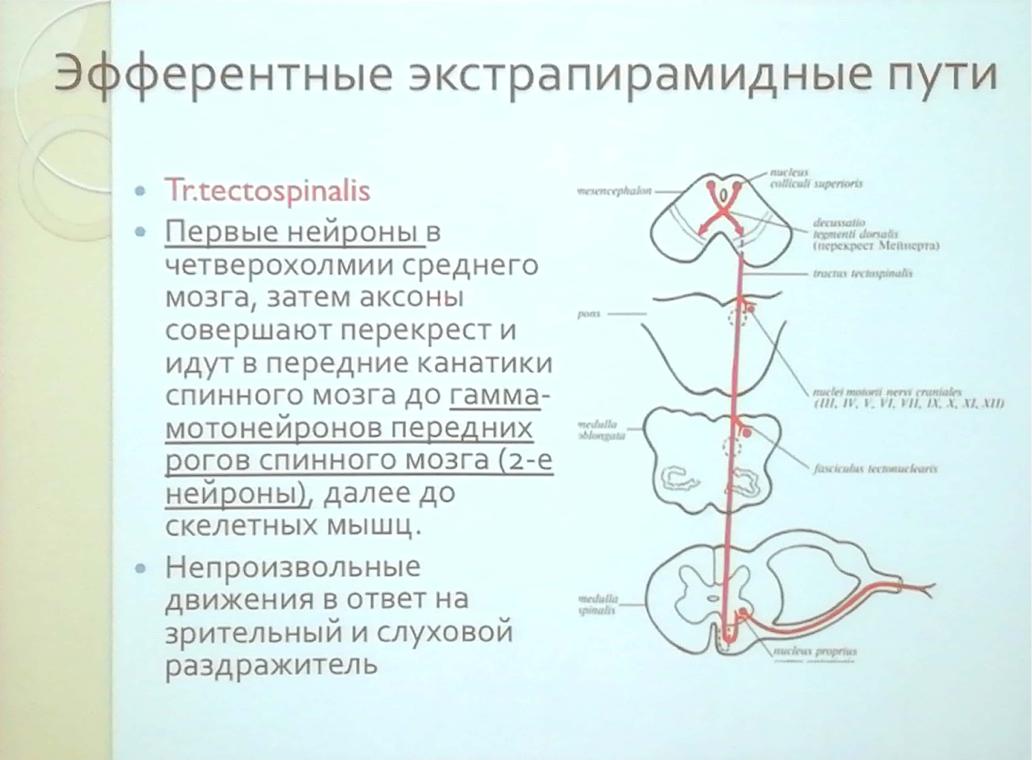
Центральный мотонейрон. Произвольные движения мышц происходят за счет импульсов, идущих по длинным нервным волокнам из коры большого мозга к клеткам передних рогов спинного мозга. Эти волокна формируют двигательный (корково-спинномозговой), или пирамидный, путь. Они являются аксонами нейронов, расположенных в прецентральной извилине, в цитоархитектоническом поле 4. Эта зона представляет собой узкое поле, которое тянется вдоль центральной щели от латеральной (или сильвиевой) борозды к передней части парацентральной дольки на медиальной поверхности полушария, параллельно чувствительной области коры постцентральной извилины.
Нейроны, иннервирующие глотку и гортань, расположены в нижней части прецентральной извилины. Далее в восходящем порядке идут нейроны, иннервирующие лицо, руку, туловище, ногу. Таким образом, все участки тела человека спроецированы в прецентральной извилине как бы вверх ногами. Мотонейроны расположены не только в поле 4, они встречаются и в соседних кортикальных полях. В то же время, подавляющее большинство их занимает 5-й кортикальный слой 4-го поля. Они «ответственны» за точные, нацеленные единичные движения. Эти нейроны включают и гигантские пирамидные клетки Беца, имеющие аксоны с толстой миелиновой оболочкой. Эти быстропроводящие волокна составляют лишь 3,4—4 % от всех волокон пирамидного пути. Большинство волокон пирамидного пути исходит из малых пирамидных, или фузиформных (веретенообразных), клеток в двигательных полях 4 и 6. Клетки поля 4 дают около 40 % волокон пирамидного пути, остальные исходят из клеток других полей сенсомоторной зоны.
В то же время, подавляющее большинство их занимает 5-й кортикальный слой 4-го поля. Они «ответственны» за точные, нацеленные единичные движения. Эти нейроны включают и гигантские пирамидные клетки Беца, имеющие аксоны с толстой миелиновой оболочкой. Эти быстропроводящие волокна составляют лишь 3,4—4 % от всех волокон пирамидного пути. Большинство волокон пирамидного пути исходит из малых пирамидных, или фузиформных (веретенообразных), клеток в двигательных полях 4 и 6. Клетки поля 4 дают около 40 % волокон пирамидного пути, остальные исходят из клеток других полей сенсомоторной зоны.
Мотонейроны поля 4 контролируют тонкие произвольные движения скелетных мышц противоположной половины тела, так как большинство пирамидных волокон переходит на противоположную сторону в нижней части продолговатого мозга.
Импульсы пирамидных клеток двигательной области коры идут по двум путям. Один – корково-ядерный путь – оканчивается в ядрах черепных нервов, второй, более мощный, корково-спинномозговой – переключается в переднем роге спинного мозга на вставочных нейронах, которые в свою очередь оканчиваются на больших мотонейронах передних рогов. Эти клетки передают импульсы через передние корешки и периферические нервы к двигательным концевым пластинкам скелетной мускулатуры.
Эти клетки передают импульсы через передние корешки и периферические нервы к двигательным концевым пластинкам скелетной мускулатуры.
Когда волокна пирамидного пути покидают двигательную область коры, они, проходя через лучистый венец белого вещества мозга, сходятся по направлению к задней ножке внутренней капсулы. В соматотопическом порядке они проходят внутреннюю капсулу (ее колено и передние две трети заднего бедра) и идут в средней части ножек мозга, нисходят через каждую половину основания моста, будучи окруженными многочисленными нервными клетками ядер моста и волокнами различных систем. На уровне понтомедуллярного сочленения пирамидный путь становится видимым извне, его волокна образуют удлиненные пирамиды по обе стороны от средней линии продолговатого мозга (отсюда его название). В нижней части продолговатого мозга 80—85 % волокон каждого пирамидного пути переходят на противоположную сторону в перекресте пирамид и образуют латеральный пирамидный путь. Остальные волокна продолжают спускаться неперекрещенными в передних канатиках в качестве переднего пирамидного пути.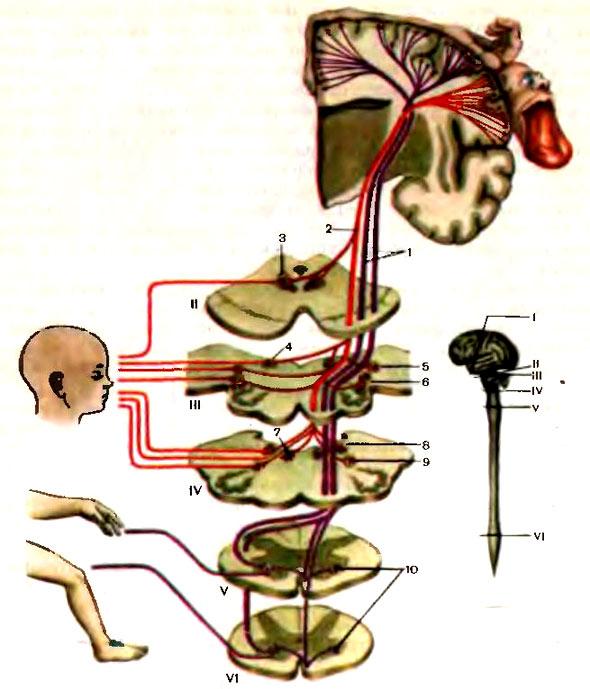 Эти волокна перекрещиваются на сегментарном уровне через переднюю комиссуру спинного мозга. В шейной и грудной частях спинного мозга некоторые волокна соединяются с клетками переднего рога своей стороны, так что мышцы шеи и туловища получают кортикальную иннервацию с обеих сторон.
Эти волокна перекрещиваются на сегментарном уровне через переднюю комиссуру спинного мозга. В шейной и грудной частях спинного мозга некоторые волокна соединяются с клетками переднего рога своей стороны, так что мышцы шеи и туловища получают кортикальную иннервацию с обеих сторон.
Перекрещенные волокна спускаются в составе латерального пирамидного пути в латеральных канатиках. Около 90 % волокон образуют синапсы со вставочными нейронами, которые в свою очередь соединяются с большими альфа– и гамма-нейронами переднего рога спинного мозга.
Волокна, формирующие корково-ядерный путь, направляются к двигательным ядрам (V, VII, IX, X, XI, XII) черепных нервов и обеспечивают произвольную иннервацию лицевой и оральной мускулатуры.
Заслуживает внимания и другой пучок волокон, начинающийся в «глазном» поле 8, а не в прецентральной извилине. Импульсы, идущие по этому пучку, обеспечивают содружественные движения глазных яблок в противоположную сторону. Волокна этого пучка на уровне лучистого венца присоединяются к пирамидному пути. Затем они проходят более вентрально в задней ножке внутренней капсулы, поворачивают каудально и идут к ядрам III, IV, VI черепных нервов.
Затем они проходят более вентрально в задней ножке внутренней капсулы, поворачивают каудально и идут к ядрам III, IV, VI черепных нервов.
Периферический мотонейрон. Волокна пирамидного пути и различных экстрапирамидных путей (ретикулярно-, покрышечно-, преддверно-, красноядерно-спинномозгового и др.) и афферентные волокна, входящие в спинной мозг через задние корешки, оканчиваются на телах или дендритах больших и малых альфа– и гамма-клеток (непосредственно либо через вставочные, ассоциативные или комиссуральные нейроны внутреннего нейронального аппарата спинного мозга) В противоположность псевдоуниполярным нейронам спинномозговых узлов нейроны передних рогов мультиполярны. Их дендриты имеют множественные синаптические связи с различными афферентными и эфферентными системами. Некоторые из них – облегчающие, другие – тормозящие по своему действию. В передних рогах мотонейроны образуют группы, организованные в колонки и не разделенные сегментарно. В этих колонках имеется определенный соматотопический порядок. В шейной части латеральные мотонейроны переднего рога иннервируют кисть и руку, а мотонейроны медиальных колонок – мышцы шеи и груди. В поясничной части нейроны, иннервирующие стопу и ногу, также расположены латерально в переднем роге, а иннервирующие туловище – медиальное. Аксоны клеток передних рогов выходят из спинного мозга вентрально как корешковые волокна, которые собираются по сегментам и образуют передние корешки. Каждый передний корешок соединяется с задним дистальнее спинномозговых узлов и вместе они образуют спинномозговой нерв. Таким образом, каждый сегмент спинного мозга имеет свою пару спинномозговых нервов.
В шейной части латеральные мотонейроны переднего рога иннервируют кисть и руку, а мотонейроны медиальных колонок – мышцы шеи и груди. В поясничной части нейроны, иннервирующие стопу и ногу, также расположены латерально в переднем роге, а иннервирующие туловище – медиальное. Аксоны клеток передних рогов выходят из спинного мозга вентрально как корешковые волокна, которые собираются по сегментам и образуют передние корешки. Каждый передний корешок соединяется с задним дистальнее спинномозговых узлов и вместе они образуют спинномозговой нерв. Таким образом, каждый сегмент спинного мозга имеет свою пару спинномозговых нервов.
В состав нервов входят также эфферентные и афферентные волокна, исходящие из боковых рогов спинномозгового серого вещества.
Хорошо миелинизированные, быстропроводящие аксоны больших альфа-клеток идут непосредственно к поперечно-полосатой мускулатуре.
Помимо больших и малых альфа-мотонейронов, передние рога содержат многочисленные гамма-мотонейроны. Среди вставочных нейронов передних рогов следует отметить клетки Реншо, тормозящие действие больших мотонейронов.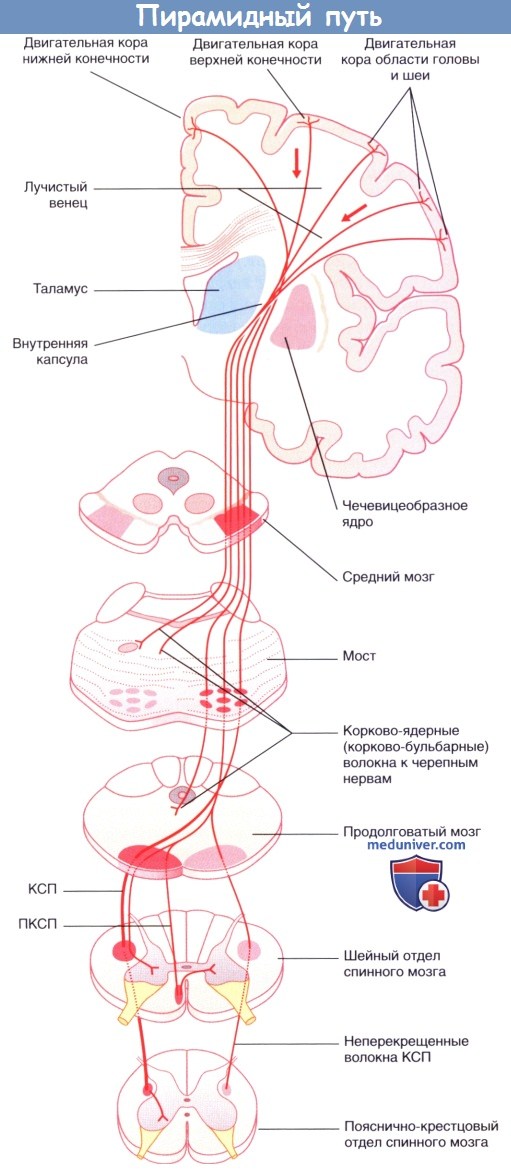 Большие альфа-клетки с толстым и быстропроводящим аксоном осуществляют быстрые сокращения мышц. Малые альфа-клетки с более тонким аксоном выполняют тоническую функцию. Гамма-клетки с тонким и медленнопроводящим аксоном иннервируют проприорецепторы мышечного веретена. Большие альфа-клетки связаны с гигантскими клетками коры полушарий большого мозга. Малые альфа-клетки имеют связь с экстрапирамидной системой. Через гамма-клетки происходит регуляция состояния мышечных проприорецепторов. Среди различных мышечных рецепторов наиболее важными являются нервно-мышечные веретена.
Большие альфа-клетки с толстым и быстропроводящим аксоном осуществляют быстрые сокращения мышц. Малые альфа-клетки с более тонким аксоном выполняют тоническую функцию. Гамма-клетки с тонким и медленнопроводящим аксоном иннервируют проприорецепторы мышечного веретена. Большие альфа-клетки связаны с гигантскими клетками коры полушарий большого мозга. Малые альфа-клетки имеют связь с экстрапирамидной системой. Через гамма-клетки происходит регуляция состояния мышечных проприорецепторов. Среди различных мышечных рецепторов наиболее важными являются нервно-мышечные веретена.
Афферентные волокна, называемые кольцеспиральными, или первичными, окончаниями, имеют довольно толстое миелиновое покрытие и относятся к быстропрово
Пирамидная система. Блог Лого-Эксперт
Пирамидная система — это система эфферентных нейронов, тела которых располагаются в коре большого мозга, оканчиваются в двигательных ядрах черепных нервов и сером веществе спинного мозга. В составе пирамидного пути (tractus pyramidalis) выделяют корково-ядерные волокна (fibrae corticonucleares) и корково-спинномозговые волокна (fibrae corticospinales).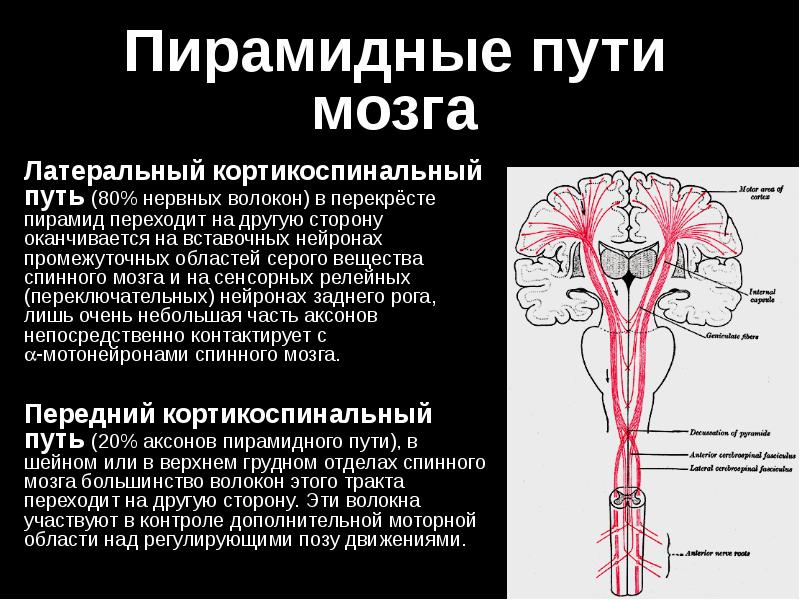 И те, и другие являются аксонами нервных клеток внутреннего, пирамидного, слоя коры большого мозга.
