Специализированные структуры свободной клеточной поверхности-микроворсинки, реснички и жгутики, миелиновая оболочка :структурная организация и функции
Работа добавлена: 2015-12-06
Специализированные структуры свободной клеточной поверхности-микроворсинки, реснички и жгутики, миелиновая оболочка :структурная организация и функции.
Специализированные органеллы и структуры встречаются не во всех клетках.
Они характерны для зрелых клеток, являются признаками направления их дифференцировки и обеспечивают в них специфические функции. Примерами таких органелл являются микроворсинки, реснички, жгутики, миофибриллы, тонофибриллы, нейрофибриллы и некоторые другие органеллы.
Микроворсинки. Это структуры клетки, располагающиеся на ее внешней поверхности и выступающие во внеклеточное пространство. При световой микроскопии микроворсинки видны как тонкие выросты клетки.
Если их много, то они формируют апикальную каемку на свободной поверхности. Эти выпячивания значительно расширяют площадь взаимодействия клетки с внешней средой.
Ферменты, прикрепленные к гликокаликсу и находящиеся в толще билипидного слоя мембраны микроворсинок, обеспечивают всасывание и/или переваривание веществ на поверхности клеток. В этом случае расширение контактной поверхности резко увеличивает эффективность подобных процессов, например комплекс микроворсинок в столбчатых эпителиоцитах тонкой кишки. В них микроворсинки тесно соприкасаются друг с другом, обильно покрывают внешнюю поверхность клетки. Толщина микроворсинок около 100 нм, а число и длина различны. Так, длина микроворсинок у столбчатых (призматических) клеток кишечника достигает 0,6…0,8 мкм.Во многих клетках величина и размеры микроворсинок не постоянны.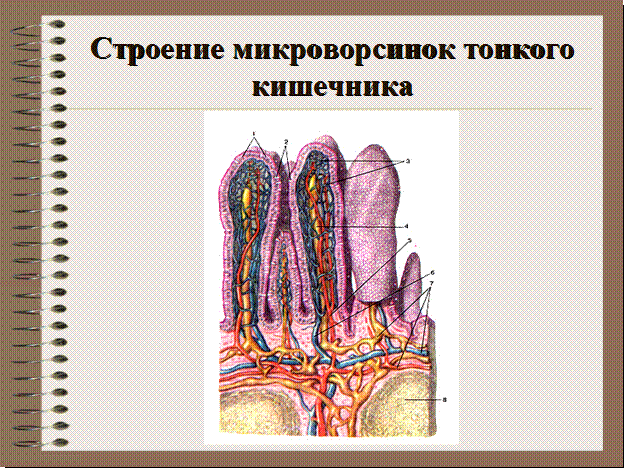 Так, в тироцигах щитовидной железы в период покоя они редкие и короткие, а при интенсивной нагрузке их высота и количество значительно увеличиваются.
Так, в тироцигах щитовидной железы в период покоя они редкие и короткие, а при интенсивной нагрузке их высота и количество значительно увеличиваются.
Микроворсинки состоят из клеточной мембраны, гиалоплазмы и тонких микрофиламентов. Актиновые (тонкие) микрофиламенты располагаются параллельно поверхности мембраны в виде компактно упакованных, упорядоченных пучков. Внутри каждой микроворсинки располагается около 20…30 актиновых нитей. Положительный полюс микрофиламентов направлен к периферии и стыкуется с электронно-плотным аморфным веществом дистальной части (верхушкой), а в основании микроворсинки актиновые микрофиламенты вплетаются в сеть подобных им структур, формирующих кутикулу.
Тонкие микрофиламенты в микроворсинке лежат параллельно друг другу на расстоянии около 10 нм, регулярно соединяясь между собой с помощью белков фимбрина и фасцина. Эти белковые комплексы образуют поперечные сцепления и объединяют тонкие микрофиламенты в компактные пучки. С мембранами тонкие микрофиламенты взаимодействуют с помощью минимиозина и виллина. Взаимодействие с минимиозинами позволяет микроворсинке сокращаться (уменьшать или увеличивать высоту).
Сходное строение с микроворсинками имеют стереоцилии. Они крупнее микроворсинок и не обладают всасывающей способностью.
Реснички и жгутики. Они представляют собой выпячивания цитоплазмы, окруженные клеточной мембраной, способные к активному движению. Органеллы хорошо заметны при большом увеличении микроскопа. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, но могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик спермия позволяет передвигаться самой клетке.
В многоклеточных организмах животных строение ресничек и жгутиков резко отличается от подобных органелл прокариот. Жгутики у бактерий образованы белком флагеллином, не имеющим отношения к комплексам микротрубочек у эукариот.
Жгутики у бактерий образованы белком флагеллином, не имеющим отношения к комплексам микротрубочек у эукариот.
Реснички у эукариот это специальные органеллы движения, встречающиеся лишь в некоторых клетках. Реснички находятся в однослойном эпителии органов дыхания и женских половых путей. В реснитчатом эпигелиоците дыхательных путей можно найти около 50…60 ресничек.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм, покрытый плазматической мембраной.
В основании ресничек и жгутика в цитоплазме видны хорошо окрашивающиеся мелкие гранулы базальные тельца. В этой области над клеткой выступает выпячивание аксонема. Аксонема («осевая нить») сложная структура, состоящая из микротрубочек и выступающая в просвет или полость органа, выстланного реснитчатым эпителием. Проксимальная часть реснички (базальное тело) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковые.
Базальное тельце по строению аналогично центриоли и состоит из девяти триплетов микротрубочек, между которыми расположены ручки, втулки и спицы. К базальному тельцу могут прикрепляться спутники, от которых отходят микротрубочки. Таким образом, базальное тельце, наряду с центросомой, является центром организации микротрубочек и может выполнять аналогичные функции.