И те, и другие являются аксонами нервных клеток внутреннего, пирамидного, слоя коры большого мозга.
Они располагаются в предцентральной извилине и прилегающих к ней полях лобной и теменной долей. В предцентральной извилине локализуется первичное двигательное поле, где располагаются пирамидные нейроны, управляющие отдельными мышцами и группами мышц. В этой извилине существует соматотопическое представительство мускулатуры.
Нейроны, управляющие мышцами глотки, языка и головы, занимают нижнюю часть извилины; выше располагаются участки, связанные с мышцами верхней конечности и туловища; проекция мускулатуры нижней конечности находится в верхней части предцентральной извилины и переходит на медиальную поверхность полушария.
Пирамидный путь образуют преимущественно тонкие нервные волокна, которые проходят в белом веществе полушария и конвергируют к внутренней капсуле.
Корково-ядерные волокна формируют колено, а корково-спинномозговые волокна — передние 2/3 задней ножки внутренней капсулы. Отсюда пирамидный путь продолжается в основание ножки мозга и далее в переднюю часть моста.
Отсюда пирамидный путь продолжается в основание ножки мозга и далее в переднюю часть моста.
На протяжении ствола мозга корково-ядерные волокна переходят на противоположную сторону к дорсолатеральным участкам ретикулярной формации, где они переключаются на двигательные ядра III, IV, V, VI, VII, IX, X, XI, XII черепных нервов; только к верхней трети ядра лицевого нерва идут неперекрещенные волокна. Часть волокон пирамидного пути проходит из ствола головного мозга в мозжечок.
В продолговатом мозге пирамидный путь располагается в пирамидах, которые на границе со спинным мозгом образуют перекрест (decussatio pyramidum). Выше перекреста пирамидный путь содержит от 700 000 до 1 300 000 нервных волокон с одной стороны. В результате перекреста 80% волокон переходит на противоположную сторону и образует в боковом канатике спинного мозга латеральный корково-спинномозговой (пирамидный) путь. Не перекрещенные волокна из продолговатого мозга продолжаются в передний канатик спинного мозга в виде переднего корково-спинномозгового (пирамидного) пути.
Волокна этого пути переходят на противоположную сторону на протяжении спинного мозга в его белой спайке (посегментно).
Большинство корково-спинномозговых волокон оканчивается в промежуточном сером веществе спинного мозга на его вставочных нейронах, лишь часть их образует синапсы непосредственно с двигательными нейронами передних рогов, которые дают начало двигательным волокнам спинномозговых нервов. В шейных сегментах спинного мозга оканчивается около 55% корково-спинномозговых волокон, в грудных сегментах 20% и в поясничных сегментах 25%.
Передний корково-спинномозговой путь продолжается только до средних грудных сегментов. Благодаря перекресту волокон в пирамидной системы левое полушарие головного мозга управляет движениями правой половины тела, а правое полушарие — движениями левой половины тела, однако мышцы туловища и верхней трети лица получают волокна пирамидного пути из обоих полушарий.
Функция пирамидной системы состоит в восприятии программы произвольного движения и проведении импульсов этой программы до сегментарного аппарата ствола головного и спинного мозга.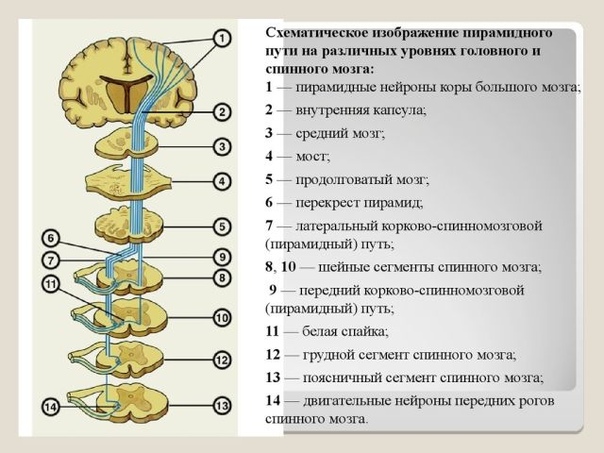
В клинической практике состояние пирамидной системы определяют по характеру произвольных движений.
Оценивают объем движений и силу сокращения поперечнополосатых мышц по шестибалльной системе (полная сила мышц — 5 баллов, «уступчивость» мышечной силы — 4 балла, умеренное снижение силы при полном объеме активных движений — 3 балла, возможность полного объема движений только после относительного устранения силы тяжести конечности — 2 балла, сохранность шевеления с едва заметным сокращением мышцы — 1 балл и отсутствие произвольного движения — 0).
Оценить силу сокращения мышц количественно можно с помощью динамометра. Для оценки сохранности пирамидного корково-ядерного пути к двигательным ядрам черепных нервов используют тесты, с помощью которых определяют функцию мышц головы и шеи, иннервируемых этими ядрами, кортикоспинального тракта — при исследовании мышц туловища и конечностей. О поражении пирамидной системы судят также по состоянию мышечного тонуса и трофике мышц.
Пирамидная система — Карта знаний
- Пирамидная система, пирамидный путь (лат. tractus pyramidalis, PNA) — система нервных структур. Поддерживает сложную и тонкую координацию движений.
Пирамидная система — одно из поздних приобретений эволюции. Низшие позвоночные пирамидальной системы не имеют, она появляется только у млекопитающих, и достигает наибольшего развития у обезьян и особенно у человека. Пирамидная система играет особую роль в прямохождении.
Источник: Википедия
Связанные понятия
Пирамидные знаки — группа патологических рефлексов, которые возникают вследствие поражения центрального нейрона коры головного мозга, либо кортикоспинальных или кортиконуклеарных проводящих путей, идущих соответственно к мотонейронам спинного мозга или двигательным ядрам черепных нервов.
Периферическая нервная система — условно выделяемая часть нервной системы, находящаяся за пределами головного и спинного мозга. Она состоит из черепных и спинальных нервов, а также нервов и сплетений вегетативной нервной системы, соединяя центральную нервную систему с органами тела.
Преддверно-улитковый нерв (лат. nervus vestibulocochlearis) — (VIII пара черепных нервов) нерв специальной чувствительности, отвечающий за передачу слуховых импульсов, а также импульсов, исходящих из вестибулярного отдела внутреннего уха.
Нерв (лат. nervus) — составная часть нервной системы; покрытая оболочкой структура, состоящая из сплетения пучков нервных волокон (главным образом, представленных аксонами нейронов и поддерживающей их нейроглии), обеспечивающее передачу сигналов между головным и спинным мозгом и органами. Совокупность всех нервов организма образует периферическую нервную систему. Соседние нервы могут образовывать нервные сплетения. Крупные нервы называются нервными стволами. Дальше от мозга нервы разветвляются, в…
Соседние нервы могут образовывать нервные сплетения. Крупные нервы называются нервными стволами. Дальше от мозга нервы разветвляются, в…
Прецентральная извилина (лат. gyrus precentralis) — участок лобной доли коры больших полушарий головного мозга. В ней начинается пирамидный путь, который, заканчиваясь на мотонейронах спинного мозга и двигательных ядрах черепных нервов, обеспечивает сознательные движения. Функционально соответствует первичной моторной коре, цитоархитектонически — полю Бродмана 4.
Упоминания в литературе
Произвольные движения – это акты двигательного поведения человека (праксии). Они осуществляются при участии коры головного мозга, экстрапирамидной системы и сегментарного аппарата спинного мозга. Произвольные движения связаны с пирамидной системой, которая является отделом нервной системы. Центральный мотонейрон двигательного проводящего пути расположен в пятом слое коры прецентральной извилины головного мозга и представлен гигантскими клетками Беца. В нижней ее части расположены нейроны, которые иннервируют мускулатуру глотки и гортани. В средней части – нейроны, иннервирующие верхние конечности, в верхней – нейроны, иннервирующие нижние конечности. Нейроны этой части коры контролируют произвольные движения конечностей противоположной половины тела. Это связано с перекрестом нервных волокон в нижнем отделе продолговатого мозга. Различают два пути нервных волокон: корково-ядерный, который оканчивается на ядрах продолговатого мозга, и корково-спинномозговой.
В нижней ее части расположены нейроны, которые иннервируют мускулатуру глотки и гортани. В средней части – нейроны, иннервирующие верхние конечности, в верхней – нейроны, иннервирующие нижние конечности. Нейроны этой части коры контролируют произвольные движения конечностей противоположной половины тела. Это связано с перекрестом нервных волокон в нижнем отделе продолговатого мозга. Различают два пути нервных волокон: корково-ядерный, который оканчивается на ядрах продолговатого мозга, и корково-спинномозговой.
Волокна пирамидной системы вызывают преимущественно возбуждение мотонейронов мышц-сгибателей, особенно влияя на отдельные мышцы и даже части мышц верхних конечностей, в частности на мышцы пальцев рук.
Инкубационный период при японском энцефалите составляет от 4 до 14 дней. Начало болезни острое, без четких продромальных явлений, с подъемом температуры до 39–4 °C. На этом уровне она остается в течение 6-10 дней.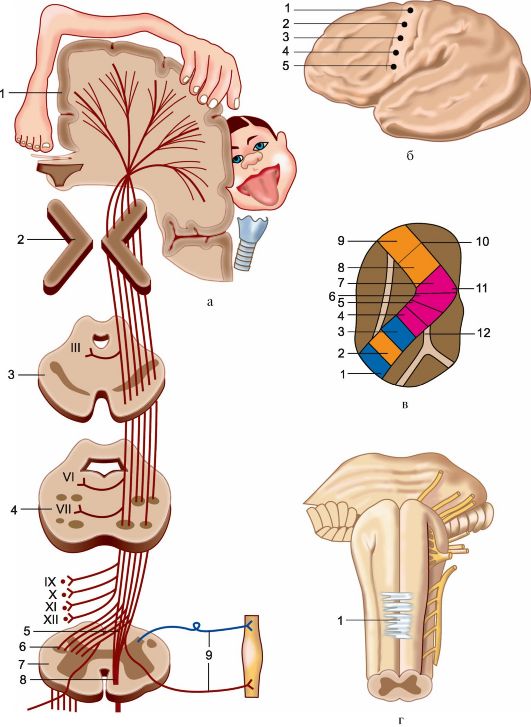 Отмечаются озноб, головная боль, особенно в области лба, боли в пояснице, животе, конечностях, тошнота, рвота. Отмечаются ригидность затылочных мышц, повышение мышечного тонуса, изменения функции глазодвигательных нервов, спутанность или потеря сознания. Сухожильные рефлексы в начале заболевания повышены. Наблюдается симптом Бабинского. Патогенетически начальный период болезни соответствует массовой диссеминации вируса через поврежденные сосудистые стенки в паренхиму мозга, что приводит к отеку мозговой ткани. В периоде разгара возникают симптомы очагового поражения нервной системы, усиливается расстройство сознания вплоть до коматозного. Иногда бывают бред, галлюцинации, психомоторное возбуждение. Мышечная гипертония распространяется на жевательные и затылочные мышцы. При глубоком поражении пирамидной системы могут возникать спастические гемипарезы и параличи. Наблюдаются гиперкинезы мышц лица верхних конечностей; при тяжелом течении болезни могут развиваться клонические или тонические судороги.
Отмечаются озноб, головная боль, особенно в области лба, боли в пояснице, животе, конечностях, тошнота, рвота. Отмечаются ригидность затылочных мышц, повышение мышечного тонуса, изменения функции глазодвигательных нервов, спутанность или потеря сознания. Сухожильные рефлексы в начале заболевания повышены. Наблюдается симптом Бабинского. Патогенетически начальный период болезни соответствует массовой диссеминации вируса через поврежденные сосудистые стенки в паренхиму мозга, что приводит к отеку мозговой ткани. В периоде разгара возникают симптомы очагового поражения нервной системы, усиливается расстройство сознания вплоть до коматозного. Иногда бывают бред, галлюцинации, психомоторное возбуждение. Мышечная гипертония распространяется на жевательные и затылочные мышцы. При глубоком поражении пирамидной системы могут возникать спастические гемипарезы и параличи. Наблюдаются гиперкинезы мышц лица верхних конечностей; при тяжелом течении болезни могут развиваться клонические или тонические судороги. Изменения в двигательной сфере, возникающие в остром периоде, нестойкие, они исчезают по мере улучшения состояния больного. В остром периоде бывает общая болезненность мышц и нервных стволов в конечностях. Лицо, склеры и верхний отдел грудной области гиперемированы. Отмечается повышенная потливость.
Изменения в двигательной сфере, возникающие в остром периоде, нестойкие, они исчезают по мере улучшения состояния больного. В остром периоде бывает общая болезненность мышц и нервных стволов в конечностях. Лицо, склеры и верхний отдел грудной области гиперемированы. Отмечается повышенная потливость.
В патологических случаях при поражении центральной нервной системы повышается рефлекторный фон, который в нормальном состоянии не отмечается у взрослого человека, хотя до созревания пирамидной системы (миелинизации) эти рефлексы можно наблюдать у новорожденных и детей до одного года. Эти рефлексогенные феномены наблюдаются у детей до периода вертикализации и ходьбы.
Связанные понятия (продолжение)
Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
Альтерни́рующие синдромы (лат. alterno — чередоваться; синонимы: альтернирующие параличи, перекрестные параличи) — синдромы, которые сочетают в себе поражение черепных нервов на стороне очага с проводниковыми расстройствами двигательной и чувствительной функций на противоположной стороне.
Мозгово́й ствол, или ствол головного мозга, — традиционно выделяющийся отдел третьего мозга, представляющий собой протяжённое образование, продолжающее спинной мозг.
Глазосердечный рефлекс (синонимы: окулокардиальный рефлекс, рефлекс Ашнера, феномен Ашнера, рефлекс Ашнера — Даньини) — уменьшение пульса на 4-8 сердечных сокращений в минуту при надавливании на глазные яблоки. Рефлекс обусловлен связями тройничного и блуждающего нервов парасимпатической нервной системы. Афферентные пути идут по глазничной ветви тройничного нерва, хотя обнаружены пути и по верхне- и нижнечелюстной ветвям. Эти афферентные пути образуют синаптические связи с висцеральным двигательным…
Сомати́ческая не́рвная систе́ма (от греч. soma — тело) — часть нервной системы человека, представляющая собой совокупность афферентных (чувствительных) и эфферентных (двигательных) нервных волокон, иннервирующих мышцы (у позвоночных — скелетные), кожу, суставы. Соматическая система — это часть периферической нервной системы, которая занимается доставкой моторной (двигательной) и сенсорной (чувственной) информации до центральной нервной системы и обратно. Эта система состоит из нервов, прикрепленных…
Подъязычный нерв (лат. nervus hypoglossus) — XII пара черепных нервов. Ядро подъязычного нерва двигательное, расположено в продолговатом мозге. Отходящие от него двигательные волокна идут к мышцам языка и обеспечивают их движение.
Цитоархитектоническое поле Бродмана 6 — область коры больших полушарий головного мозга, которая располагается в лобной доле в передних отделах прецентральной и задних отделах верхней и средней лобных извилин. При поражении у человека возникает кинетическая (син. эфферентная, премоторная) апраксия.
Тройничный нерв (лат. nervus trigeminus) — V пара черепных нервов, самый крупный из 12 черепных нервов. Нерв смешанного типа, содержит чувствительные и двигательные ядра и волокна. Своё название получил вследствие того, что чувствительная часть на выходе из тройничного узла разделяется на три основные ветви: верхнюю — глазничный нерв (лат. nervus ophthalmicus), среднюю — верхнечелюстной нерв (лат. nervus maxillaris) и нижнюю — нижнечелюстной нерв (лат. nervus mandibularis). Эти три ветви обеспечивают…
Центра́льная не́рвная систе́ма (ЦНС) — основная часть нервной системы животных и человека, состоящая из нейронов, их отростков и вспомогательной глии; у беспозвоночных представлена системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных (включая человека) — спинным и головным мозгом.
Красное ядро (лат. Nucleus ruber) — структура в среднем мозге, участвующая в координации движений. Оно состоит из хвостовой магноцеллюлярной (крупноклеточной) и ростральной — парвоцеллюлярной (мелкоклеточной) части. Красное ядро находится в покрышке среднего мозга рядом с чёрной субстанцией. Красное ядро и чёрная субстанция являются подкорковыми центрами экстрапирамидной двигательной системы.
Лицевой нерв (лат. nervus facialis), седьмой (VII) из двенадцати черепных нервов, выходит из мозга между варолиевым мостом и продолговатым мозгом. Лицевой нерв иннервирует мимические мышцы лица. Также в составе лицевого нерва проходит промежуточный нерв ответственный за иннервацию слёзной железы, стременной мышцы и вкусовой чувствительности двух передних третей языка.
Нервные связи таламуса — это пучки нервных волокон (белого вещества), представляющие собой пучки аксонов отдельных нейронов, и связывающие таламус с другими областями головного мозга, а также со спинным мозгом и органами чувств.
Экстрапирамидная система — Карта знаний
- Экстрапирамидная система
(лат. extra — вне, снаружи, в стороне + pyramis, греч. πϋραμίς — пирамида) — совокупность структур (образований) головного мозга, участвующих в управлении движениями, поддержании мышечного тонуса и позы, минуя кортикоспинальную (пирамидную) систему. Структура расположена в больших полушариях и стволе головного мозга.
Экстрапирамидные проводящие пути образованы нисходящими проекционными нервными волокнами, neurofibrae projectiones descendentes, по происхождению не относящимися к гигантским пирамидным клеткам (клеткам Беца) коры больших полушарий мозга. Эти нервные волокна обеспечивают связи нейронов подкорковых структур (мозжечок, базальные ядра, ствол мозга) головного мозга со всеми отделами нервной системы, расположенными дистальнее.
Экстрапирамидная система состоит из следующих структур головного мозга:
* базальные ганглии,
* красное ядро,
* интерстициальное ядро,
* тектум,
* чёрная субстанция,
* ретикулярная формация моста и продолговатого мозга,
* ядра вестибулярного комплекса,
* мозжечок
премоторная область коры,Экстрапирамидная система — эволюционно более древняя система моторного контроля по сравнению с пирамидной системой. Имеет особое значение в построении и контроле движений, не требующих активации внимания. Является функционально более простым регулятором по сравнению с регуляторами пирамидной системы.
Экстрапирамидная система осуществляет непроизвольную регуляцию и координацию движений, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций (смех, плач). Обеспечивает плавность движений, устанавливает исходную позу для их выполнения.
При поражении экстрапирамидной системы нарушаются двигательные функции (например, могут возникнуть гиперкинезы, паркинсонизм), снижается мышечный тонус.
Экстрапирамидная система (systema extrapyramidale) объединяет двигательные центры коры головного мозга, его ядра и проводящие пути, которые не проходят через пирамиды продолговатого мозга; осуществляет регуляцию непроизвольных компонентов моторики (мышечного тонуса, координации движений, позы).
От пирамидной системы экстрапирамидная система отличается локализацией ядер в подкорковой области полушарий и стволе головного мозга и многозвенностью проводящих путей. Первичными центрами системы являются хвостатое и чечевицеобразное ядра полосатого тела, субталамическое ядро, красное ядро и чёрное вещество среднего мозга. Кроме того, в экстрапирамидную систему входят в качестве интеграционных центры коры большого мозга, ядра таламуса, мозжечок, преддверные и оливные ядра, ретикулярная формация. Частью экстрапирамидной системы является стриопаллидарная система, которая объединяет ядра полосатого тела и их афферентные и эфферентные пути. В стриопаллидарной системе выделяют филогенетически новую часть — стриатум, к которой относятся хвостатое ядро и скорлупа чечевицеобразного ядра, и филогенетически старую часть — паллидум (бледный шар). Стриатум и паллидум различаются по своей нейроархитектонике, связям и функциям.
Стриатум получает волокна из коры большого мозга, центрального ядра таламуса и чёрного вещества. Эфферентные волокна из стриатума направляются в паллидум, а также в чёрное вещество. Из паллидума волокна идут в таламус, гипоталамус, к субталамическому ядру и в ствол головного мозга. Последние образуют чечевицеобразную петлю и частично оканчиваются в ретикулярной формации, частично идут к красному ядру преддверным и оливным ядрам. Следующее звено экстрапирамидных путей составляют ретикулярно-спинномозговой, красноядерно-спинномозговой, преддверно-спинномозговой и оливоспинномозговой пути, оканчивающиеся в передних столбах и промежуточном сером веществе спинного мозга. Мозжечок включается в экстрапирамидную систему посредством путей, соединяющих его с таламусом, красным ядром и оливными ядрами.
Функционально экстрапирамидная система неотделима от пирамидной системы. Она обеспечивает упорядоченный ход произвольных движений, регулируемых пирамидной системой; регулирует врожденные и приобретённые автоматические двигательные акты, обеспечивает установку мышечного тонуса и поддержание равновесия тела; регулирует сопутствующие движения (например движения рук при ходьбе) и выразительные движения (мимика).
Источник: Википедия
Связанные понятия
Тала́мус, иногда — зри́тельные бугры (лат. Thalamus; от др.-греч. θάλαμος — «камера, комната, отсек») — отдел головного мозга, представляющий собой большую массу серого вещества, расположенную в верхней части таламической области промежуточного мозга хордовых животных, в том числе и человека. Впервые описан древнеримским врачом и анатомом Галеном. Таламус — это парная структура, состоящая из двух половинок, симметричных относительно межполушарной плоскости. Таламус находится глубже структур большого…
Ядра таламуса — это группы нейронов таламуса, выделяемые на основании особенностей их анатомо-гистологической структуры и цитоархитектоники, а также паттерна связей с другими частями головного мозга.Все ядра таламуса можно подразделить на шесть групп: передние, медиальные, боковые (латеральные), ретикулярные, внутриламинарные ядра и ядра средней линии. Тонкий Y-образный слой миелинизированных нервных волокон, так называемая внутренняя медуллярная пластинка, разграничивает между собой переднюю, медиальную…
Головно́й мозг (лат. cerebrum, др.-греч. ἐγκέφαλος) — главный орган центральной нервной системы подавляющего большинства хордовых, её головной конец; у позвоночных находится внутри черепа. В анатомической номенклатуре позвоночных, в том числе человека, мозг в целом чаще всего обозначается как encephalon — латинизированная форма греческого слова; изначально латинское cerebrum стало синонимом большого мозга (telencephalon).
Функциональные различия ядер таламуса — это различия, проводимые между ядрами таламуса не на основании их анатомо-гистологических характеристик или особенностей их цитоархитектоники, а на основании характера выполняемых ими физиологических функций, а также на основании типа их связей с другими частями мозга и типа передаваемой по этим связям и обрабатываемой этими ядрами информации. С этой точки зрения, среди ядер таламуса функционально выделяют первичные сенсорные и моторные ретрансляционные ядра…
Вегетати́вная не́рвная систе́ма (от лат. vegetatio — возбуждение, от лат. vegetativus — растительный), ВНС, автономная нервная система, ганглионарная нервная система (от лат. ganglion — нервный узел), висцеральная нервная система (от лат. viscera — внутренности), органная нервная система, чревная нервная система, systema nervosum autonomicum (PNA) — часть нервной системы организма, комплекс центральных и периферических клеточных структур, регулирующих функциональный уровень организма, необходимый…
Гипотала́мус (лат. hypothalamus, от греч. ὑπό — «под» и θάλαμος — «комната, камера, отсек, таламус») — небольшая область в промежуточном мозге, включающая в себя большое число групп клеток (свыше 30 ядер), которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма. Гипоталамус связан нервными путями практически со всеми отделами центральной нервной системы, включая кору, гиппокамп, миндалину, мозжечок, ствол мозга и спинной мозг. Вместе с гипофизом гипоталамус образует гипоталамо-гипофизарную…
Мозгово́й ствол, или ствол головного мозга, — традиционно выделяющийся отдел третьего мозга, представляющий собой протяжённое образование, продолжающее спинной мозг.
Ядро одиночного пути (лат. nucleus tractus solitarii) — одно из ядер продолговатого мозга у человека и млекопитающих. Представляет собой тяж серого вещества, который тянется вдоль одиночного пути. Отростки нейронов ядра входят в состав лицевого, языко-глоточного и блуждающего нервов. Ядро является местом входа чувствительных нервов от внутренних органов, служит переключателем вагусных рефлексов. Участвует в автономной регуляции сердечно-сосудистой, иммунной, пищеварительной и дыхательной систем…
Аллоталамус — это особенные, атипичные ядра таламуса, отличающиеся по своему анатомо-гистологическому строению и по содержанию нейромедиаторов от всех остальных ядер таламуса, имеющих типичное строение. Эти типичные ядра таламуса, в противоположность аллоталамусу, совокупно называют изоталамусом.
Пирамидные знаки — группа патологических рефлексов, которые возникают вследствие поражения центрального нейрона коры головного мозга, либо кортикоспинальных или кортиконуклеарных проводящих путей, идущих соответственно к мотонейронам спинного мозга или двигательным ядрам черепных нервов.
Нейроанатомия — это область биологических наук, изучающая анатомическое строение (структурная нейроанатомия) и функциональную организацию (функциональная нейроанатомия) нервных систем различных животных, обладающих ею. В отличие от животных, обладающих радиальной симметрией (например, медуз), у которых нервная система представляет собой диффузную нервную сеть, животные, обладающие билатеральной симметрией, имеют отдельные, чётко анатомически и гистологически отграниченные от других тканей, нервные…
Цитоархитектоническое поле Бродмана 6 — область коры больших полушарий головного мозга, которая располагается в лобной доле в передних отделах прецентральной и задних отделах верхней и средней лобных извилин. При поражении у человека возникает кинетическая (син. эфферентная, премоторная) апраксия.
Нервные связи таламуса — это пучки нервных волокон (белого вещества), представляющие собой пучки аксонов отдельных нейронов, и связывающие таламус с другими областями головного мозга, а также со спинным мозгом и органами чувств.
Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
Ганглий (др.-греч. γάγγλιον — узел), или нервный узел — скопление нервных клеток, состоящее из тел, дендритов и аксонов нервных клеток и глиальных клеток. Обычно ганглий имеет также оболочку из соединительной ткани. Имеются у многих беспозвоночных и всех позвоночных животных. Часто соединяются между собой, образуя различные структуры (нервные сплетения, нервные цепочки и т. п.).
Первичная моторная кора, или первичная двигательная кора (англ. Primary motor cortex) — область моторной коры, расположенная в средне-задней части прецентральной извилины, примыкающей к центральной борозде. Соответствует цитоархитектоническому полю Бродмана 4 и каудальной части поля 6. Содержит клетки Беца и другие мотонейроны, аксоны которых достигают сегментов спинного мозга. Одной из основных функций первичной моторной коры является контроль произвольных дискретных движений, совершаемых группами…
Неоталамус (лат. neothalamus) — эволюционно (филогенетически) более молодая часть таламуса, включающая в себя, в частности, латеральную группу ядер, ядра подушки, а также метаталамус, состоящий, в свою очередь, из медиального и латерального коленчатых тел. Она противопоставляется эволюционно более древнему палеоталамусу.
Красное ядро (лат. Nucleus ruber) — структура в среднем мозге, участвующая в координации движений. Оно состоит из хвостовой магноцеллюлярной (крупноклеточной) и ростральной — парвоцеллюлярной (мелкоклеточной) части. Красное ядро находится в покрышке среднего мозга рядом с чёрной субстанцией. Красное ядро и чёрная субстанция являются подкорковыми центрами экстрапирамидной двигательной системы.
Ретикулярная формация (лат. reticulum — сеточка, formatio — образование) — это образование, тянущееся вдоль всей оси ствола головного мозга. Своим названием оно обязано сетчатой структуре, образуемой его нервными клетками с очень сложными связями. Формация состоит из ретикулярных ядер и большой сети нейронов с разветвлёнными аксонами и дендритами, представляющих единый комплекс, который осуществляет активацию коры головного мозга и контролирует рефлекторную деятельность спинного мозга. Эта сеть нейронов…
Вставочный нейрон (синонимы: интернейрон, промежуточный нейрон; англ. interneuron, relay neuron, association neuron, bipolar neuron) — нейрон, связанный только с другими нейронами, в отличие от двигательных нейронов, иннервирующих мышечные волокна, и сенсорных нейронов, преобразующих стимулы из внешней среды в электрические сигналы.
Нервная система дрозофилы фруктовой (Drosophila melanogaster) является двусторонне-симметричной и состоит из мозга с двумя полушариями и брюшной нервной цепи, которая представляет собой сросшиеся в цепочку подглоточный ганглий (suboesophageal ganglion), три грудных ганглия (thoracic ganglia) и несколько брюшных ганглиев. Ганглии в брюшной нервной цепи содержат моторные нейроны и интернейроны, которые необходимы для управления тем сегментом тела, который они иннервируют. Мозг в свою очередь отвечает…
Фильтрация сенсорной информации — фильтрация афферентных сигналов нервной системой. В результате такой фильтрации на определённые уровни обработки поступает только часть полученной предшествующими уровнями сенсорной информации.
Передние ядра таламуса, или передняя группа ядер таламуса, — это совокупность таламических ядер, находящихся в ростральной части дорсального таламуса. К передней группе ядер таламуса причисляют антеромедиальное ядро, антеродорсальное ядро, антеровентральное ядро. К передней группе ядер таламуса нередко относят также латеральное дорсальное ядро (оно же поверхностное ядро) таламуса.
Нервная система человека часто делится на центральную нервную систему (ЦНС) и периферическую нервную систему (ПНС). ЦНС состоит из головного и спинного мозга. ПНС состоит из всех других нервов и нейронов, которые не лежат в пределах ЦНС. Преобладающее большинство нервов (которые фактически являются аксонами нейронов) принадлежит ПНС. Периферийная нервная система делится на соматическую нервную систему и вегетативную нервную систему.
Пирамидная система, пирамидный путь (лат. tractus pyramidalis, PNA) — система нервных структур. Поддерживает сложную и тонкую координацию движений.
Прилежащее ядро (лат. nucleus accumbens, сокращенно — NAcc, другие названия: прилегающее ядро перегородки, полулежащее ядро) — группа нейронов в вентральной части полосатого тела, является важной частью мезолимбического пути, участвующего в системе вознаграждений, формировании удовольствия, смеха, зависимости, агрессии, страха и эффекта плацебо.
Центра́льная не́рвная систе́ма (ЦНС) — основная часть нервной системы животных и человека, состоящая из нейронов, их отростков и вспомогательной глии; у беспозвоночных представлена системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных (включая человека) — спинным и головным мозгом.
Субталамус, иначе называемый преталамус или периталамус, вентральный таламус — это часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо субталамуса, входят также таламус, эпиталамус и метаталамус, но не входят гипоталамус и гипофиз, относимые к гипоталамической области. Его наиболее выраженной структурой является так называемое субталамическое ядро. Субталамус соединён нервными связями с бледным шаром, базальным ядром в конечном мозге…
Нервная ткань — ткань эктодермального происхождения, представляет собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию.
Ноцице́пция; ноциперце́пция; физиологи́ческая боль — это активность в афферентных (чувствительных) нервных волокнах периферической и центральной нервной системы, возбуждаемая разнообразными стимулами, обладающими пульсирующей интенсивностью. Данная активность генерируется ноцицепторами, или по-другому рецепторами боли, которые могут отслеживать механические, тепловые или химические воздействия, превышающие генетически установленный порог возбудимости. Получив повреждающий стимул, ноцицептор передаёт…
Симпати́ческая не́рвная систе́ма (от греч. συμπαθής чувствительный, сочувственный) — часть автономной (вегетативной) нервной системы, ганглии которой расположены на значительном расстоянии от иннервируемых органов. Активация вызывает возбуждение сердечной деятельности.
Рефлекторная дуга (нервная дуга) — путь, проходимый нервными импульсами при осуществлении рефлекса.
Лимбическая система (от лат. limbus — граница, край) — совокупность ряда структур головного мозга, расположенных на обеих сторонах таламуса, непосредственно под конечным мозгом. Окутывает верхнюю часть ствола головного мозга, будто поясом, и образует его край (лимб). Это не отдельная система, но скопление структур из конечного мозга, промежуточного мозга (диэнцефалона), и среднего мозга (мезэнцефалона).
Наруше́ние созна́ния — состояние расстроенного сознания, синдромы его выключения (кома, сопор, оглушение) или помрачения (онейроид, делирий, сумеречное состояние сознания). Полноценное функционирование сознания предполагает состояние бодрствования, обусловленное полноценной реализацией когнитивной функции полушарий головного мозга и их взаимоотношений с пробуждающими механизмами ретикулярной формации, распространение ядер и проводящих путей которой обнаружены в диэнцефальной области, среднем мозге…
Общая структурно-функциональная модель мозга — концепция мозга как материального субстрата психики, разработанная А. Р. Лурией на основе изучения нарушений психической деятельности при различных локальных поражениях центральной нервной системы. Согласно данной модели, мозг может быть разделён на три основных блока, которые имеют собственное строение и роль в психическом функционировании…
Эволюция таламуса — это процесс последовательного усложнения в ходе филогенеза хордовых структуры и функций таламуса. Эволюция таламуса у хордовых является одним из частных, но достаточно важных, проявлений общего процесса эволюции головного мозга, и, в целом, нервной системы у этих видов.
Периферическая нервная система — условно выделяемая часть нервной системы, находящаяся за пределами головного и спинного мозга. Она состоит из черепных и спинальных нервов, а также нервов и сплетений вегетативной нервной системы, соединяя центральную нервную систему с органами тела.
Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
Головно́й мозг челове́ка (лат. encephalon) является органом центральной нервной системы, состоящей из множества взаимосвязанных между собой нервных клеток и их отростков.