Аксонема по составу опорного аппарата отличается от базального тельца. Стенку цилиндра аксонемы образуют 9 дуплетов микротрубочек. Кроме периферических дуплетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. Эти микротрубочки лежат отдельно на расстоянии примерно 25 нм. В целом систему микротрубочек реснички описывают как (9 х 2 + 2) в отличие от (9 х 3 + 0) системы центриолей и базальных телец.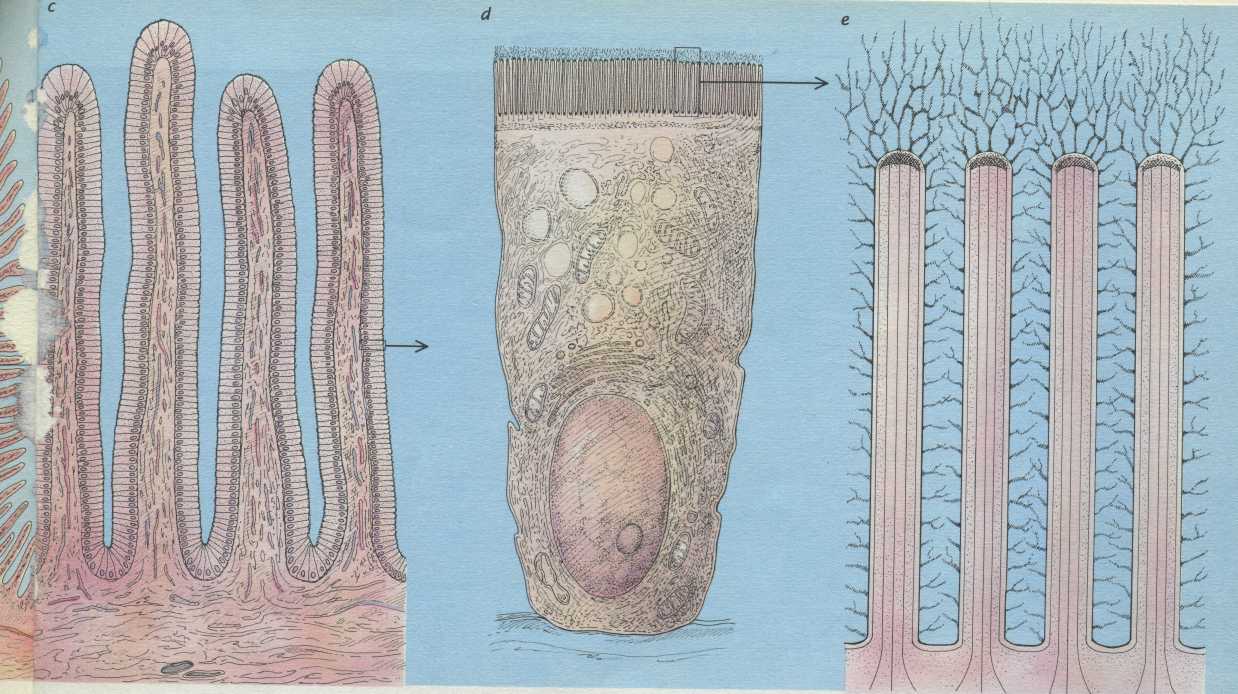
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое. Каждая из двух первых микротрубочек девяти триплетов базального тельца является основой для дуплетов микротрубочек цитоплазматического выроста, таким образом, две микротрубочки триплетов базального тельца являются микротрубочками дуплетов аксонемы. Они полимеризуются от базального тельца на основании реснички.
Из дуплета а-микротрубочка полная и образована 13 тубулинами. Неполная b-микротрубочка составлена 11 тубулинами, так как две глобулы белка являются общими с а-микротрубочками. Центральные микротрубочки формируются от центральной втулки базального тельца. Все микротрубочки достигают своим дистальным отрицательным полюсом гомогенный матрикс аксонемы.
Соседние дуплеты соединены между собой «ручками», образованными белками динеинами. Динеины обладают АТФазной активностью, способны изменять свою стереологическую структуру. В присутствии ионов кальция динеины сокращаются с потреблением энергии. Это позволяет микротрубочкам скользить относительно друг друга. Кроме динеина в составе ресничек выделяют нидоген.
К центральным микротрубочкам от периферических дуплетов радиально направляются спицы. Центральные же микротрубочки объединены втулкой.
Свободные клетки, имеющие реснички и жгутики, способны передвигаться, а неподвижные клетки движением ресничек могут перемещать жидкость и различные частицы в полых органах. При движении ресничек и жгутиков длина их не уменьшается, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна: маятникообразная, крючкообразная или волнообразная.
Основной белок ресничек тубулин не способен к сокращению, укорочению, поэтому движение ресничек осуществляется за счет активности белка динеина.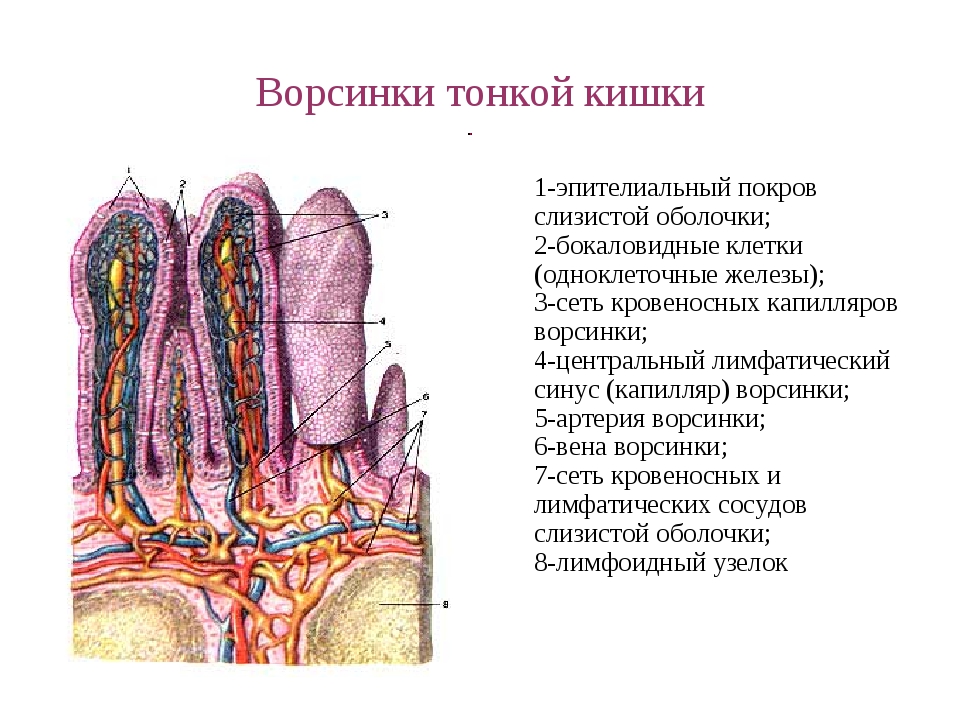 Незначительные смещения дуплетов микротрубочек относительно друг друга вызывают изгиб всей реснички, а если такое локальное смещение происходит вдоль жгутика, то возникает волнообразное движение.
Незначительные смещения дуплетов микротрубочек относительно друг друга вызывают изгиб всей реснички, а если такое локальное смещение происходит вдоль жгутика, то возникает волнообразное движение.
Волнообразное движение жгутика спермия позволяет ему передвигаться с очень высокой скоростью до 5 мм в минуту.
Волнообразное перемещение ресничек мерцательного эпителия происходит строго согласованно, но скорость движения в разных направлениях отличается. Обычно в какую-то одну сторону реснички сокращаются с большой скоростью, а в противоположном направлении их положение изменяется плавно. Это обеспечивает ток жидкости в сторону быстрого сокращения ресничек. Дефекты ресничек могут приводить к различным нарушениям, например к наследственному рецидивирующему бронхиту и хроническому синуситу, возникающим в результате нарушений функции ресничного эпителия.