Чёрная субстанция, также чёрное вещество (лат. Substantia nigra) — составная часть экстрапирамидной системы, находящаяся в области четверохолмия среднего мозга. Играет важную роль в регуляции моторной функции, тонуса мышц, осуществлении статокинетической функции участием во многих вегетативных функциях: дыхании, сердечной деятельности, тонусе кровеносных сосудов. Впервые обнаружена французским анатомом и врачом Феликсом Вик-д’Азиром в 1784 году.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться один с другим, формируя биологические…
Прямая извилина (лат. gyrus rectus; англ. straight gyrus, StG) — извилина в нижней части лобной доли, расположенная между медиальной обонятельной бороздой и медиальным краем полушария. Соответствует медиальной части цитоархитектонического поля Бродмана 11.
Зрительная кора является частью коры больших полушарий головного мозга, отвечающей за обработку визуальной информации. В основном, она сосредоточена в затылочной доле каждого из полушарий головного мозга..
Моторная кора, также известная как двигательная кора (англ. Motor cortex) — области коры больших полушарий, отвечающих за планирование, контроль и выполнение произвольных движений.
Подушка таламуса (лат. pulvinar thalami, или просто лат. pulvinar) — это группа ядер (телец нейронов), расположенных в таламусе (части промежуточного мозга позвоночных) животных. Эта группа ядер совокупно называется пульвинаром, или подушкой таламуса (лат. pulvinar thalami), а как коллекция отдельных ядер — пульвинарными ядрами, или ядрами подушки таламуса (лат. nuclei pulvinares thalami).
Цитоархитектоническое поле Бродмана 7 — область коры больших полушарий головного мозга, которая располагается в верхних отделах теменной доли, позади постцентральной извилины и цитоархитектонического поля Бродмана 5 и спереди коры затылочной доли.
Олигодендроциты, или олигодендроглия — это вид нейроглии, открытый Пио дель Рио-Ортегой (1928 год). Олигодендроциты есть только в центральной нервной системе, которая у позвоночных включает в себя головной мозг и спинной мозг.
Покрышка мозга, или тегментум (лат. tegmentum) — это общее название совокупности нескольких областей ствола мозга. Покрышка мозга находится между желудочковой системой мозга и базальными и вентральными структурами на каждом из уровней, через которые «покрышка мозга» проходит. На уровне среднего мозга она расположена дорсально по отношению к ножке мозга и отделена полулунной областью чёрного вещества от основания ножки. Таким образом, на уровне среднего мозга покрышка мозга образует его нижнюю поверхность…
Преддверно-улитковый нерв (лат. nervus vestibulocochlearis) — (VIII пара черепных нервов) нерв специальной чувствительности, отвечающий за передачу слуховых импульсов, а также импульсов, исходящих из вестибулярного отдела внутреннего уха.
Пирамидная система — Википедия
Материал из Википедии — свободной энциклопедии
Пирамидная система — красный цвет.
Поперечный разрез спинного мозга. Пирамидная система — красный цвет.
Пирамидная система, пирамидный путь (лат. tractus pyramidalis, PNA) — система нервных структур. Поддерживает сложную и тонкую координацию движений[1][2].
Пирамидная система — одно из поздних приобретений эволюции. Низшие позвоночные пирамидальной системы не имеют, она появляется только у млекопитающих, и достигает наибольшего развития у обезьян и особенно у человека[1]. Пирамидная система играет особую роль в прямохождении[2].
Пирамидный путь
Корково-спинномозговой (пирамидный) путь (лат. tractus corticospinalis)
Кора полушарий головного мозга в V слое содержит клетки Беца (или гигантские пирамидные клетки)[3].
В 1874 году учёный Владимир Алексеевич Бец обнаружил и описал гигантские пирамидальные клетки коры головного мозга (клетки Беца).
Пирамидный путь осуществляется нервными волокнами, которые исходят от клеток Беца и спускаются в спинной мозг, не прерываясь. Пирамидный путь проходит через внутреннюю капсулу, ствол мозга, отдавая на своем пути ответвления (коллатерали) с экстрапирамидной системой, а также с подкорковыми ядрами (двигательными ядрами черепно-мозговых нервов)[2][3].
Волокна перекрещиваются на границе головного и спинного мозга (большая часть — в продолговатом мозге, меньшая — в спинном). Далее они проходят через спинной мозг (передние и боковые столбы спинного мозга). В каждом сегменте спинного мозга эти волокна образуют синаптические окончания (см. Синапс), которые отвечают за определенный участок тела (шейный отдел спинного мозга — за иннервацию рук, грудной — за туловище, а поясничный отдел — за ноги)[3]. Импульсы от коры головного мозга эти волокна передают либо непосредственно, либо через вставочные нейроны[1].
Непосредственное раздражение определенных участков коры головного мозга приводит к судорогам мышц, соответствующих участку коры — проекционной двигательной зоне. При раздражении верхней трети передней центральной извилины возникает судорога мышц ноги, средней — руки, нижней — лица, причем, на стороне, противоположной очагу раздражения в полушарии. Эти судороги носят название парциальных (джексоновских). Их открыл английский невролог Д. Х. Джексон (1835—1911). В проекционной двигательной зоне каждого полушария головного мозга представлены все мышцы противоположной половины тела[2].
Типы нервных волокон
Пирамидная система человека содержит около 1 млн нервных волокон. Различают следующие типы волокон[1]:
| Тип нервных волокон | Диаметр | Скорость проведения | Функция |
|---|---|---|---|
| Толстые, быстропроводящие | 16 мкм | до 80 м/с | обеспечивают быстрые фазные движения |
| Тонкие, медленнопроводящие | 4 мкм | от 25 до 7 м/с | отвечают за тоническое состояние мышц |
Наибольшее количество пирамидных клеток (клеток Беца) иннервирует мелкие мышцы, отвечающие за тонкие дифференцированные движения кисти, мимику и речевой акт. Значительно меньшее их количество иннервирует мышцы туловища и нижних конечностей[2].
Патология
Повреждения пирамидной системы проявляются параличами, парезами, патологическими рефлексами[1].
Поражение пирамидной системы может быть вызвано воспалением (см. Энцефалит), нарушением мозгового кровообращения (см. Инсульт), опухолью, черепно-мозговой травмой и другими причинами[2].
В зависимости от локализации патологического процесса различают следующие проявления[2].
Центральный (спастический) паралич, или парез
Также называется — пирамидная недостаточность, спастическая атаксия, болезнь Пьера Мари. Возникает при поражении проекционной зоны коры головного мозга. Если поражены клетки Беца в коре головного мозга (или их аксоны), то возникает спастический (от слова спазм, то есть когда тонус мышц повышен) паралич. При этом клетка Беца начинает посылать избыточное количество нервных импульсов к мышцам. Это приводит к повышению мышечного тонуса и рефлексов, и возникает дрожание. Это состояние называют центральным параличом (при неполной утрате произвольных движений — центральным парезом). При центральном параличе(парезе) нарушения питания конечности(гипотрофии, атрофии) не происходит[3].
Периферический (вялый) паралич, или парез
Если поражены периферические нервы или сплетения, то возникает вялый, или периферический паралич (а при неполной утрате функции — парез) — снижение мышечного тонуса, вплоть до полной парализации мышцы[3].
Методы диагностики пирамидной недостаточности
- Магнитно-резонансная томография (МРТ) — обязательный метод обследования при эпилепсии и судорогах.
- Компьютерная томография головного мозга (по рекомендации Международной лиги борьбы против эпилепсии, КТ производится в качестве дополнительного метода обследования, или когда невозможно сделать МРТ).
- Электромиография — это метод исследования нервно-мышечной системы посредством регистрации электрических потенциалов мышц.
- Электроэнцефалография (ЭЭГ исследование) — позволяет выявить судороги. Более 65 % судорог происходит во сне, поэтому необходима запись ээг во время физиологического, естественного сна. Из-за непостоянного характера судорог проводят длительный мониторинг (видео или холтеровский). Исследование выявляет появления диффузных дельта волн, также синхронизацию волн тета-диапа
Пирамидальные тракты — Pyramidal tracts
В пирамидных трактах включают как corticobulbar тракта и кортикоспинальный тракт . Это скопления эфферентных нервных волокон от верхних мотонейронов, которые идут из коры головного мозга и оканчиваются либо в стволе мозга ( кортикобульбарный ), либо в спинном мозге ( кортикоспинальный ) и участвуют в контроле моторных функций тела.
Кортикобульбарный тракт проводит импульсы от мозга к черепным нервам . Эти нервы контролируют мышцы лица и шеи и участвуют в мимике, жевании, глотании и других двигательных функциях.
Кортикоспинальный тракт проводит импульсы от головного мозга к спинному мозгу. Он состоит из переднего и бокового трактов . Кортикоспинальный тракт участвует в произвольных движениях. Большинство волокон кортикоспинального тракта пересекаются в продолговатом мозге , в результате чего мышцы контролируются противоположной стороной мозга. Кортикоспинальный тракт содержит аксоны пирамидных клеток, крупнейшими из которых являются клетки Беца , расположенные в коре головного мозга.
Пирамидальные тракты названы потому, что они проходят через пирамиды продолговатого мозга. Кортикоспинальные волокна сходятся к точке при спуске от внутренней капсулы к стволу мозга в нескольких направлениях, создавая впечатление перевернутой пирамиды. Вовлечение пирамидного тракта на любом уровне приводит к пирамидным признакам .
Миелинизации из пирамидальных волокон является неполным при рождении и постепенно прогрессирует в направлении черепно-хвостового и , таким образом , постепенно набирает функциональность . Большая часть миелинизации завершается к двухлетнему возрасту, а после этого она очень медленно прогрессирует в кранио-каудальном направлении до двенадцати лет.
Структура
Пирамидальные тракты
Термин « пирамидные пути» относится к верхним мотонейронам, которые берут начало в коре головного мозга и оканчиваются в спинном мозге ( кортикоспинальный ) или стволе мозга ( кортикобульбарный ). Нервы возникают в коре головного мозга , проходят вниз и могут пересекать стороны в продолговатом мозге и путешествовать как часть спинного мозга, пока не синапсируются с интернейронами в сером столбце спинного мозга.
Есть некоторые вариации в терминологии. В пирамидных трактах окончательно охватывают корковы трактаты , и многие авторы включают corticobulbar тракты .
Кортикоспинальный тракт
Нервные волокна в кортикоспинальном тракте происходят из пирамидных клеток в слое V коры головного мозга . Волокна возникают из первичной моторной коры (около 30%), дополнительной моторной области и премоторной коры (вместе также около 30%), а соматосенсорная кора , теменная доля и поясная извилина снабжают остальное. Клетки имеют свои тела в коре головного мозга, а аксоны образуют основную часть пирамидных трактов. Аксоны нервов перемещаются из коры через заднюю часть внутренней капсулы , через ножку головного мозга в ствол мозга и передний продолговатый мозг . Здесь они образуют два выступа, называемые продолговатыми пирамидами продолговатого мозга . Ниже выступов большинство аксонов переходят на противоположную сторону, с которой они произошли, это называется перекрестом . Пересекающиеся аксоны перемещаются к внешней части продолговатого мозга и формируют боковой кортикоспинальный тракт , тогда как оставшиеся волокна образуют передний кортикоспинальный тракт . Около 80% аксонов пересекаются и образуют боковой кортикоспинальный тракт; 10% волокон не пересекаются и не присоединяются к тракту, а 10% волокон проходят по переднему кортикоспинальному тракту.
Нервные аксоны , путешествующие вниз тракт являются эфферентными нервными волокнами этих верхних двигательных нейронов . Эти аксоны перемещаются по трактам в белом веществе спинного мозга, пока не достигнут позвоночного уровня мышцы, которую они будут иннервировать. В этот момент аксоны синапсируются с нижними мотонейронами . Большинство аксонов непосредственно не синапс с нижними двигательными нейронами, а синапс с интернейроном, который затем синапс с нижним двигательным нейроном. Обычно это происходит в переднем сером столбце . Аксоны нервов бокового кортикоспинального тракта, которые не пересекаются в продолговатом мозге, делают это на уровне спинного мозга, в котором они заканчиваются.
Эти тракты содержат более 1 миллиона аксонов, и большинство аксонов миелинизированы. Кортикоспинальные тракты миелинизируются в основном в течение первого и второго года после рождения. Большинство нервных аксонов имеют небольшой диаметр (<4 мкм). Около 3% нервных аксонов имеют гораздо больший диаметр (16 мкм) и возникают из клеток Беца , в основном в области ног первичной моторной коры. Эти клетки примечательны своей высокой скоростью проводимости, более 70 м / с, самой быстрой передачей любых сигналов от головного мозга к спинному мозгу.
Горизонтальный разрез нижней части моста, показывающий волокна кортикоспинального тракта (№ 19), проходящие через ядра моста.
Кортикобульбарный тракт
Волокна из вентральной моторной коры проходят по кортикоспинальному тракту через внутреннюю капсулу, но оканчиваются в ряде мест в среднем мозге ( кортико-мезэнцефалический тракт ), мосте ( кортикопонтинный тракт ) и продолговатом мозге ( кортико-бульбарный тракт ). В верхние моторные нейроны из corticobulbar тракта синапсы с интернейронов или непосредственно с нижних двигательных нейронов , расположенных в двигательных черепных нервов ядер , а именно глазодвигательный , блоковый , двигатель ядро тройничного нерва , отводящий , лицевой нерв и вспомогательное оборудование , а в ядре ambiguus к подъязычные , блуждающие и аксессуары нервов . Эти ядра снабжены нервами с обеих сторон мозга, за исключением частей лицевого нерва, которые контролируют мышцы нижней части лица. Эти мышцы иннервируются только нервами с контралатеральной (противоположной) стороны коры.
Функция
Нервы в пределах корково тракта участвует в движении из мышц тела. Из-за перекрещивания волокон мышцы снабжаются энергией со стороны мозга, противоположной мышечной. Нервы кортикобульбарного тракта участвуют в движении мышц головы. Они участвуют в глотании, фонации и движениях языка. В силу вовлечения лицевого нерва кортикобульбарный тракт также отвечает за передачу выражения лица . За исключением нижних мускулов мимики, все функции кортикобульбарного тракта включают сигналы с обеих сторон мозга.
Экстрапирамидная система относится к трактам в спинном мозге , участвующем в непроизвольных движениях , но не часть пирамидных путей. В их функции входит контроль осанки и мышечного тонуса .
Клиническое значение
- Повреждение волокон кортикоспинальных трактов на любом участке их пути от коры головного мозга до нижнего конца спинного мозга может вызвать синдром верхнего двигательного нейрона .
- Через несколько дней после повреждения верхних мотонейронов появляется набор двигательных признаков и симптомов, включая спастичность , гиперактивные рефлексы, потерю способности выполнять тонкие движения и подошвенный разгибатель, известный как признак Бабинского .
- Симптомы обычно возникают наряду с другими сенсорными проблемами.
- Причины могут включать такие расстройства, как инсульт , церебральный паралич , субдуральное кровоизлияние , абсцессы и опухоли , нейродегенеративные заболевания, такие как множественная системная атрофия , воспаление, такое как менингит и рассеянный склероз , а также травмы спинного мозга, в том числе из-за смещения межпозвоночного диска .
- Если кортикобульбарный тракт поврежден только с одной стороны, то будет затронута только нижняя часть лица, однако, если есть поражение как левого, так и правого трактов, то результатом будет псевдобульбарный паралич . Это вызывает проблемы с глотанием, речью и эмоциональную лабильность .
- Сильные непроизвольные движения, приводящие к потере трудоспособности, такие как гемибаллизм или тяжелая хорея, могут утомить пациента и стать опасной для жизни ситуацией.
- В прошлом, это условие лечилось частичным разрезом пирамидного тракта либо в первичной моторной коре или на круз Cerebri (pedunculotomy).
Дополнительные изображения
Рассечение ствола мозга. Боковой вид.
Поверхностное рассечение ствола мозга. Вид снизу.
Моторный тракт.
В популярной культуре
В «Европейских каникулах» от National Lampoon семья Грисволд выигрывает отпуск на игровом шоу под названием « Свинья в мешке», когда их противники не могут правильно ответить на вопрос о пирамидальных трактах.
Ссылки
внешняя ссылка
Нейроанатомия онлайн: Лаборатория 9 — Нисходящие пути к спинному мозгу
Кортикоспинальный (пирамидный) путь
Кортикоспинальный путь — это путь с одним нейроном от коры головного мозга к серой части спинного мозга. Этот путь состоит из всех аксонов, которые: (1) происходят из клеток коры головного мозга, (2) проходят через пирамиды продолговатого мозга и (3) заканчиваются в спинном мозге. Кортикоспинальный путь — одна из наиболее заметных нисходящих волоконных систем в нервной системе человека.Это касается выполнения волевых движений, особенно изолированных движений пальцев и рук. Разрушение этого пути приводит к дефициту произвольных движений, который наиболее выражен в дистальных отделах конечностей. Проксимальные суставы и грубые движения поражаются менее серьезно и не навсегда. Поражения, затрагивающие только кортикоспинальный путь, вызывают дефицит, который обычно не является серьезным и может быть трудно обнаружить через некоторое время.
Большинство клеток, образующих этот путь, расположены в прецентральной извилине (моторная кора), части лобной доли впереди прецентральной извилины (премоторная и дополнительная моторная зона) и части теменной доли (постцентральная извилина). и задние теменные области).Аксоны этих корковых клеток сходятся в лучевой коронке, проходят вниз в задней конечности внутренней капсулы, образуя crus cerebri вместе с кортикобульбарными и кортикопонтинными волокнами. Кортикоспинальные волокна спускаются через мост компактными пучками вместе с другими кортикофугальными волокнами. В ростральном мозговом слое кортикоспинальные волокна образуют компактный пучок, пирамиду, который спускается к каудальному мозгу, где около 90% волокон пересекаются в пирамидальном перекресте. Перекрещенные волокна образуют латеральный кортикоспинальный тракт спинного мозга и расположены в задней части латерального семенного канатика, медиальнее заднего спиноцеребеллярного тракта.Непересекающиеся волокна, образующие передний кортикоспинальный тракт, располагаются медиально в переднем семяпроводе вдоль стенки передней срединной щели.
Боковой кортикоспинальный тракт простирается по всей длине спинного мозга и постепенно уменьшается в размере по мере того, как все больше и больше волокон уходит, чтобы оканчиваться в сером веществе спинного мозга. Ниже L3 , т.е. каудальнее того места, где находится задний спиноцеребеллярный тракт, латеральный кортикоспинальный тракт расположен вдоль заднебокового края спинного мозга.Волокна боковых кортикоспинальных трактов оканчиваются ипсилатеральным канатиком (т. Е. Контралатеральны исходным клеткам).
Меньшая часть пирамидных волокон спускается, не пересекаясь, как передний или прямой кортикоспинальный тракт, в передний семенной канатик. Большинство волокон доходят только до верхнего грудного отдела спинного мозга. Волокна воздействуют на мотонейроны, иннервирующие мышцы верхних конечностей и шеи. Большинство волокон непересеченного переднего кортикоспинального тракта пересекаются в передней белой спайке спинного мозга и оканчиваются серой контрлатеральной стороной.Некоторые остаются не перечеркнутыми. Контралатеральная проекция (поперечный латеральный кортикоспинальный тракт) влияет на нейроны, иннервирующие дистальные (в основном) и проксимальные мышцы конечностей. Ипсилатеральная кортикальная проекция (неперекрещенные передние кортикоспинальные волокна) влияет на двигательные нейроны, иннервирующие более проксимальные мышцы конечностей. Около 55% всех кортикоспинальных волокон оканчиваются в шейном канатике, 20% — в грудном и 25% — в пояснично-крестцовом сегментах. Это говорит о том, что кортикоспинальный контроль над верхней конечностью (руками и пальцами) намного больше, чем над нижней конечностью.
Введение в нейроны и нейронные сети | Раздел 1, Вступительная глава | Neuroscience Online: Электронный учебник для неврологии | Кафедра нейробиологии и анатомии
Три фунта желеобразного материала, обнаруженные в наших черепах, — это самая сложная машина на Земле и, возможно, во Вселенной. Его феноменальные свойства были бы невозможны без сотен миллиардов нейронов, составляющих его, и, что важно, без связей между этими нейронами. К счастью, многое известно о свойствах отдельных нейронов и простых нейронных сетей, и аспекты сложных нейронных сетей только начинают разгадываться.Эта глава начнется с обсуждения нейрона, элементарного узла или элемента мозга, а затем перейдет к обсуждению способов, которыми отдельные нейроны общаются друг с другом. Что делает нервную систему таким фантастическим устройством и отличает мозг от других органов тела, так это не то, что она имеет 100 миллиардов нейронов, а то, что нервные клетки способны взаимодействовать друг с другом таким высоко структурированным образом, что формируют нейронные сети . Чтобы понять нейронные сети, необходимо понять способы, которыми один нейрон общается с другим через синаптические связи, и процесс, называемый синаптической передачей.Синаптическая передача бывает двух основных видов: возбуждение и торможение. Всего несколько взаимосвязанных нейронов (микросхема ) могут выполнять сложные задачи, такие как опосредование рефлексов, обработка сенсорной информации, генерирование движения и опосредование обучения и памяти. Более сложные сети ( макросхем, ) состоят из множества встроенных микросхем. Макросхемы опосредуют высшие функции мозга, такие как распознавание объектов и познание. Итак, в нервной системе повсеместно распространены сети нескольких уровней.Сети также преобладают в пределах нейронов. Эти наноцепей составляют базовый биохимический механизм, обеспечивающий ключевые свойства нейронов, такие как обучение и память, а также генезис нейрональной ритмичности.
Нейрон
Основные морфологические особенности нейронов
100 миллиардов нейронов в головном мозге имеют ряд общих черт (рис. 1). Нейроны отличаются от большинства других клеток тела тем, что они поляризованы и имеют различные морфологические области, каждая из которых выполняет определенные функции. Дендриты — это область, где один нейрон получает связи от других нейронов. Тело клетки или сома содержит ядро и другие органеллы, необходимые для функционирования клетки. Аксон является ключевым компонентом нервных клеток, по которому информация передается от одной части нейрона (например, тела клетки) к концевым областям нейрона. Аксоны могут быть довольно длинными, достигая метра или около того в некоторых сенсорных и двигательных нервных клетках человека.Синапс является конечной областью аксона, и именно здесь один нейрон формирует соединение с другим и передает информацию посредством процесса синаптической передачи . Нейрон цвета морской волны на рисунке 1 (нажмите «Нейрон, связанный с постсинаптическим нейроном») называется постсинаптическим нейроном. Терминал коричневого цвета слева, следовательно, называется пресинаптическим нейроном. Один нейрон может получать контакты от множества разных нейронов.На рис. 1 (щелкните «Нейрон, получающий синаптический вход») показан пример трех пресинаптических нейронов, контактирующих с одним постсинаптическим нейроном коричневого цвета, но было подсчитано, что один нейрон может получать контакты до 10 000 других клеток. Следовательно, потенциальная сложность сетей огромна. Точно так же любой нейрон может контактировать до 10 000 постсинаптических клеток. (Обратите внимание, что нейрон коричневого цвета, который был пресинаптическим по отношению к нейрону цвета морской волны, является постсинаптическим по отношению к розовым, зеленым и синим нейронам.Таким образом, большинство «пресинаптических» нейронов являются «постсинаптическими» по отношению к некоторым другим нейронам.
Рисунок 1 (щелкните «Синапс») также показывает расширенный вид синапса. Обратите внимание, что пресинаптическая клетка не связана напрямую с постсинаптической клеткой. Они разделены промежутком, известным как синаптическая щель . Следовательно, чтобы общаться с постсинаптической клеткой, пресинаптический нейрон должен выпускать химический мессенджер. Этот посланник находится внутри везикул, содержащих нейротрансмиттер, (синие точки обозначают нейромедиатор).Потенциал действия, который вторгается в пресинаптический терминал, заставляет эти везикулы сливаться с внутренней поверхностью пресинаптической мембраны и высвобождать их содержимое посредством процесса, называемого экзоцитоз . Освободившийся передатчик диффундирует через промежуток между пре- и постсинаптической клеткой и очень быстро достигает постсинаптической стороны синапса, где связывается со специализированными рецепторами, которые «распознают» передатчик. Связывание с рецепторами приводит к изменению проницаемости ионных каналов в мембране и, в свою очередь, к изменению мембранного потенциала постсинаптического нейрона, известного как постсинаптический синаптический потенциал (PSP) .Таким образом, передача сигналов между нейронами связана с изменениями электрических свойств нейронов. Чтобы понять нейроны и нейронные цепи, необходимо понимать электрические свойства нервных клеток.
Потенциал покоя и потенциал действия
Потенциалы покоя. На рисунке 2 показан пример идеализированной нервной клетки. Во внеклеточную среду помещен микроэлектрод. Микроэлектрод — это не что иное, как небольшой кусок стеклянной капиллярной трубки, который растягивается под действием тепла, образуя очень тонкий наконечник диаметром порядка 1 микрона.Микроэлектрод заполняется проводящим раствором и затем подключается к подходящему записывающему устройству, например, осциллографу или самописцу. Когда электрод находится вне клетки во внеклеточной среде, регистрируется нулевой потенциал, поскольку внеклеточная среда является изопотенциальной. Однако если электрод проникает в ячейку так, что кончик электрода теперь находится внутри ячейки, на записывающем устройстве наблюдается резкое отклонение. Регистрируется потенциал около -60 милливольт внутри отрицательного по отношению к внешней стороне.Этот потенциал называется потенциалом покоя и остается постоянным в течение неопределенных периодов времени при отсутствии какой-либо стимуляции. Если электрод удалить, снова регистрируется нулевой потенциал. Потенциалы покоя — это не просто характеристики нервных клеток; все клетки тела обладают потенциалом покоя. Что отличает нервные клетки и другие возбудимых мембран (например, мышечные клетки), так это то, что они способны изменять свой потенциал покоя. В случае нервных клеток для интеграции информации и передачи информации, а в случае мышечных клеток — для производства мышечных сокращений.
Потенциалы действия. На рисунке 3 показан еще один эскиз идеализированного нейрона. Этот нейрон был пронзен одним электродом для измерения потенциала покоя и вторым электродом, называемым стимулирующим электродом. Стимулирующий электрод подключается через переключатель к батарее. Если батарея ориентирована так, что положительный полюс подсоединен к переключателю, замыкание переключателя сделает внутреннюю часть элемента несколько более положительной в зависимости от размера батареи.(Такое уменьшение поляризованного состояния мембраны называется деполяризацией ). На рис. 3 представлена анимация, в которой переключатель многократно размыкается и замыкается, и каждый раз, когда он замыкается, в схему подключается батарея большего размера. Первоначально замыкание переключателя вызывает только небольшие деполяризации. Однако потенциалы становятся больше, и в конечном итоге деполяризация становится достаточно большой, чтобы вызвать потенциал действия , также известный как спайк или импульс .Потенциал действия связан с очень быстрой деполяризацией, позволяющей достичь пикового значения около +40 мВ всего за 0,5 миллисекунды (мс). За пиком следует столь же быстрая фаза реполяризации .
Напряжение, при котором деполяризация становится достаточной для запуска потенциала действия, называется порогом . Если для генерации надпороговой деполяризации используется батарея большего размера, единичный потенциал действия все равно генерируется, и амплитуда этого потенциала действия такая же, как и потенциал действия, запускаемый только пороговым стимулом.Простая запись на рисунке 3 иллюстрирует две очень важные особенности потенциалов действия. Во-первых, они вызываются методом по типу по принципу «все или ничего». Либо потенциал действия вызывается стимулами на уровне порога или выше, либо потенциал действия не вызывается. Во-вторых, потенциалы действия — это очень короткие события продолжительностью всего несколько миллисекунд. Возникновение потенциала действия в некоторой степени аналогично подаче спички на предохранитель. Определенная температура необходима для воспламенения предохранителя (т.е.е., предохранитель имеет порог). Спичка, выделяющая большее количество тепла, чем пороговая температура, не приведет к тому, что предохранитель будет гореть ярче или быстрее. Так же, как потенциалы действия вызываются по принципу «все или ничего», они также передаются по принципу «все или ничего». Как только потенциал действия инициируется в одной области нейрона, такой как тело клетки, этот потенциал действия будет распространяться по аксону (подобно горящему предохранителю) и в конечном итоге проникнет в синапс, где он может инициировать процесс синаптической передачи.
В примере на рисунке 3 был сгенерирован только один потенциал действия, потому что продолжительность каждого из двух надпороговых стимулов была настолько короткой, что было достаточно времени только для инициирования одного потенциала действия (т. Е. Стимул закончился раньше, чем потенциал действия завершил свой цикл деполяризации-реполяризации). Но, как показано в анимации на рисунке 4, более продолжительные стимулы могут привести к возникновению множественных потенциалов действия, частота которых зависит от интенсивности стимула.Таким образом, очевидно, что нервная система кодирует информацию не в терминах изменения амплитуды потенциалов действия, а в их частоте. Это очень универсальное свойство. Чем выше интенсивность механического стимула к сенсорному рецептору, тем больше количество потенциалов действия; чем больше степень растяжения рецептора растяжения мышцы, тем больше количество потенциалов действия; чем больше интенсивность света, тем большее количество потенциалов действия передается в центральную нервную систему.Точно так же в двигательной системе, чем больше количество потенциалов действия в двигательном нейроне, тем сильнее будет сокращение мышцы, которая получает синаптическую связь от этого двигательного нейрона. Инженеры называют этот тип кодирования информации частотно-импульсной модуляцией .
Синаптические потенциалы и синаптическая интеграция
На рисунке 5 показаны три нейрона. Единый зеленый цвет будет называться возбуждающим нейроном по причинам, которые вскоре станут ясны.Он устанавливает связь с постсинаптическим нейроном синего цвета. Следы ниже (нажмите «Play») иллюстрируют последствия запуска потенциала действия в зеленом нейроне. Этот потенциал действия в пресинаптическом нейроне приводит к снижению мембранного потенциала постсинаптической клетки. Мембранный потенциал изменяется от значения покоя около -60 милливольт до более деполяризованного состояния. Этот потенциал называется возбуждающим постсинаптическим потенциалом (ВПСП) .Он является «возбуждающим», потому что перемещает мембранный потенциал к пороговому значению, и является «постсинаптическим», потому что это потенциал, зарегистрированный на постсинаптической стороне синапса. Как правило (и это важный момент), единичный потенциал действия в пресинаптической клетке не производит достаточно большой ВПСП, чтобы достичь порога и запустить потенциал действия. Но, если в пресинаптической клетке запускаются множественные потенциалы действия, соответствующие множественные возбуждающие потенциалы могут суммироваться посредством процесса, называемого временным суммированием , чтобы достичь порога и запустить потенциал действия.ВПСП можно рассматривать как «сигнал движения» постсинаптическому нейрону для передачи информации по сетевому пути.
Нейрон, окрашенный в красный цвет на рис. 5, называется тормозным нейроном. Как и зеленый нейрон, он также устанавливает синаптический контакт с синим постсинаптическим нейроном. Он также высвобождает химический передатчик-мессенджер, но последствия связывания передатчика от голубой клетки с рецепторами на постсинаптической клетке противоположны последствиям передатчика, выпущенного зеленым нейроном.Следствием потенциала действия красного пресинаптического нейрона является увеличение мембранного потенциала синего постсинаптического нейрона. Мембранный потенциал более отрицательный, чем был раньше (гиперполяризация ), и поэтому мембранный потенциал дальше от порогового значения. Этот тип потенциала называется ингибирующим постсинаптическим потенциалом (IPSP) , потому что он имеет тенденцию предотвращать активацию постсинаптического нейрона потенциала действия. Это «стоп-сигнал» для постсинаптической клетки.Таким образом, зеленый нейрон говорит «вперед», а красный нейрон говорит «стоп». Что же делать постсинаптическому нейрону?
Нейроны похожи на счетные машины. Они постоянно складывают возбуждающий и тормозящий синаптический вход во времени (временное суммирование) и по площади дендритов, получающих синаптические контакты ( пространственное суммирование) , и если это суммирование находится на пороговом уровне или выше, они запускают потенциал действия. Если сумма ниже порога, потенциал действия не инициируется.Этот процесс называется синаптическая интеграция и проиллюстрирован на рисунке 5. Первоначально два потенциала действия в зеленом нейроне производили суммирующие ВПСП, которые запускали потенциал действия в синем нейроне. Но если IPSP от тормозного нейрона возникает непосредственно перед двумя потенциалами действия в возбуждающем нейроне, сумма одного IPSP и двух EPSP ниже порогового значения, и в постсинаптической клетке не возникает никакого потенциала действия. Тормозящий нейрон (и подавление в целом) — это способ стробирования или регулирования способности возбуждающего сигнала запускать постсинаптическую клетку.
Нейронные сети
Мотивы микросетей
Как указывалось ранее в этой главе, нейрон может получать контакты до 10 000 пресинаптических нейронов, и, в свою очередь, любой нейрон может контактировать до 10 000 постсинаптических нейронов. Комбинаторная возможность может привести к чрезвычайно сложным нейронным цепям или сетевым топологиям , которые могут быть очень трудными для понимания. Но, несмотря на потенциально огромную сложность, многое можно узнать о функционировании нейронных цепей, исследуя свойства подмножества простых конфигураций цепей.На рисунке 6 показаны некоторые из этих мотивов микросхем или микросетей. Несмотря на простоту, они могут делать многое из того, что должна делать нервная система.
Возбуждение с прямой связью . Позволяет одному нейрону передавать информацию своему соседу. Их длинные цепочки можно использовать для передачи информации через нервную систему.