Движения ресничек и жгутиков зависят от содержания внутриклеточного кальция, АТФ, ионного состава межклеточного вещества, обеспечения кислородом, глюкозой и др.
Гормоны и биологически активные вещества регулируют движения ресничек и жгутиков. Влияние того или иного гормонального фактора зависит от специализации клетки и ее рецепторного аппарата.
Реснички образуются за счет центриолей. От материнской центриоли синтезируется дочерняя центриоль, но процесс этот не заканчивается на дуплете органелл, а вновь синтезированная органелла смещается на периферию (в субмембранное пространство). Процесс может повторяться многократно. В результате под мембраной образуется множество базальных телец, которые служат основой для образования ресничек.
Базальные инвагинации (впячивания). Это внедрения цитолеммы в цитоплазму, прилежащие к базальной мембране или иной плотной структуре. Чаще всего базальные впячивания встречаются в эпителии в базальной части клетки. В других тканях они встречаются гораздо реже, например впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, так называемая гофрированная каемка. Базальные инвагинации существенно увеличивают внутреннюю (базальную) поверхность клетки.
Чаще всего базальные впячивания встречаются в эпителии в базальной части клетки. В других тканях они встречаются гораздо реже, например впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, так называемая гофрированная каемка. Базальные инвагинации существенно увеличивают внутреннюю (базальную) поверхность клетки.
Нередко в зонах инвагинации цитолеммы видны многочисленные митохондрии. Совокупность базальных инвагинаций и митохондрий формирует базальную исчерченность, которая хорошо заметна при большом увеличении микроскопа в дистальных и проксимальных канальцах нефронов почек. Базальная исчерченность в эпителиоцитах канальцев это признак активных процессов трансмембранного переноса веществ; наряду с увеличением поверхности присутствует высокий уровень энергетического потребления, обеспечиваемый АТФ.
Базальные впячивания формируют сложный лабиринт каналов и ходов, взаимных переплетений. На поверхности клеточной мембраны обнаруживают много ионных каналов, рецепторов, значительную ферментативную активность.
Форму базальных впячиваний поддерживают структуры цитоскелета: промежуточные филаменты и тонкие микрофиламенты. Они соединяются с внутренней поверхностью мембраны с помощью интегральных мембранных белков, которые, в свою очередь, сцеплены друг с другом через гликокаликс. Состав промежуточных филаментов разнообразен и зависит от тканевой принадлежности клетки: в эпителии это цитокератины, которые формируют микрофибриллы (гонофибриллы).
Миофибрилла. Это специализированная органелла мышечной ткани, основная функция которой сокращение. При световой микроскопии миофибриллы заметны в скелетной и сердечной мышечной тканях, где они имеют вид мощных пучков волокон, расположенных строго упорядоченно и придающих симпластам или клеткам продольную и поперечную исчерченность.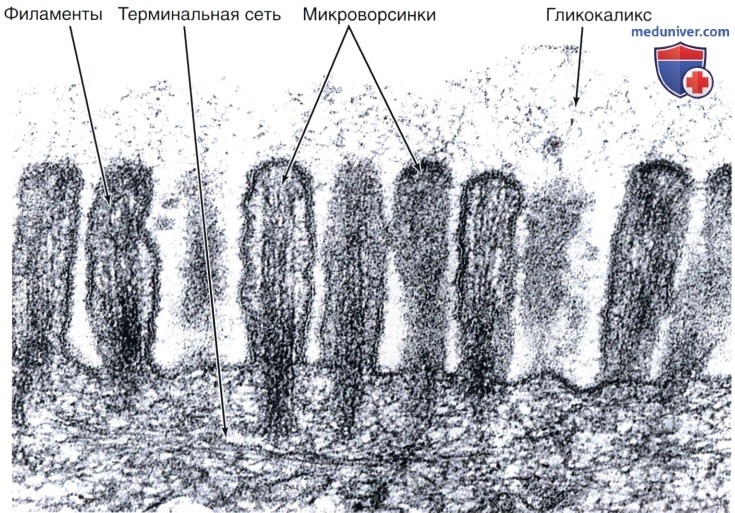
Миофибрилла представляет собой систему взаимодействующих друг с другом тонких и толстых микрофиламентов (миофиламентов). Отдельные миофиламенты можно рассмотреть лишь при электронной микроскопии, но каждая миофибрилла (особенно в поперечнополосатых мышечных тканях) состоит из сотен таких нитей. Распределение миофиламентов может быть строго упорядочено в продольном направлении и придавать структуре поперечную и продольную исчерченность либо распределяться в виде сети (в гладких мышечных клетках). В последнем случае миофиламенты при световой микроскопии выявить не удается.
Сокращение в миофибрилле обеспечивается взаимодействием белков, образующих миофиламенты. Сокращение это энергоемкий процесс, зависящий от внутриклеточного содержания ионов кальция. Сокращение может быть вызвано прямым нервным либо гуморальным влиянием. Любое из специфических возбуждений сопровождается деполяризацией мембраны с увеличением внутриклеточного содержания ионов кальция, что активизирует взаимодействие актина с миозином.
Рассмотрим строение миофибриллы на примере скелетного мышечного волокна (симпласта). В скелетном мышечном волокне миофибрилла продолжается на всю длину симпласта. Миофибриллы поперечнополосатых мышечных тканей имеют поперечную исчерченность, которая формируется при чередовании светлых (изотропных, I-дисков) и темных (анизотропных, A-дисков) дисков.
Анизотропные диски в поляризованном свете обеспечивают двойное лучепреломление, а изотропные такой способностью не обладают. Светлые диски при световой микроскопии имеют вид слабо окрашенных полос. На электронной микрофотографии видно, что светлые диски не содержат толстых нитей (миофиламентов). При очень большом увеличении светового микроскопа иногда в центре светлого I-диска заметна темная линия телофрагма, или Z-линия (зона соединения между собой тонких нитей).
Темные диски при электронной микроскопии видны как зона параллельно лежащих толстых миофиламентов. Значительная часть темного диска содержит как толстые, так и тонкие миофиламенты. При световой и электронной микроскопиях эти участки просматриваются как наиболее темные. В центре темного А-диска под очень большим увеличением светового микроскопа иногда можно увидеть мезофрагму, или М-линию область соединения толстых нитей. Она видна как тончайшая темная поперечная полоска. Участок A-диска, в котором отсутствуют тонкие нити, называется Н-зоной. В отличие от темного диска в целом Н-зона несколько светлее окрашена.
Соотношение длин анизотропного и изотропного дисков величина непостоянная и зависит от степени сокращения или расслабления. Так, в момент максимального сокращения длина изотропного диска минимальна, а Н-зона отсутствует. При расслаблении длина изотропного диска и Н-зоны максимальна. Размеры анизотропного диска при этом остаются относительно постоянными.
Толстые нити (миофиламенты) имеют поперечный диаметр около 10…12 нм, они образованы сложно устроенными белками миозинами. Каждая молекула миозина содержит две тяжелые и две легкие цепи полипептидов меромиозинов, то есть миозин это тетрамер. Он состоит из тела (хвоста), шейки и головки. Шейка и тело сформированы из двух взаимно переплетающихся полипептидных цепочек тяжелых цепей меромиозинов. Головка раздвоена и к ней присоединяются две легкие цепи меромиозина. Легкий меромиозин (миозин) способен разрушать АТФ, то есть обладает АТФазной активностью. Между шейкой и головкой миозина есть «шарнирное» соединение место, легко изменяющее свою пространственную ориентацию или изгибающееся подобно суставу. Это происходит в момент взаимодействия головки с белками тонкого миофиламенга актинами.
Тонкий миофиламент по строению близок к тонким микрофиламентам и состоит из двойной цепочки актинов. Они спирально закручены. В отличие от обычных актиновых нитей тонкие миофиламенты достаточно стабильные структуры и не подвергаются постоянному распаду и полимеризации. Эта устойчивость объясняется присоединением к актиновым цепочкам их стабилизирующего белка тропомиозина (фибриллярного белка).
Они спирально закручены. В отличие от обычных актиновых нитей тонкие миофиламенты достаточно стабильные структуры и не подвергаются постоянному распаду и полимеризации. Эта устойчивость объясняется присоединением к актиновым цепочкам их стабилизирующего белка тропомиозина (фибриллярного белка).
Кроме него в тонком миофиламенте имеются и другие белки тропонины, составляющие комплекс из трех глобул. Эти глобулы представлены С-, I- и Т-тропонинами. С-тропонин связывается с ионами кальция, I-тропонин препятствует взаимодействию актина с головкой миозина, а Т-тропонин присоединяется к тропомиозину. В покое актины связаны с тропомиозином и тропонинами так, что актин блокирован и не может взаимодействовать с миозином.
При возбуждении мышечной клетки в матриксе цитоплазмы резко увеличивается содержание ионов кальция. Они соединяются с С-тропонином, к которому имеют высокую степень сродства. Это изменяет форму тропонинового комплекса, что сопровождается перестройкой пространственной конфигурации тропомиозина и изменением формы актиновой нити в целом. В результате молекулы актина могут взаимодействовать с головками миозина. Миозиновые головки соединяются с ближайшими актинами, но при этом происходит сокращение миозинов в зонах шарнирных соединений. В результате толстая нить слегка продвигается вперед в направлении центральной части изотропного диска.
Следующим шагом является разрушение АТФ легкими цепями меромиозина. Этой энергии хватает, чтобы разорвать связь миозина с актином. Шарнирное соединение «выпрямляется», то есть занимает исходное положение, но оказавшаяся чуть впереди головка вновь связывается с последующими молекулами актина. Вновь происходит сокращение и движение вперед. Таким образом, миозиновые головки как бы «шагают» по актиновым нитям за счет шарнирного соединения и АТФазной активности миозина.
При прекращении возбуждения в мышечном волокне содержание ионов кальция вновь снижается, С-тропонин высвобождается, это приводит к тому, что актиновый (тонкий) миофиламент вновь восстанавливает свою прежнюю структуру, и актин «закрывается» тропомиозином. В этой ситуации взаимодействие миозина с актином становится вновь невозможным, и миофибрилла занимает исходное положение происходит расслабление мышечного волокна.
Распределение миофибрилл в скелетной мышце отличается от сердечной. В кардиомиоцитах миофибриллы занимают в основном периферию клетки, тогда как в скелетном симпласте они располагаются центрально. В гладком миоците толстых миофиламентов фактически нет, и тонкие миофиламенты взаимодействуют с молекулами минимиозина.
Возможно эти работы будут Вам интересны.
1. Структурная и функциональная организация рибосомы
2. Многомашинные и многопроцессорные вычислительные комплексы. Определение, типы связей и структурная организация. Особенности программного обеспечения
3. Психоаналитическая концепция Фрейда. Структурная гипотеза Фрейда. Функции механизма, нейтрализующий конфликт между психическими инстанциями
4. Главная цепь дыхательных ферментов в митохондриях, ее структурная организация и биологическая роль. Цитохромы, цитохромоксидаза, химическая природа и роль в окислительных процессах
5. Организация, структура и функции web-сервера
6. Клеточная теория, вклад Р. Вихрова и его критиков в развитие клеточной теории
7. Химичекий состав и строение структуры хромосомы. Динамика её структуры в клеточном цикле
8. Специализированные агентства по связям с общественностью
9. Специализированные (нештатные) формирования госсанэпидслужбы России
10. Управляющее меню FAR. Программа оболочка Far
Строение жгутиков и ресничек — От Земли до Неба
Микроворсинки — структуры клетки, располагающиеся на ее свободной поверхности и выступающие во внеклеточное пространство,
Эти выпячивания значительно расширяют поверхность клетки.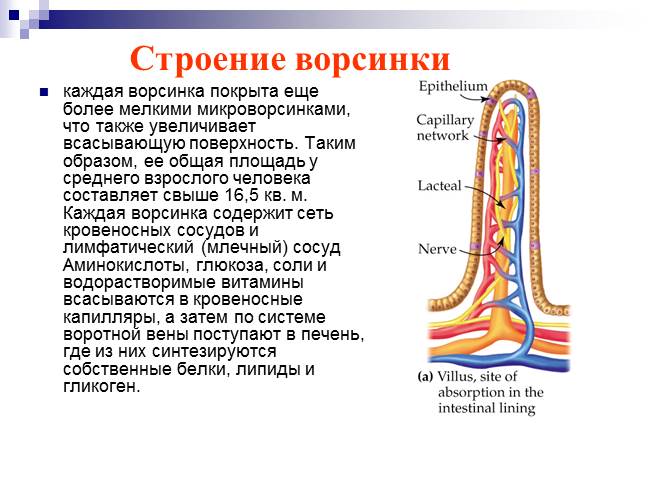 Микроворсинки активно всасывают и (или) активно переваривают вещества на поверхности клеток. Микроворсинкам присущи некоторым популяциям эпителиоцитов. Нередко для клеток, имеющих много микроворсинок, характерна высокая ферментативная активность и счет белков, прикрепляющихся к гликокаликсу (столбчатые энтероциты кишечника, тироциты щитовидной железы и т. д.).
Микроворсинки активно всасывают и (или) активно переваривают вещества на поверхности клеток. Микроворсинкам присущи некоторым популяциям эпителиоцитов. Нередко для клеток, имеющих много микроворсинок, характерна высокая ферментативная активность и счет белков, прикрепляющихся к гликокаликсу (столбчатые энтероциты кишечника, тироциты щитовидной железы и т. д.).
Микроворсинки сформированы клеточной мембраной к гиалоплазмой.