Запрещение прямой связи .Пресинаптическая клетка возбуждает тормозящий интернейрон (интернейрон — это нейрон, расположенный между двумя нейронами), и этот тормозной интернейрон затем подавляет следующую клетку-последователь. Это способ выключить или ограничить возбуждение в нейроне ниже по потоку в нейронной цепи.
Схождение / расхождение . Одна постсинаптическая клетка получает конвергентный вход от ряда различных пресинаптических клеток, и любой отдельный нейрон может создавать дивергентные связи со многими разными постсинаптическими клетками.Дивергенция позволяет одному нейрону взаимодействовать со многими другими нейронами в сети. Конвергенция позволяет нейрону получать входные данные от многих нейронов в сети.
Боковое торможение . Пресинаптическая клетка возбуждает тормозящие интернейроны, и они подавляют соседние клетки в сети. Как подробно описано ниже в этой главе, этот тип схемы может использоваться в сенсорных системах для улучшения контуров.
Запрет обратной связи / повторения .На панели E1 пресинаптическая клетка соединяется с постсинаптической клеткой, а постсинаптическая клетка, в свою очередь, соединяется с интернейроном, который затем подавляет пресинаптическую клетку. Этот контур может ограничивать возбуждение в пути. Некоторое начальное возбуждение будет отключено после того, как красный интернейрон станет активным. На панели E2 каждый нейрон в замкнутой цепи подавляет нейрон, с которым он связан. Казалось бы, эта схема ничего не делает, но, как будет показано далее в этой главе, она может привести к генерации сложных паттернов всплесковой активности.
Обратная связь / рекуррентное возбуждение . На панели F1 пресинаптический нейрон возбуждает постсинаптический нейрон, а этот постсинаптический нейрон возбуждает пресинаптический нейрон. Этот тип цепи может выполнять функцию переключателя, потому что после активации пресинаптической клетки эта активация может сохраняться. Активация пресинаптического нейрона могла включить эту сеть, и она могла остаться. На панели F2 показаны варианты возбуждения по обратной связи, при которых пресинаптический нейрон возбуждает постсинаптический нейрон, который может по обратной связи возбуждать сам себя (a, autapse ) или другие нейроны, которые в конечном итоге имеют обратную связь (b) с самим собой.
Эти простые мотивы — вездесущие компоненты многих нейронных цепей. Давайте рассмотрим несколько примеров того, что могут делать эти сети.
Возбуждение с прямой связью и подавление с прямой связью
Одна из наиболее изученных микросхем — это схема, которая обеспечивает простое рефлекторное поведение. На рисунке 7 показана схема так называемого рефлекса рефлекса коленного рефлекса или растяжения. Невролог ударяет по колену резиновым молоточком, в результате чего нога разгибается.Этот тест используется как простой способ проверить целостность некоторых сенсорных и моторных путей в спинном мозге. Удар молотка растягивает мышцу и приводит к возбуждению потенциалов действия в сенсорных нейронах в мышце, которые чувствительны к растяжению. (Потенциалы действия представлены маленькими яркими «огоньками» на анимации.) Потенциалы действия инициируются по принципу «все или ничего» и распространяются в спинной мозг, где аксон разделяется (раздваивается) на две ветви.
Давайте сначала обсудим ветвь слева, которая формирует синаптическую связь (зеленый треугольник) с Extensor (E) моторным нейроном (окрашен в синий цвет). Потенциал действия сенсорного нейрона вторгается в синаптический конец сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение двигательного нейрона. Растяжение мышцы приводит к возникновению потенциала действия в двигательном нейроне (МН), который затем распространяется по периферическому нерву и вторгается в синапс в мышце, вызывая высвобождение передатчика и потенциал действия в мышце.Потенциал действия в мышечной клетке приводит к сокращению мышцы и разгибанию конечности. Итак, здесь у нас есть простая схема возбуждения с прямой связью, которая опосредует поведение.
Теперь давайте рассмотрим правую ветвь аксона сенсорного нейрона на рисунке 7. Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение постсинаптического интернейрона, окрашенного в черный цвет.Этот нейрон называется интернейроном , потому что он расположен между одним нейроном (здесь SN) и другим нейроном (здесь MN). Возбуждение интернейрона приводит к инициированию действия и последующему высвобождению передатчика из пресинаптического терминала интернейрона (черный треугольник), но для этой ветви цепи передатчик ведет к IPSP в постсинаптическом сгибателе (F ) мотонейрон (окрашен в красный цвет). Функциональные последствия этого подавления с прямой связью заключаются в снижении вероятности того, что двигательный нейрон-сгибатель станет активным и вызовет несоответствующее сгибание ноги.
Сходимость и расхождение
Упрощенная схема, опосредующая рефлекс растяжения, представлена на рисунке 8. Однако правильное функционирование схемы рефлекса растяжения также зависит от конвергенции и расхождения. У одного сенсора есть несколько ветвей, которые расходятся и образуют синаптические связи со многими отдельными моторными нейронами (щелкните «Дивергенция»). Следовательно, когда мышца сокращается в результате нажатия невролога, это происходит потому, что несколько мышечных волокон активируются одновременно несколькими двигательными нейронами.Кроме того, когда мышца растягивается, активируется не один, а несколько сенсорных нейронов, и все эти сенсорные нейроны проецируются в спинной мозг, где они сходятся к отдельным моторным нейронам-разгибателям (нажмите «Конвергенция»). Итак, рефлекс растяжения возникает из-за комбинированных эффектов активации нескольких сенсорных нейронов и моторных нейронов-разгибателей.
Боковое торможение
Улучшение кромки.Боковое торможение очень важно для обработки сенсорной информации. Одним из примеров является явление в зрительной системе, называемое усилением краев. На рисунке 9 показаны две полосы: темно-серая полоса слева и светло-серая полоса справа. Хотя темная полоса и светлая полоса имеют равномерную яркость по всему полю, при внимательном рассмотрении обнаруживается, что светло-серая полоса кажется несколько светлее на границе темно-серой полосы, чем в других областях поля.Напротив, темно-серая полоса на границе выглядит несколько темнее, чем в других областях темного поля. Это явление усиления границ, которое помогает визуальной системе извлекать важную информацию из визуальных сцен. Усиление краев опосредуется, по крайней мере частично, боковым торможением в сетчатке.
Давайте сначала рассмотрим схему без бокового торможения (Рисунок 10, щелкните «Без бокового торможения»). Свет падает на сетчатку (Часть A), а его интенсивность можно описать ступенчатым градиентом (Часть B).В качестве упрощения предположим, что темно-серая область имеет интенсивность пяти единиц, а светло-серая область — десять единиц. Градиент света активирует фоторецепторы, и фоторецепторы создают синаптические связи с нейронами второго порядка. Предположим, что интенсивность света 5 единиц приводит к 5 спайкам / с, а интенсивность света 10 единиц приводит к 10 спайкам / с (Часть C) в фоторецепторах, и что синаптическая сила достаточна (здесь обозначена как +1) так что интенсивность света 5 единиц приводит к 5 импульсам / с, а интенсивность света 10 единиц приводит к 10 импульсам / с (Часть C) соответственно в нейронах второго порядка.Если никакой дальнейшей обработки информации не происходит, воспринимаемый градиент будет точно таким же, как градиент интенсивности света (часть B, красный след). Но это не то, что воспринимается, и латеральное торможение объясняет разницу.
Теперь рассмотрим расширенную схему с боковым торможением (нажмите «С боковым торможением»). Каждый из фоторецепторов устанавливает тормозные синаптические связи со своим соседним нейроном второго порядка. Сила торможения (обозначается -0.2) меньше силы возбуждения (пожертвовано +1). Прежде чем смотреть на границу, рассмотрите выход схемы в однородных областях каждого поля. Далеко справа от границы все клетки получают одинаковое возбуждение и одинаковое торможение. Без латерального торможения интенсивность света в 10 единиц произвела бы 10 спайков в секунду в нейроне второго порядка. Но из-за тормозящего соединения со стороны соседних нейронов справа и слева выходная мощность снижается до 6 импульсов в секунду.То же самое верно для клеток далеко слева от границы, но величина возбуждения меньше и, соответственно, величина торможения меньше. Ключевая обработка происходит на границе или краю. Обратите внимание, что нейрон справа от границы получает такое же подавление от нейрона справа, но получает меньшее подавление от нейрона слева от него по другую сторону границы. Следовательно, он получает больше чистого возбуждения и имеет выходной сигнал 7 пиков / с, а не 6 пиков / с его соседа справа.Теперь посмотрите на нейрон слева от границы. Он получает слабое торможение от своего соседа слева, но более сильное торможение от своего соседа справа по другую сторону границы. Следовательно, он получает меньше чистого возбуждения и имеет выходную мощность 2 пика / с, а не 3 пика / с его соседа слева. Таким образом, в результате бокового торможения информация, передаваемая в нервную систему, и воспринимаемый градиент будут версией исходной с усиленной границей или краем (рис. 10B)!
Диапазон
Маха.Простой контур сетчатки с боковым торможением может объяснить феномен усиления краев. Этим также можно объяснить визуальную иллюзию, известную как полосы Маха. На рисунке 11 показан градиент светлых и темных вертикальных полос, поперек которых проходит тонкая горизонтальная линия. Похоже, что горизонтальная линия имеет неравномерное распределение интенсивности: она темнее в области светлых вертикальных градиентов и светлее в области темных вертикальных градиентов. Это визуальная иллюзия .Иллюзию можно раскрыть, наложив маску на вертикальный градиент. (Нажмите «Play», чтобы добавить маску.) Теперь вы можете видеть, что горизонтальная полоса имеет равномерную интенсивность. В некоторых областях он воспринимается более темным, потому что клетки сетчатки, которые реагируют на более темную область горизонтальной полосы, сильно подавляются клетками, реагирующими на яркую область вертикальной полосы. Напротив, полоса воспринимается более яркой в некоторых областях, потому что клетки сетчатки, которые реагируют на более светлую область горизонтальной полосы, только слабо ингибируются клетками, реагирующими на темную область вертикальной полосы.
Обратная связь / запрет повторения
Запрет обратной связи в микросхемах. Подавление обратной связи играет общую роль в гашении возбуждения через нейронную цепь. Классическим примером является клетка Renshaw в спинном мозге. Аксон ветви спинномозгового мотонейрона. Одна ветвь иннервирует мышцу, как описано ранее (например, рис. 7), а другая ветвь создает возбуждающую синаптическую связь с интернейроном, называемым клеткой Реншоу.Интернейрон, в свою очередь, подавляет двигательный нейрон, замыкая петлю. Другой пример подавления обратной связи обнаружен в гиппокампе. Пирамидные клетки СА3-типа создают возбуждающие связи с клетками корзины и обратную связь клеток корзины, чтобы ингибировать клетки СА3. Термин повторяющееся ингибирование применяется к простым схемам подавления обратной связи, таким как цепь Реншоу в спинном мозге и цепь корзиночных клеток в гиппокампе.
Подавление обратной связи в наноцепях.Подавление обратной связи распространено не только во многих нейронных цепях; он также распространен в биохимических цепях. Здесь он может служить подложкой для генерации колебаний. Они могут охватывать множество временных масштабов от секунд до дней в зависимости от молекулярных компонентов схемы.
Рис. 12. Из Byrne, Canavier, Lechner, Clark and Baxter, 1996. |
- Эндогенное взрывное поведение в нейронах.Идеализированный нейрон, описанный ранее в этой главе, молчал в отсутствие стимуляции (например, рисунок 3). Однако некоторые нейроны активируют потенциалы действия в отсутствие стимуляции, и, в некоторых случаях, паттерны возбуждения могут демонстрировать взрывной паттерн, в котором последовательные высокочастотные всплески активности сменяются периодами покоя. Такие нейронные свойства могут быть важны для создания ритмического поведения, такого как дыхание. На рис. 12 показан пример записи нейрона беспозвоночного, имеющего эндогенный импульсный ритм.Этот конкретный нейрон называется параболическим всплескивающим устройством, потому что характер интервалов между всплесками является длинным в начале и конце цикла всплеска, но очень коротким в середине цикла. Клетка запускает всплеск потенциалов действия, а затем замолкает, но вскоре происходит еще один всплеск, и этот процесс продолжается бесконечно каждые десять-пятнадцать секунд. Взрыв происходит, даже если нейрон хирургическим путем удален из ганглия и помещен в культуру, чтобы не было синаптических связей с другими нейронами.Таким образом, для этого ритма не нужна нейронная сеть — она эндогенная. Но это действительно связано с наноконтуром внутри клетки. На рисунке 13А показана очень упрощенная версия этой сети, в которой подчеркивается ключевой принцип работы. Критическим для этой сетевой функции является канал в
мембрана (обозначена g SI ), проницаемая для Ca 2+ . Поскольку концентрация Ca 2+ относительно высока во внеклеточной среде и низка внутри клетки, Ca 2+ будет двигаться вниз по градиенту концентрации и тем самым деполяризует клетку.В конце концов, деполяризация достигает порога, и клетка начинает срабатывать. Поджигание приводит к дополнительному притоку Ca 2+ (зеленая стрелка) и накоплению Ca 2+ внутри клетки. Ключевым этапом является то, что накопление Ca 2+ ингибирует (красная стрелка) дальнейший приток кальция и прекращает всплеск. Всплеск остается прекращенным, пока уровни внутриклеточного Ca 2+ остаются повышенными. Но уровни Ca 2+ не остаются повышенными надолго.Они восстанавливаются внутриклеточными буферами и удаляются из клетки с помощью насосов (I NaCa и I CaP ). Когда внутриклеточные уровни Ca 2+ снижаются, ингибирование канала снимается ( растормаживание ), и нейрон снова начинает деполяризоваться, и начинается новый всплеск. По сути, то, что мы видим здесь на уровне наносети, является повторением сети, препятствующей обратной связи (рис. 13B). Первоначальный процесс возбуждения приводит к активации тормозного процесса, который отключается, чтобы отключить процесс возбуждения.В такой сети возникнут колебания, если возбуждающее побуждение будет непрерывным, но тормозящий процесс будет терять свою эффективность.
Рисунок 14. По материалам Hastings et al., Nature Rev. Neurosci. , 2003.
Циркадные ритмы.Второй пример подавления обратной связи — это наноцепь регуляции генов, лежащих в основе циркадных ритмов. Циркадный ритм позвоночных обусловлен работой группы нейронов в области мозга, называемой супрахиазматическим ядром, которая расположена чуть выше зрительного нерва. Эти нейроны оказывают сильное влияние как на высвобождение гормонов, таких как мелатонин, кортизол, так и на вегетативные функции, такие как температура тела (рис. 14). Несмотря на огромное влияние этого генератора, его работа сводится к очень простой схеме, и действительно, не к нейронной схеме, а к другой наноконтуре.Основной механизм, по-видимому, сохраняется у всех видов животных, включая человека. На рисунке 15 представлена упрощенная принципиальная схема основных компонентов. Участвует несколько генов, но основной механизм включает ген под названием на , где на — на период. Этот ген был впервые идентифицирован у плодовой мухи Drosophila , но также присутствует у позвоночных. Ген на приводит к производству на матричной РНК. на мРНК покидает ядро и попадает в цитоплазму, где приводит к синтезу белка PER.PER диффундирует или транспортируется обратно в ядро, где репрессирует дальнейшую транскрипцию гена на . Концептуально эта система очень похожа на механизм разрывного нейрона, рассмотренный выше. Ген активируется, он производит сообщение и белок, а белковая обратная связь подавляет экспрессию гена. Но как цикл повторяется? Ключевым механизмом является деградация PER. Белок PER разлагается и разлагается в течение 24 часов.Таким образом, когда белок PER разрушается, подавление или репрессия снимается (растормаживание), позволяя этому гену снова начать создавать информационную РНК и белок. Итак, как только этот цикл начинается, он повторяется снова и снова через 24 часа. Это основной механизм, лежащий в основе циркадных ритмов и мощного воздействия, которое они оказывают на ряд различных физиологических систем. По сути, все наши циркадные ритмы начинаются с сети подавления молекулярной обратной связи.
Запрет обратной связи в кольцевых цепях.Повторяющееся торможение может, по крайней мере в принципе, объяснить генерацию сложных моторных паттернов, примером которых является передвижение на четвероногих ногах. Четвероногие животные интересны тем, что четвероногие способны не только двигать четырьмя ногами, но и генерировать различные типы циклов активности, называемые походками. На рисунке 16 показаны четыре аллюра. Первая панель — это прогулка (тап по собаке). Последовательность начинается с разгибания левой передней конечности. Затем следуют разгибания правой задней конечности, правой передней конечности и левой задней конечности.На рыси (вторая панель рисунка 16) (постукивание по собаке) левая передняя и правая задние конечности находятся в фазе друг с другом и на 180 градусов не совпадают по фазе с правой передней и левой задней конечностями. В связке (третья панель) (постукивание по собаке) левая передняя и правая передние конечности находятся в фазе, но на 180 градусов не совпадают по фазе с левой задней и задней задними конечностями. Галоп (четвертая панель) (постукивание по собаке) — вариант скачек, при котором есть небольшая разница фаз между правой и левой передними и задними конечностями.