Структурной основой каркаса микроворсинок служат тонкие микрофиламенты. (+)-конец микрофиламентов направлен на периферию и стыкуется с электронноплотным аморфным веществом дистальной части микроворсинки. Тонкие микрофиламенты в микроворсинке лежат параллельно друг к другу на расстоянии около 10 нм, регулярно соединяясь между собой с помощью белков — фимбрина и фаоцима. С мембранами они взаимодействуют с помощью минимиозина и фасцина. На поверхности микроворсинок нередко выявляется высокая концентрация белков с транспортно-насосными функциями (всасывающие эпителии).
Сходное строение имеют стереоциллии, однако в отличие от микроворсинок, стереоцилии имеют гораздо большие размеры и не обладают всасывающими функциями.
Реснички и жгутики. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, ко могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик сперматозоида позволяет передвигаться самой клетке.
Реснички состоят из базалъного тельца и выступающей из клетки аксонемы. Базальное тельце по строению аналогично дочерней центриоле и состоит из девяти триплетов микротрубочек.
В основе аксонемы тоже лежат микротрубочки. Они формируются от базального тельца и достигают своим дистальным (-) концом гомогенного матрикса. Имеется девять дуплетов микротрубочек и две центральные микротрубочки. Каждый периферический дуплет представлен: а- микротрубочкой (полной) и б — микротрубочкой (неполной). Соседние дуплеты соединены между собой линейном (обладающим АТФ-азной активностью и позволяющим микро-трубочкам скользить по отношению друг к другу) и индогеном.
Соседние дуплеты соединены между собой линейном (обладающим АТФ-азной активностью и позволяющим микро-трубочкам скользить по отношению друг к другу) и индогеном.
К центральным микротрубочкам от периферических радиально направляются спицы. Центральные же микротрубочки объединены втулкой.
Базальные инвагинации (впячивания).
Базальные впячивания – это внедрения цитолеммы в цитоплазму, прилежащие к базальной мембране или иной плотной структуре, например, впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, существенно расширяющие поверхность клетки с внутренней (базальной) поверхности. Нередко в зонах впячиваний цитолеммы видны многочисленные мнтохондрии. Совокупность базальных инвагинаций и митохондрий формирует базальную исчерченность.
Наличие базальной исчерченности — признак активных процессов трансмембранного перекоса веществ, когда наряду с увеличением поверхности значение имеет и высокий уровень энергетического потребления, обеспечиваемый АТФ.
Базальные впячивания формируют сложный лабиринт каналов и ходов, взаимопереплетений. На их поверхности обнаруживаете высокая концентрация ионных каналов, рецепторов, нередко — ферментативная активность. Форму базальных впячиваний поддерживают структуры цитоскелета — в первую очередь промежуточные филаменты и тонкие микрофиламенты. Состав промежуточных филаментов разнообразен и зависит от тканевой принадлежности клетки. В эпителиях — это цитокератины. Они формируют микрофибриллы. В зоне впячиваний часто можно встретить полудесмосомальные контакты.
86
Источник: StudFiles.net
Вращающим элементом служит базальное тельце, движущееся внутри бактериальной мембраны, подобно ротору в электромоторе. Интересно, что и источник энергии для вращения жгутика не энергия химических веществ (типа АТФ), а разность электрохимических потенциалов на мембране бактериальной клетки.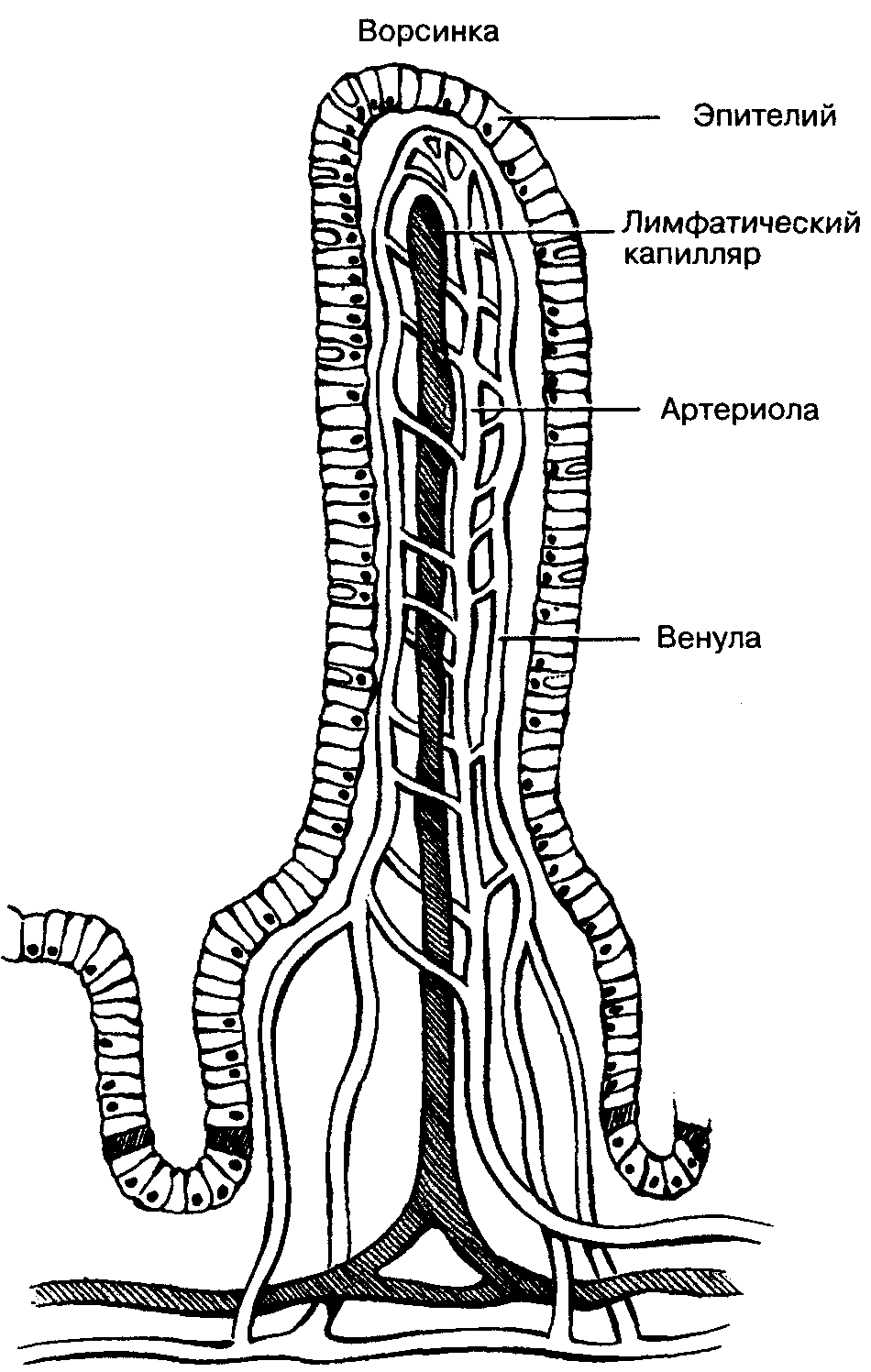
Жгутики и реснички животных и растений гораздо крупнее. Они имеют диаметр около 250 нм и достигают в длину нескольких миллиметров. В отличие от бактериальных эти жгутики покрыты мембраной и обладают собственной подвижностью.
Строение и принцип работы жгутиков и ресничек совершенно одинаковы. Различия между ними лишь в количестве: обычно на одну клетку приходится один или несколько жгутиков, а ресничек до нескольких тысяч.
Под мембраной у жгутиков (ресничек) располагается стержневая структура — аксонема. Она состоит из 9 спаренных микротрубочек, расположенных по окружности, и 2 одиночных микротрубочек в центре. Центральные и периферические микротрубочки соединены между собой системой связок. В состав периферических микротрубочек входит белок тубулин и специфический для жгутиков белок — динеин. Динеин использует энергию АТФ (см. Аденозинтрифосфорная кислота (АТФ), расщепляя ее до АДФ (аденозиндифосфорной кислоты), и, взаимодействуя с тубулином, превращает энергию в механическую работу по перемещению спаренных микротрубочек относительно соседних — скольжению.
iv>
е центральные микротрубочки и система связок превращают скольжение отдельных микротрубочек внутри аксонемы в изгибание всей ак-сонемы, а это и приводит в движение жгутик (ресничку). Движение состоит либо из псевдовращения (такое движение мы производим, например, делая вращательные движения рукой) — так чаще всего бьются жгутики; либо из возвратно-поступательных колебаний в одной плоскости, типичных для ресничек.
В основании жгутиков (ресничек) лежат базальные тельца, но в отличие от бактериальных они не связаны с движением, а служат для роста аксонемы и закрепления жгутика (реснички) в клетке. Часто от базальных телец в глубь цитоплазмы отходят дополнительные заякоривающие структуры — исчерченные корешки.
Жгутики и реснички обеспечивают движение свободноживущих клеток. Реснички создают поток жидкости вдоль поверхности неподвижных клеток (например, в дыхательных путях согласованное биение миллионов ресничек обеспечивает удаление из легких пылевых частиц).
У некоторых животных (например, млекопитающих) имеются наряду с подвижными ресничками еще и неподвижные — чувствительные реснички. Последние не имеют двух центральных микротрубочек и иногда вообще лишены аксонемы, но всегда имеют в основании базальное тельце. Таковы палочки сетчатки глаза, реснички обонятельных луковиц, реснички во внутреннем ухе (орган равновесия).
Источник: alnam.ru
- ПРЕДИСЛОВИЕ РЕДАКТОРОВ ПЕРЕВОДА
- ПРЕДИСЛОВИЕ
- Благодарности
- ЧАСТЬ I. БИОМОЛЕКУЛЫ
ГЛАВА 1. БИОХИМИЯ-МОЛЕКУЛЯРНАЯ ЛОГИКА ЖИВЫХ ОРГАНИЗМОВ - 1.1. Для живой материи характерны некоторые отличительные особенности
- 1.2. Биохимия стремится понять природу живого состояния
- 1.3. Все живые организмы содержат органические макромолекулы, построенные по общему плану
- 1.4. Обмен веществ и энергии в живых организмах
- 1.5. Ферменты, играющие роль катализаторов в живых клетках, управляют сложно организованной сетью химических реакций
- 1.6. Клетки используют энергию в химической форме
- 1.7. Процессы клеточного метаболизма находятся под постоянным контролем
- 1.8. Живые организмы способны к точному самовоспроизведению
ГЛАВА 2. КЛЕТКИ - 2.1. Все клетки обладают некоторыми общими структурными характеристиками
- 2.2. Клетки должны иметь очень малые размеры
- 2.3. Существуют два больших класса клеток — прокариотические и эукариотические
- 2.4. Прокариоты — самые простые и самые мелкие клетки
- 2.5. Escherichia coli — самая известная из прокариотических клеток
- 2.6. Эукариотические клетки крупнее и сложнее прокариотических
- 2.7. Ядро эукариот — это очень сложная структура
- 2.8. Митохондрии — «силовые установки» эукариотических клеток, поставляющие энергию
- 2.9. Эндоплазматический ретикулум образует каналы в цитоплазме
> - 2.
 10. Тельца Гольджи — секреторные органеллы
10. Тельца Гольджи — секреторные органеллы - 2.11. Лизосомы — контейнеры с гидролитическими ферментами
- 2.12. Пероксисомы — пузырьки, разрушающие перекись водорода
- 2.13. Микрофиламенты участвуют в сократительных процессах клеток
- 2.14. Микротрубочки также связаны с клеточными движениями
- 2.15. Микрофиламенты, микротрубочки и микротрабекулярная сеть образуют цитоскелет
- 2.16. Реснички и жгутики позволяют клеткам передвигаться
- 2.17. В цитоплазме содержатся также гранулярные тельца
- 2.18. Цитозоль — непрерывная водная фаза цитоплазмы
- 2.19. Клеточная мембрана имеет большую площадь поверхности
- 2.20. На поверхности многих животных клеток имеются также «антенны»
- 2.21. Эукариотические клетки растений имеют некоторые специфические особенности
- 2.22. Вирусы — надмолекулярные паразиты
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 3. СОСТАВ ЖИВОЙ МАТЕРИИ: БИОМОЛЕКУЛЫ - 3.1. Химический состав живой материи отличается от химического состава земной коры
- 3.2. Большинство биомолекул содержит углерод
- 3.3. Биомолекулы имеют специфическую форму и определенные размеры
- 3.4. Функциональные группы органических биомолекул определяют их химические свойства
- 3.5. Многие биомолекулы асимметричны
- 3.6. Основные классы биомолекул в клетках представлены очень крупными молекулами
- 3.7. Макромолекулы образуются из небольших молекул, играющих роль строительных блоков
- 3.8. Молекулы, используемые в качестве строительных блоков, имеют простую структуру
- 3.9. Структурная иерархия в молекулярной организации клеток
- 3.10. Биомолекулы первыми возникли в процессе химической эволюции
- 3.11. Химическую эволюцию можно воспроизвести в лабораторных условиях
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 4. ВОДА - 4.1. Необычные физические свойства воды обусловлены ее способностью участвовать в образовании водородных связей
- 4.2. Водородные связи широко распространены в биологических системах и играют в них важную роль
- 4.3. Вода как растворитель обладает необычными свойствами
- 4.4. Растворенные вещества изменяют свойства воды
- 4.5. Состояние равновесия обратимых реакций характеризуется константой равновесия
- 4.6. Ионизацию воды можно охарактеризовать величиной константы равновесия
- 4.7. Шкала pH: обозначения концентраций ионов Н+ и ОН-
- 4.8. Свойства кислот и оснований тесно связаны со свойствами воды
- 4.9. Слабые кислоты имеют характерные кривые титрования
- 4.10. Буферы — это смеси слабых кислот и сопряженных с ними оснований
- 4.11. Фосфат и бикарбонат — важные биологические буферные системы
- 4.12. Приспособленность живых организмов к водной среде