Как нервная система генерирует эти походки? И необходимы ли для каждой отдельные нейронные цепи? К сожалению, нейробиологи не знают ответов на эти вопросы, но было бы полезно изучить некоторые возможности. Это подход в области нейробиологии под названием Вычислительная и теоретическая нейробиология . Один из способов создания походки проиллюстрирован на рисунке 17. Возьмите четыре отдельных нейрона, каждый из которых обладает эндогенной импульсной активностью, как показано ранее на рисунке 12, и назначьте активность в каждом из этих нейронов контролю конкретной конечности.Нейроны могут быть «запущены» так, чтобы они имели соответствующие фазовые соотношения для генерации походки, такие как граница, показанная на рисунке 16. Трудность будет заключаться в запуске нейронов точно в точное время. Другой проблемой могут быть небольшие «дрейфы» в колебательных периодах четырех независимых нейронов, которые со временем приведут к тому, что паттерн станет несогласованным (Рисунок 18). Эта собака не выиграет ни одной гонки и, вероятно, не сможет ходить.
Итак, очевидно, что нейроны должны быть связаны.Один из способов сделать это — использовать повторяющуюся цепь подавления, состоящую из четырех связанных нейронов, чтобы сформировать так называемую «кольцевую» схему, где каждый нейрон в цепи имеет эндогенную импульсную активность, и каждый нейрон связан с другим с помощью тормозной синаптической связи ( Рисунок 19A).
Рис. 19. По материалам Canavier, Butera, Dror, Baxter, Clark and Byrne, 1997. |
Чтобы получить правильные фазовые соотношения для походки, вместо того, чтобы назначать нейрон 3 правую переднюю конечность, он назначается для управления правой задней конечностью, а нейрон 4 назначается для управления правой передней конечностью ( простой поворот схемы) (рис. 19Б).При реализации с помощью компьютерного моделирования эта единственная схема способна генерировать четвероногие ворота. Более того, одна и та же схема с небольшими изменениями свойств отдельных нейронов может генерировать каждую из четырех походок, показанных на рисунке 17 (рисунок 20).
Этот результат указывает на важный момент в отношении нейронных сетей.Чтобы понять их, необходимо понимать не только топологию сети, но и природу связей между нейронами (возбуждающие они или тормозящие), а также свойства отдельных узлов (т. Е. нейроны). Кроме того, это моделирование иллюстрирует явление, называемое динамической реконфигурацией . Для генерации этих четырех разных походок не обязательно иметь четыре разные сети — все это можно сделать с помощью одной схемы.Фактическая схема, генерирующая квадрапедальную походку, более сложна, чем схема на рис. 19. Заинтересованный читатель может обратиться к недавнему обзору Оле Кейна (см. Дополнительную литературу).
Обратная связь / рекуррентное возбуждение
Периодическое возбуждение в нано-схемах и микросхемах, по-видимому, имеет решающее значение для процессов обучения и памяти. Обучение включает изменения биофизических свойств нейронов и изменения синаптической силы. Накапливающиеся данные указывают на то, что положительная обратная связь внутри биохимических каскадов и генных сетей является важным компонентом для индукции и поддержания этих изменений.Более того, периодическое возбуждение обнаруживается по крайней мере в некоторых микросхемах, участвующих в процессах памяти. Яркий пример находится в области CA3 гиппокампа.
Рисунок 21. Изменено из Byrne and Roberts, 2009. |
На рисунке 21 показаны ключевые особенности рекуррентной возбуждающей цепи CA3. Шесть различных пирамидных нейронов гиппокампа обозначены как U , V , W , X , Y и Z .Каждый из этих нейронов получает синаптическую связь от пресинаптических нейронов, обозначенных a , b , c , d , e и f. Эти пресинаптические нейроны могут быть активными или неактивными с 0 и черным цветом, представляющим неактивный нейрон, и 1 и зеленым цветом, указывающим активный. Важным аспектом этой схемы является то, что синаптические связи от входного пути достаточно сильны, чтобы активировать (запустить) пирамидный нейрон, с которым они связаны.Например, если активирован нейрон a , будет активирован нейрон Z , который представлен как 1 на панели вывода. Эта топология — не что иное, как возбуждение с прямой связью. Взаимное возбуждение делает эту схему особенной. Например, нейрон Z и другие пирамидные нейроны имеют коллатерали аксонов, которые посредством обратной связи соединяются друг с другом. Но они не только устанавливают связь между собой. Каждый нейрон устанавливает связь с каждым из четырех других пирамидных нейронов в цепи.Таким образом, каждая пирамида получает сходящуюся информацию от всех других клеток в сети, и, в свою очередь, выход каждого пирамидного нейрона расходится, чтобы создать синаптические связи со всеми другими пирамидными нейронами в цепи. (Следовательно, этот повторяющийся мотив возбуждения вложил в себя мотивы конвергенции и расхождения.) Матрица связности состоит из 36 элементов.
Для того, чтобы эта сеть могла чему-либо научиться, в схему необходимо встроить правило обучения синаптической пластичности.Один из широко распространенных правил известен как Hebb Learning Rule . По сути, он заявляет, что синапс изменит свою силу, если этот синапс активен (то есть высвобождает передатчик), и в то же время активна постсинаптическая клетка. Комбинация этого правила обучения и повторяющегося возбуждающего контура приводит к некоторым интересным эмерджентным свойствам. Например, если нейрон Z активирован входом a, , сила его соединения с самим собой (синапс 1 ) изменится, как показано зеленым синапсом на рисунке 21 (анимация находится в 2-секундном цикле до иллюстрируют контрольные и модифицированные синапсы).Однако синапс 1 не будет единственным синапсом, который будет усилен. Например, синапс 13 также будет усилен, потому что нейрон Z был активен в то же время, когда нейрон X был активирован входом c . Напротив, синапс 7 не усилен, потому что нейрон Y не был активен в то же время, что и нейрон Z . Конечный результат этой конвергенции и расхождения и правила обучения состоит в том, что исходный входной шаблон активности будет сохранен как изменения в элементах матрицы связности.Поэтому эта схема получила название сети с автоматической ассоциацией . Важная концепция здесь состоит в том, что «память» не находится ни в одном синапсе; это распределенных в сети.
Сводка
Значительный прогресс был достигнут в понимании того, как разные простые нейронные сети участвуют в обработке информации и посредническом поведении. Возбуждение с прямой связью и торможение с прямой связью опосредуют рефлекторное поведение.Боковое торможение важно для усиления краев. Повторяющееся возбуждение — важный механизм памяти. Повторяющееся торможение может иметь важное значение для формирования двигательного поведения. Сходимость и расхождение заложены в этих микросхемах. Одни и те же типы сетевых мотивов воспроизводятся в биохимических и генных сетях.
Следующий уровень понимания находится на уровне нейронных сетей, которые опосредуют более сложные, так называемые функции мозга высшего порядка.Их понимание становится возможным благодаря использованию электрофизиологических и оптических методов записи и современных методов визуализации, таких как функциональная магнитно-резонансная томография (фМРТ) и диффузионно-тензорная визуализация (DTI) . ФМРТ позволяет исследователям идентифицировать области мозга, которые заняты когнитивными задачами, тогда как DTI позволяет визуализировать пути, связывающие одну область мозга с другой. Рисунок 22 (любезно предоставлено Тимом Элмором, доктором философии, отдел.из нейрохирургии, Медицинская школа Техасского университета в Хьюстоне) — это вид сбоку человеческого мозга, показывающий пути, соединяющие корковые области, выявленные с помощью DTI !.
|
Рисунок 22. Предоставлено Тимом Эллмором, доктором философии. |
Распознавание объекта — это пример прогресса в понимании макросхем.Как показано на рисунке 23, обработка визуальной информации начинается в сетчатке, а затем задействуется несколько областей коры головного мозга, такие как затылочная кора и височная кора. Внутри этой макросхемы находятся модули, извлекающие информацию более высокого порядка. Предположительно, каждый модуль включает в себя сотни, если не тысячи отдельных микросхем. Задача на будущее — определить, как эти модули работают и как они взаимодействуют с другими модулями. Хотя соединения с прямой связью присутствуют, соединения обратной связи и боковые соединения широко распространены.Задача огромна, но, возможно, достижению цели будет способствовать использование преимуществ того, что было изучено о принципах работы наносхем и микросхем. Чтобы понять макросхемы, необходимо знать больше, чем топологию сетевых соединений. Необходимо будет знать, как работает каждый модуль, и о динамике межмодульных соединений.
Рисунок 23.Из Феллемана и Ван Эссена, 1991. |
Проверьте свои знания
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью Этот ответ НЕПРАВИЛЬНЫЙ.
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
А.Прямое возбуждение
B. Запрещение прямой связи. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Запрет обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
с.Запрет обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое торможение. Ответ ПРАВИЛЬНЫЙ!
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E. Расхождение. Этот ответ НЕПРАВИЛЬНЫЙ.
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция. Этот ответ ПРАВИЛЬНЫЙ!
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция Этот ответ НЕПРАВИЛЬНЫЙ.
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание. Этот ответ НЕПРАВИЛЬНЫЙ.
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
Б. Пространственное суммирование
C. Запрет обратной связи
Д.Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи. Ответ ПРАВИЛЬНЫЙ!
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
Д.Возбуждение обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение. Ответ НЕПРАВИЛЬНЫЙ.
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение кромки Этот ответ НЕПРАВИЛЬНЫЙ.
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
B. Улучшение кромки
C. Память. Ответ ПРАВИЛЬНЫЙ!
D. Передвижение
E.Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
Б.Улучшение края
C. Память
D. Передвижение
E. Колебания нейронов. Этот ответ НЕПРАВИЛЬНЫЙ.
пожертвований Neuroscience Online поможет финансировать разработку новых функций и контента. |
Что такое пирамидальный нейрон? (с изображениями)
В центральной нервной системе пирамидный нейрон — это тип нервной клетки, названный так из-за формы ее клеточного тела.Тело клетки, или сома, пирамидных нейронов имеет отчетливую форму, которая дала им название. Их также называют пирамидными клетками, и они были обнаружены у рыб, птиц, рептилий и млекопитающих. Пирамидный нейрон имеет ту же базовую структуру, что и все другие типы нейронов, поэтому он состоит из тела клетки, прикрепленного к аксону и дендритам. Уникальным пирамидальный нейрон делает расположение дендритов и тот факт, что и аксон, и дендриты подвергаются обширному ветвлению.
Различные типы нейронов.
В центральной нервной системе пирамидные нейроны обнаруживаются только в различных областях переднего мозга. В частности, эти типы нейронов отправляют и получают нервные импульсы в коре головного мозга, гиппокампе и миндалевидном теле. Считается, что у млекопитающих пирамидные нейроны играют ключевую роль в когнитивных функциях, таких как восприятие, рассуждение, запоминание, мышление и понимание.
Пирамидные нейроны обнаруживаются только в областях переднего мозга.
Пирамидный нейрон имеет две отдельные группы дендритов: одна находится в нижней части тела клетки, а другая — в верхней части тела клетки. Эти группы дендритов называются базальными и апикальными дендритами соответственно. Это основная структура пирамидного нейрона, и она может варьироваться в зависимости от того, где находится клетка в центральной нервной системе.Мало того, что структура пирамидных нейронов может различаться в зависимости от того, в какой части мозга они находятся, но также и от точного местоположения, в котором они находятся внутри структуры.
В основании пирамидных нейронов обычно есть несколько базальных дендритов, которые широко разветвляются.Таким образом, базальное дендритное дерево больше похоже на куст с множеством ветвей. От вершины тела клетки один апикальный дендрит обычно выходит на некоторое расстояние до того, как разветвляется. В некоторых случаях этот дендрит может расщепляться на два отдельных дендрита, которые расширяются и разветвляются на конце.
Каждый пирамидальный нейрон содержит множество дендритов, которые значительно разветвлены, поэтому он может взаимодействовать с сотнями и даже тысячами других нейронов.Многие исследования изучали, как нервные импульсы принимаются пирамидными нейронами. Это не совсем понятно, поскольку нервные импульсы часто принимаются одним или другим дендритным деревом, а также могут быть получены определенными частями каждого дерева.
PPT — 1 — ДОПОЛНИТЕЛЬНАЯ ПИРАМИДНАЯ СИСТЕМА 2 — ПОВРЕЖДЕНИЯ ДВИГАТЕЛЬНЫХ НЕЙРОНОВ PowerPoint Presentation
1 — ДОПОЛНИТЕЛЬНАЯ ПИРАМИДНАЯ СИСТЕМА 2 — ПОВРЕЖДЕНИЯ ДВИГАТЕЛЯ НЕЙРОНА тракты известны как ДОПОЛНИТЕЛЬНЫЕ ПИРАМИДНЫЕ ТРАКТЫ.
КОМПОНЕНТЫ ЭКСТРАПИРАМИДНОЙ СИСТЕМЫ • БАЗАЛЬНАЯ ГАНГЛИЯ • МОЗГОВАЯ СТЕМА Дает начало следующим трактам: • Руброспинальный тракт • Вестибулоспинальный тракт • Ретикуло-спинальный тракт • Тектоспинальный тракт
Мозг
43 руб. — Красное ядро получает вход как от мозжечка, так и от моторных областей коры головного мозга. • Выходной сигнал — через руброспинальный тракт направляется к контрлатеральным моторным нейронам спинного мозга (пересекает противоположную сторону на уровне ядра, а аксоны расположены в латеральном белом веществе спинного мозга перед кортикоспинальным трактом.• Функции — участвуют в движениях дистальных отделов конечностей (рук и ног), также регулируют тонус и позу. • Он возбуждает сгибатели и подавляет мышцы-разгибатели.
ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ • Расположение — вестибулярные ядра расположены в мосту и мозговом веществе • Вход — они получают входной сигнал от вестибулярного аппарата внутреннего уха и мозжечка • Выходной сигнал — в основном от боковых вестибулярных ядер к спинному мозгу в вестибулоспинальном тракте. Остается ипсилатеральный • Функция — возбуждает ипсилатеральный разгибатель.Тормозит мышцы-сгибатели • Регулирует мышечный тонус для поддержания баланса в ответ на движение головы
РЕТИКУЛОСПИНАЛЬНЫЙ ТРАКТ • Расположение — Ретикулярная формация в центральном сером веществе ствола головного мозга • Входные данные — Афферентный вход в ретикулярную формацию поступает из спинного мозга, вестибулярные ядра, мозжечок, сенсомоторная кора, бледный шар и лат. Гипоталамус • Выход — нисходящий тракт возникает из ядер в мосту и мозговом веществе 1] Мост — мост Ретикулоспинальный тракт проходит ипсилатерально; Функция — Возбуждает осевые мышцы-разгибатели 2] Медулла — Медуллярный ретикулоспинальный тракт проходит ипсилатерально (некоторые также перекрещиваются). Функция — Тормозит по отношению к аксиальному разгибателю. стимулы • Выход — передает нервные импульсы от верхнего бугорка (среднего мозга) к контралатеральным скелетным мышцам, которые двигают голову и глаза в ответ на зрительные стимулы • Функция — участвует в контроле мышц шеи в ответ на зрительные стимулы
СПУСКАЮЩИЙСЯ ДОПОЛНИТЕЛЬНЫЙ ПИРАМИД ДВИГАТЕЛЬНЫЙ ТРАКТ К ВНУТРЕННЕМУ ПОЗВОНОЧНИКУ И ДВИГАТЕЛЬНЫЙ НЕЙРОН и без пересечения Эти пути заканчиваются на интернейронах передних рогов.Иногда они оканчиваются непосредственно на двигательных нейронах передних рогов.
РЕЗЮМЕ ПИРАМИДНЫХ И ЭКСТРАПИРАМИДНЫХ ПУТЕЙ
ПОПЕРЕЧНЫЙ РАЗДЕЛ СПИННОГО МОЗГА
8
СИСТЕМА СИСТЕМЫ
ВНЕШНЯЯ ЧАСТЬ
СИСТЕМА СИСТЕМЫ . многие области мозга • Конечным звеном экстрапирамидного пути является ствол мозга, на который влияет моторная кора, мозжечок, базальные ядра [поэтому эти области мозга косвенно регулируют двигательную активность] Примечание. Прямое влияние на переднюю домашнюю клетку осуществляется первичной моторной корой
ФУНКЦИИ ДОПОЛНИТЕЛЬНОЙ ПИРАМИДАЛЬНОЙ СИСТЕМЫ МУЛЬТИНЕЙРОНАЛЬНАЯ СИСТЕМА • РЕГУЛИРОВАНИЕ ПОЛОЖЕНИЯ ТЕЛА, ВКЛЮЧАЯ НЕДОБРОВОЛЬНЫЕ ДВИЖЕНИЯ БОЛЬШИХ ГРУПП МЫШЕЧНИКОВ И КОНЕЧНОСТЕЙ • РЕГУЛИРОВАНИЕ ДОБРОВОЛЬНЫХ ДВИЖЕНИЙ 9105 • Имеется
005. между Пайрой мидальные и экстрапирамидальные системы, например, при выполнении тонкой работы, такой как работа иглой (пирамидная система), человек должен подсознательно принять определенное положение рук (экстрапирамидальная система), которая позволяет выполнять вашу работу
ПРИМЕНЯЕТСЯ • Экстра пирамидальные тракты некоторые из них возбуждают, а другие подавляют общий эффект мышечного тонуса — сильный тормозящий эффект по отношению к гамма-моторному нейрону в клетке переднего рога. Каким будет эффект от экстрапирамидных поражений? • Гипертония Почему? — Потому что сильный тормозящий эффект на гамма мотонейрон теряется.
Разница между пирамидными и экстрапирамидными путями • ДОПОЛНИТЕЛЬНЫЕ ПИРАМИДНЫЕ ТРАКТЫ • -Руброспинальный • -Вестибулоспинальный • -Ретикулоспинальный • -Тектоспинальный • Они берут начало в среднем мозге и ядрах ствола головного мозга и имеют влияние на базальные ганглии и коры головного мозга стимулировать или ингибировать эти ядра • Нет прямого контроля моторной коры или базальных ганглиев на спинном мозге, но через ядра в среднем и стволе мозга ПИРАМИДНЫЕ ТРАКТЫ • -Боковой кортикоспинальный -Ant.или вентральная кортикоспинальная — кортикобульбарная • Тела клеток, которые участвуют в пирамидных путях, расположены в прецентральной извилине (первичная, премоторная и дополнительная моторная кора) и соматосенсорной области. • Пирамидный тракт спускается напрямую, без синаптических перерывов от моторной коры головного мозга к спинному мозгу (на интернейрон и клетки муравьиного рога)
Различие между пирамидными и экстрапирамидными путями Продолжение. . . • ВНЕПИРАМИДНЫЕ ПУТИ • Основные экстрапирамидные тракты, некоторые из которых пересекаются, а другие не пересекаются (см. Таблицу, приведенную выше) • Функция: • Контроль положения тела, включающий непроизвольные движения осевых и проксимальных мышц конечностей. ПИРАМИДНЫЕ ТРАКТЫ • 80% кортикоспинальных трактов (латеральные) пересекается в мозговом веществе. 20% кортикоспинального тракта (вентрального) пересекается в спинном мозге Из-за пересечения коры головного мозга контролирует противоположную сторону тела • Функция: — Шир.Кортикоспинальный тракт — мелкие движения пальцев, напр. Письмо, работа иглой — Вентральный кортикоспинальный тракт — Осевое или постуральное движение
ОБЩИЕ СЛОВА, ИСПОЛЬЗУЕМЫЕ В НЕЙРОФИЗИОЛОГИИ И КЛИНИЧЕСКОЙ НЕВРОЛОГИИ • ПИРАМИДНЫЕ ТРАКТЫ • ЭКСТРАПИРАМИДНЫЕ ТРАКТЫ • Экстрапирамидальные пирамидальные пути • ПИОНАМИДАЛЬНЫЕ ЛИЦА ПОВРЕЖДЕНИЯ — ГИПЕРТОНИЯ (ригидность) • UMN (Верхний мотонейрон) — Двигательные тракты, идущие от мозга к муравью. Роговые клетки [Пирамидные и экстрапирамидные тракты]
Двигательная система Это система из двух нейронов 1 — Верхний мотонейрон — От моторной коры до клетки переднего рога спинного мозга 2 — Нижний мотонейрон — Начинается от клетки переднего рога и заканчивается на мышце e.грамм. все периферические нервы
2- ПОВРЕЖДЕНИЯ ДВИГАТЕЛЬНЫХ НЕЙРОНОВ
Причины поражения UMN • Повышенный тонус (спастичность) • Повышенные рефлексы • Клонус: повторяющиеся сокращения и расслабление мышц колеблющимся образом каждую секунду • Бабинский Признак: стимуляция подошвы стопы вдоль внешней границы вызывает вытягивание большого пальца стопы вверх и раздувание других пальцев стопы (обычно у взрослых эта стимуляция вызывает подошвенный рефлекс, то есть сгибание большого и малого пальцев вниз.) Примечание: в возрасте до одного года рефлекс Бабинского обычно присутствует. Зачем ?
Признак Бабинского — твердый признак поражения верхнего мотонейрона , означает повреждение латерального кортикоспинального тракта
LMN • LMN (мотонейрон Лоу) — мотонейрон от клеток переднего рога до скелетных мышц (периферических нервы). • Нижний мотонейрон также от ядер черепных нервов к скелетным мышцам лица и головы.
Поражение LMN • Поражение LMN (как повреждение последнего пути к мышце) • Снижение тонуса (гипотония / вялость).• Снижение силы мышц. • Снижение рефлексов. • Истощение мышц.
UMN LESION Паралич влияет на движение, а не на мышцы. Мышечное истощение возникает только из-за неиспользования, поэтому незначительное. Иногда отмечается при хронических тяжелых поражениях. Спастичность складного типа. Мышцы гипертонические. ПОВРЕЖДЕНИЕ LMN Поражаются отдельные мышцы или группы мышц. Истощение выражено. Вялость. Мышцы гипотонические. РАЗНИЦА МЕЖДУ ПОВРЕЖДЕНИЕМ НЕЙРОНА ВЕРХНЕГО И НИЖНЕГО ДВИГАТЕЛЯ
Поражение UMN Сухожильные рефлексы повышены.Клонус часто присутствует. Снижение или изменение поверхностных рефлексов. Абдоминальный рефлекс отсутствует. Знак Бабинского + ve, — усиление рывков челюстью. Поражение LMN Сухожильные рефлексы снижены или отсутствуют. Поверхностные рефлексы часто не изменяются.
ОБЩИЕ СЛОВА, ИСПОЛЬЗУЕМЫЕ В НЕЙРОФИЗИОЛОГИИ И КЛИНИЧЕСКОЙ НЕВРОЛОГИИ • ГЕМИПЛЕГИЯ — Паралич (потеря силы) половины тела • ГЕМИПАРЕЗ — Частичная потеря силы половины тела • ПАРАПЛЕГИЯ — Паралич обеих ног ПАРАПАРЕЗ — частичная потеря силы в обеих ногах • КВАДРИПЕГИЯ — Паралич всех четырех конечностей • МОНОПЛЕГИЯ — Паралич одной конечности
Мы увидим некоторые поражения сенсорной и двигательной системы
R L Поражение какое нарушение вызывает правая спинная колонка в L1? Щелкните для ответа. Повреждение правой спинной колонны в L1 вызывает отсутствие легкого прикосновения, вибрации и ощущения положения в правой ноге.Ниже T6 существует только fasciculus gracilis. Щелкните для объяснения
Ипсилатеральная потеря легкого прикосновения, вибрации и положения Поражение правой дорсальной колонки Щелкните, чтобы анимировать DRG R L L1 Поражение дорсальной колонны Распространенные причины включают рассеянный склероз, проникающие ранения и сдавление опухолями.
R L Какое поражение вызывает поражение правого бокового спиноталамического тракта на уровне L1? Щелкните для ответа. Повреждение правого бокового спиноталамического тракта на уровне L1 вызывает отсутствие боли и температурных ощущений в левой ноге.Нажмите для объяснения
Контралатеральная потеря боли и чувствительности к температуре Поражение правого бокового спиноталамического тракта Нажмите, чтобы оживить DRG R L L1 Боковое поражение спиноталамического тракта. К частым причинам относятся рассеянный склероз, проникающие ранения и сдавление от опухолей.
R L Какое поражение вызывает поражение правого бокового кортикоспинального тракта на уровне L1? Нажмите для ответа. Повреждение правого бокового кортикоспинального тракта на уровне L1 вызывает признаки верхних мотонейронов (слабость или паралич, гиперрефлексия и гипертонус) в правой ноге.Нажмите для объяснения
Признаки ипсилатеральных верхних мотонейронов генерализованные ниже уровня поражения Признаки UMN Слабость (спастический паралич) Гиперрефлексия (+ Babinski, clonus) Гипертония Правое боковое поражение кортикоспинального тракта UMN Нажмите, чтобы оживить RL Общий L1 Латеральный кортико-спинальный тракт Причины включают проникающие ранения, латеральную компрессию опухолями и рассеянный склероз.
Какие нарушения вызывает полное пересечение правой половины спинного мозга (синдром Гемикорда или Брауна-Секара) в L1? R L Щелкните для ответа. Повреждение правых спинных столбов в L1 приводит к отсутствию легкого прикосновения, вибрации и ощущения положения в правой ноге.Повреждение латерального кортикоспинального тракта вызывает симптомы верхнего мотонейрона в правой ноге (моноплегия), а повреждение латерального спиноталамического тракта вызывает отсутствие боли и температурных ощущений в левой ноге. Нажмите для объяснения
R L Поражение дорсального столба Ипсилатеральная потеря легкого прикосновения, вибрации и ощущения положения Боковое поражение кортикоспинального тракта Признаки ипсилатерального верхних мотонейронов Боковое поражение спиноталамического тракта Контралатеральная потеря чувства боли и температуры Поражение гемикорда Поражение гемикорда ( Синдром Брауна-Секара) Нажмите, чтобы оживить L1. Распространенные причины включают проникающие ранения, латеральное сжатие опухолями и рассеянный склероз.Постройте повреждение
Ипсилатеральная потеря легкого прикосновения, вибрации и положения Чувство Ипсилатеральные верхние двигательные нейроны Признаки Контралатеральная потеря боли и ощущения температуры Поражение гемикорды Поражение гемикорды (синдром Брауна-Секварда) UMN Нажмите, чтобы оживить DRG RL DRG L1 Поражение дорсальной колонны Поражение бокового кортикоспинального тракта Поражение бокового спиноталамического тракта
Спасибо
Нейробиология Лекция 8 — Верхние двигательные нейронные системы (пирамидные и экстрапирамидные) Карточки
-развитие пирамидной системы напрямую связано со способностью животного выполнять умелые движения
-хорошо развиты у приматов с пальцами и противоположными большими пальцами
-слабо развита у домашних животных, особенно у лошадей, быков и овец
❤NEURON 1
-клеточные тела пирамидной системы расположены в коре головного мозга в двигательной области, которая находится в предкладовой области
-соматотопически устроен
-регионы, в которых задействованы более высококвалифицированные функции, имеют большее представительство
-мышцы с небольшими двигательными единицами также имеют большую площадь представительства
— первичная моторная область одного головного мозга обслуживает мускулатуру на противоположной стороне тела
-от тела клетки аксон нейрона 1 проходит через внутреннюю капсулу в crus cerebri (все еще сохраняя соматотопическое расположение) на вентральной поверхности среднего мозга
-аксоны пирамидной системы делятся на кортикоядерные аксоны (управляющие мышцы головы) и кортикоспинальные аксоны (управляющие скелетными мышцами тела)
— кортикоядерные волокна пересекаются около ядер черепных нервов, которые иннервируют поперечно-полосатую мышцу головы (III, IV, V, VI, VII, IX), прежде чем войти в эти ядра в синапсы внутри них
-полосатая мышца головы включает экстраокулярные мышцы (CN III, IV, VI), мышцы жевания (CN V), мышцы мимики (CN VII) и язык (IX)
— оставшиеся аксоны (кортикоспинальные аксоны) продолжаются в пирамиде (соматотропное расположение) после прохождения под поперечными волокнами моста, затем перекрещиваются на уровне продолговатого мозга и пересекаются с противоположной стороной спинного мозга, образуя латеральный кортикоспинальный тракт
— несколько аксонов не перекрещиваются, а остаются ипсилатеральными в вентральном кортикоспинальном тракте перед перекрестом в спинном мозге
❤NEURON 2
— скрещенная пирамидная система аксонов формирует латеральный кортикоспинальный тракт медиальнее краниальных выступающих спиноцеребеллярных трактов в дозолатеральном канатике
— самый синапс на интернейронах в вентральной серой колонке, которые влияют на расположенные там LMN
❤NEURON 3
-Тело клетки в черепном нерве (III, IV, V, VI, VII или IX) или в сером веществе брюшного рога спинного мозга
-эта клетка является гамма-мотонейроном, но иногда может быть альфа-мотонейроном.
Центральная нервная система и викторина 2
Слайд 065-1N спинной мозг Кросс Массона Веб-микроскоп
Слайд 065-2 спинной мозг поясничный H&E cross Webscope Imagescope
Слайд 065-1 спинной мозг поясничный H&E крест Вебскоп Имископе
Слайд 066a грудной спинной мозг грудной спинной мозг luxol blue cross Webscope Imagescope
Просмотрите организацию спинного мозга, используя свой атлас.Изучите поперечный разрез поясничного отдела спинного мозга на слайде № 65-2 . При малом увеличении отличите внутреннее серое вещество от внешнего белого вещества и определите дорсальные и вентральные рога серого вещества. Вы также должны идентифицировать дорсальные и вентральные рога на слайде Slide 65-1N , окрашенном трихромом Массона. На этих слайдах дорсальные рога расположены «вверх», но вы должны уметь отличить дорсальные и вентральные рога на основании морфологии , а клетки представляют , а не ориентацию.Перикария больших соматических мотонейронов # 065-2 Вебскоп, расположенный в вентральном роге спинного мозга, иннервирует скелетные мышцы конечностей и туловища, которые эмбриологически происходят из сомитов (следовательно, соматические мышцы).
Почему перикарионы нейронов дорсального рога меньше, чем нейроны вентрального рога?
Нейроны в спинном роге — это, по сути, интернейроны, которые проецируются в другие области ЦНС (например, мотонейроны в спинном мозге или сенсорный вход в мозг), поэтому они имеют гораздо меньший общий объем и, следовательно, гораздо меньше метаболических требований по сравнению с моторными нейронами которые проецируются на мышцы, которые могут находиться на расстоянии более метра.Помните, что перикарион является центром метаболической поддержки каждого нейрона, поэтому двигательные нейроны требуют гораздо более крупных перикарионов.
Slide 66a Webscope Имисскоп показывает часть грудного отдела спинного мозга. Помимо дорсального и вентрального рогов, две структуры, особенно очевидные в грудном шнуре, — это дорсальное ядро Кларка и латеральное продолжение вентрального рога. Дорсальное ядро Clarke # 066a WebScope находится в дорсальном роге и содержит относительно крупные мультиполярные нейроны, которые получают проприоцептивную информацию от ганглиозных клеток дорсального корешка, которые иннервируются мышечными веретенами туловища и нижней конечности.Затем клетки ядра Кларка передают эту информацию через аксонные проекции, которые простираются до мозжечка (отсюда и причина того, почему клетки такие большие), где она обрабатывается для обеспечения скоординированного движения. Латеральное расширение вентрального рога # 065-2 содержит относительно большие мультиполярные висцеральные двигательные нейроны промежуточно-латерального клеточного столба, который простирается от уровней T1 до L2 спинного мозга. Клетками здесь являются преганглионарных симпатических нейронов , аксоны которых оканчиваются либо ганглиями симпатической цепи, либо «висцеральными» (или «преаортальными») ганглиями, связанными с основными ветвями брюшной аорты (например,грамм. целиакия, аортокоренальные и верхние / нижние брыжеечные ганглии). Обратите внимание, что крестцовые уровни спинного мозга (уровни S2-4) также содержат висцеральные двигательные нейроны в боковом роге, но это парасимпатических. Многие нейроны спинного мозга могут казаться сморщенными и окруженными пустым пространством из-за плохой фиксации. Клетки, которые хорошо сохранились, имеют черты, характерные для большинства нейронов: крупное тело клетки, большое бледное ядро, вещество Ниссля и отростки клетки (большинство из которых являются дендритами).Тонкая сеть дендритных отростков и нервных волокон (аксонов), лежащих между клетками серого вещества, называется нейропилем. Белое вещество содержит нервные волокна (аксоны), входящие и выходящие из серого вещества и перемещающиеся вверх и вниз по спинному мозгу, связывая его с мозгом. Нервная ткань состоит из двух основных категорий клеток: нейронов и опорных клеток (глии). И нейроны, и глия имеют тонкие отростки, выходящие из тела клетки, которые, как правило, не могут быть определены в световом микроскопе без специальных методов окрашивания.Астроциты в ЦНС обеспечивают метаболическую поддержку нейронов и играют важную роль в поддержании гематоэнцефалического барьера (см. слайд 13270 астроцитов ниже). Олигодендроциты (другой тип глиальных клеток) ответственны за миелинизацию аксонов ЦНС. Напомним, что клетки Шванна — это глиальные клетки, ответственные за миелинизацию периферической нервной системы. Миелин богат липидами и при общем осмотре выглядит белым. Таким образом, в «белом веществе» головного и спинного мозга миелинизированные аксоны являются преобладающим компонентом нейрональных клеток, а большинство ядер, которые вы видите в белом веществе, в основном состоят из глиальных клеток.«Серое вещество» содержит относительно больше нейрональных и глиальных перикариев, а также немиелинизированных (, например, дендритных) отростков. Другой важный тип глиальных клеток, о котором вам следует знать, — это микроглия, которая представляет собой небольшие клетки, полученные из моноцитов крови.