- 4.13. «Кислые» дожди загрязняют наши озера и реки
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
- ГЛАВА 5. АМИНОКИСЛОТЫ И ПЕПТИДЫ
- 5.1. Общие структурные свойства аминокислот
- 5.2. Почти все аминокислоты содержат асимметрический атом углерода
- 5.3. Стереоизомеры обозначаются в соответствии с их абсолютной конфигурацией
- 5.4. Оптически активные аминокислоты в белках представляют собой L-стереоизомеры
- 5.5. Классификация аминокислот на основе их R-групп
- 5.6. Восемь аминокислот содержат неполярные R-группы
- 5.7. Семь аминокислот содержат незаряженные полярные R-группы
- 5.8. Две аминокислоты содержат отрицательно заряженные (кислые) R-группы
- 5.9. Три аминокислоты содержат положительно заряженные (основные) R-группы
- 5.10. В некоторых белках присутствуют нестандартные аминокислоты
- 5. 11. В водных растворах аминокислоты ионизированы
- 5.12. Аминокислоты могут вести себя и как кислоты, и как основания
- 5.13. Аминокислоты имеют характерные кривые титрования
- 5.14. По кривой титрования можно предсказать, какой электрический заряд несет данная аминокислота
- 5.15. Аминокислоты различаются по своим кислотно-основным свойствам
- 5.16. Кислотно-основные свойства аминокислот служат основой для аминокислотного анализа
- 5.17. Электрофорез на бумаге позволяет разделять аминокислоты в соответствии с их электрическим зарядом
- 5.18. Ионообменная хроматография служит более эффективным способом разделения аминокислот
- 5.19. Химические реакции, характерные для аминокислот
- 5.20. Пептиды — это цепочки аминокислот
- 5.21. Разделение пептидов может быть основано на различиях в их ионизационных свойствах
- 5.22. Химические реакции, характерные для пептидов
- 5.23. Некоторые пептиды обладают высокой биологической активностью
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
- ГЛАВА 6. БЕЛКИ: КОВАЛЕНТНАЯ СТРУКТУРА И БИОЛОГИЧЕСКИЕ ФУНКЦИИ
- 6.1. Белки обладают множеством различных биологических функций
- 6.2. Белки можно классифицировать также по форме их молекул
- 6.3. В ходе гидролиза брлки распадаются на аминокислоты
- 6.4. Некоторые белки имеют в своем составе не только аминокислоты, но и другие химические группы
- 6.5. Белки — это очень крупные молекулы
- 6.6. Белки можно выделить и подвергнуть очистке
- 6.7. Определение аминокислотной последовательности полипептидных цепей
- 6.8. Инсулин — это первый белок, для которого была установлена аминокислотная последовательность
- 6.9. В настоящее время известны последовательности многих других белков
- 6.10. Гомологичные белки разных видов имеют гомологичные последовательности
- 6. 11. Различия между гомологичными белками можно выявить по иммунной реакции
- 6.12. Белки претерпевают структурные изменения, называемые денатурацией
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 7. ФИБРИЛЛЯРНЫЕ БЕЛКИ - 7.1. Термины «конфигурация» и «конформация» имеют разный смысл
- 7.2. Как это ни парадоксально, нативные белки имеют только одну или всего лишь несколько конформаций
- 7.3. a-Кератины — фибриллярные белки, синтезируемые клетками эпидермиса
- 7.4. Рентгеноструктурный анализ показывает, что в кератинах имеются повторяющиеся структурные единицы
- 7.5. Рентгеноструктурные исследования пептидов свидетельствуют о жесткости и плоской конфигурации пептидных групп
- 7.6. В a-кератине полипептидные цепи имеют форму a-спирали
- 7.7. Некоторые аминокислотные остатки препятствуют образованию a-спирали
- 7.8. В a-кератинах содержится много аминокислот, способствующих образованию a-спиральной структуры
- 7.9. В нативных a-кератинах a-спиральные полипептидные цепи скручены наподобие каната
- 7.10. a-Кератины нерастворимы в воде из-за преобладания в их составе аминокислот с неполярными R-группами
- 7.11. b-Кератины имеют другую конформацию полипептидной цепи, называемую b-структурой
- 7.12. Перманентная завивка волос — пример биохимической технологии
- 7.13. Коллаген и эластин — главные фибриллярные белки соединительных тканей
- 7.14. Коллаген — самый распространенный белок у высших животных
- 7.15. Коллаген обладает как обычными, так и необычными свойствами
- 7.16. Полипептиды в коллагене представляют собой трехцепочечные спиральные структуры
- 7.17. Структура эластина придает особые свойства эластической ткани
- 7.18. Что говорят нам фибриллярные белки о структуре белков?
- 7.19. Другие типы фибриллярных или нитевидных белков встречающихся в клетках
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 8. ГЛОБУЛЯРНЫЕ БЕЛКИ: СТРУКТУРА И ФУНКЦИЯ ГЕМОГЛОБИНА - 8.1. Полипептидные цепи глобулярных белков свернуты в плотную компактную структуру
- 8.2. Рентгеноструктурный анализ миоглобина — выдающееся достижение в исследовании белков
- 8.3. Миоглобины, выделенные из разных видов, имеют сходную конформацию
- 8.4. Глобулярные белки различных типов имеют неодинаковую структуру
- 8.5. Аминокислотная последовательность белка определяет его третичную структуру
- 8.6. Силы, стабилизирующие третичную структуру глобулярных белков
- 8.7. Свертывание полипептидных цепей происходит с очень высокой скоростью
- 8.8. Олигомерные белки имеют как третичную, так и четвертичную структуру
- 8.9. Метод рентгеноструктурного анализа позволил установить как третичную, так и четвертичную структуру гемоглобина
- 8.10. По своей третичной структуре a- и b-цепи гемоглобина очень сходны с миоглобином
- 8.11. Была установлена четвертичная структура и некоторых других олигомерных белков
- 8.12. Эритроциты — специализированные клетки, переносящие кислород
- 8.13. Для миоглобина и гемоглобина характерны разные кривые связывания кислорода
- 8.14. Кооперативное связывание кислорода делает гемоглобин более эффективным переносчиком кислорода
- 8.15. Гемоглобин служит также переносчиком С02 и ионов H+
- 8.16. Оксигенация гемоглобина вызывает изменение его пространственной конформации
- 8.17. Серповидноклеточная анемия — «молекулярная болезнь» гемоглобина
- 8.18. Гемоглобин больных серповидноклеточной анемиеи имеет измененную аминокислотную последовательность
- 8.19. Серповидная форма эритроцитов обусловлена склонностью молекул гемоглобина S к агрегации
- 8.20. «Неправильные» аминокислоты появляются в белках в результате генных мутаций
- 8.21. Можно ли найти «молекулярное лекарство» для серновидноклеточного гемоглобина?
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
- ГЛАВА 9. ФЕРМЕНТЫ
- 9.1. История биохимии — это в значительной мере история исследования ферментов
- 9.2. Ферменты обнаруживают все свойства белков
- 9.3. Ферменты классифицируются на основе реакций, которые они катализируют
- 9.4. Ферменты ускоряют химические реакции, снижая энергию активации
- 9.5. Концентрация субстрата оказывает огромное влияние на скорость реакций, катализируемых ферментами
- 9.6. Существует количественная связь между концентрацией субстрата и скоростью ферментативной реакции
- 9.7. Каждый фермент имеет характерную величину Км для данного субстрата
- 9.8. Многие ферменты катализируют реакции с участием двух субстратов
- 9.9. Каждый фермент имеет определенный оптимум pH
- 9.10. Количество фермента можно определить по его активности
- 9.11. Ферменты проявляют специфичность по отношению к своим субстратам
- 9.12. Ферменты можно ингибировать определенными химическими соединениями
- 9.13. Существуют обратимые ингибиторы двух типов — конкурентные и неконкурентные
- 9.14. Неконкурентное ингибирование тоже обратимо, но не может быть ослаблено или устранено повышением концентрации субстрата
- 9.15. Факторы, определяющие каталитическую эффективность ферментов
- 9.16. Рентгеноструктурный анализ выявил важные структурные особенности ферментов
- 9.17. В ферментных системах есть «дирижер», роль которого выполняет регуляторный фермент
- 9.18. Аллостерические ферменты регулируются путем нековалентного присоединения к ним молекул модуляторов
- 9.19. Аллостерические ферменты ингибируются или активируются их модуляторами
- 9.20. Поведение аллостерических ферментов не описывается уравнением Михаэлиса — Ментеи
- 9.21. Субъединицы аллостерических ферментов сообщаются между собой
- 9.22. Некоторые ферменты регулируются путем обратной ковалентной модификации
- 9. 23. Многие ферменты существуют в нескольких формах
- 9.24. Нарушение каталитической активности ферментов может быть обусловлено мутациями
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 10. ВИТАМИНЫ И МИКРОЭЛЕМЕНТЫ: ИХ РОЛЬ В ФУНКЦИОНИРОВАНИИ ФЕРМЕНТОВ - 10.1. Витамины — незаменимые органические микрокомпоненты пищи
- 10.2. Витамины являются важными компонентами коферментов и простетических групп ферментов
- 10.3. Витамины можно разделить на два класса
- 10.4. Тиамин (витамин B1) функционирует в форме тиаминпирофосфата
- 10.5. Рибофлавин (витамин В2) — компонент флавиновых нуклеотидов
- 10.6. Никотинамид — это активная группа коферментов NAD и NADP
- 10.7. Пантотеновая кислота — компонент кофермента А
- 10.8. Пиридоксин (витамин В6) играет важную роль в метаболизме аминокислот
- 10.9. Биотин является активным компонентом биоцитина — простетической группы некоторых ферментов, катализирующих реакции карбоксилирования
- 10.10. Фолиевая кислота служит предшественником кофермента тетрагидрофолиевой кислоты
- 10.11. Витамин B12 — предшественник кофермента В12
- 10.12. Биохимическая функция витамина С (аскорбиновой кислоты) не известна
- 10.13. Жирорастворимые витамины представляют собой производные изопрена
- 10.14. Витамин А, вероятно, выполняет несколько функций
- 10.15. Витамин D — предшественник гормона
- 10.16. Витамин Е защищает клеточные мембраны от кислорода
- 10.17. Витамин К-компонент карбокснлнрукнцего фермента
- 10.18. В пище животных должны содержаться многочисленные неорганические вещества
- 10.19. Для действия многих ферментов требуется железо
- 10.20. В некоторых окислительных ферментах содержится также медь
- 10.21. Для действия многих ферментов необходим цинк
- 10. 22. Некоторым ферментам требуются ионы марганца
- 10.23. В состав витамина В12 входит кобальт
- 10.24. Селен является и незаменимым микроэлементом, и ядом
- 10.25. Для некоторых ферментов требуются другие микроэлементы
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и залами
ГЛАВА 11. УГЛЕВОДЫ: СТРОЕНИЕ И БИОЛОГИЧЕСКИЕ ФУНКЦИИ - 11.1. Углеводы делятся на три класса в зависимости от числа остатков сахаров
- 11.2. Существует два семейства моносахаридов: альдозы и кетозы
- 11.3. Моносахариды обычно содержат несколько асимметрических центров
- 11.4. Типичные моносахариды имеют циклическую структуру
- 11.5. Простые моносахариды могут служить восстановителями
- 11.6. Дисахариды содержат две моносахаридные единицы
- 11.7. Полисахариды содержат большое число моносахаридных остатков
- 11.8. Некоторые полисахариды представляют собой форму запасания «клеточного топлива»
- 11.9. Целлюлоза — наиболее распространенный структурный полисахарид
- 11.10. Клеточные стенки содержат в больших количествах структурные и защитные полисахариды
- 11.11. Гликопротеины — гибридные молекулы
- 11.12. На поверхности клеток животных присутствуют гликопротеины
- 11.13. Гликозаминогликаны и протеогликаны — важные компоненты соединительной ткани
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 12. ЛИПИДЫ И МЕМБРАНЫ - 12.1. Жирные кислоты — структурные компоненты большинства липидов
- 12.2. Триацилглицеролы — это глицероловые эфиры жирных кислот
- 12.3. Триацилглицеролы — форма запасания липидов
- 12.4. Воска — эфиры жирных кислот и длинноцепочечных спиртов
- 12.5. Фосфолипиды — основные липидные компоненты мембран
- 12.6. Сфинголипиды — также важные компоненты мембран
- 12.7. Стероиды — неомыляемые липиды, обладающие специфическими функциями
- 12.8. Липопротеины сочетают свойства липидов и белков
- 12.9. Полярные липиды образуют мицеллы, монослои и бислои
- 12.10. Полярные липиды и белки — основные компоненты мембран
- 12.11. Мембраны имеют жидкостно-мозаичную структуру
- 12.12. Мембраны асимметричны, т.е. имеют неравноценные стороны
- 12.13. Мембраны эритроцитов исследованы очень подробно
- 12.14. Лектины — специфические белки, способные связываться с определенными клетками и вызывать их агглютинацию
- 12.15. Мембраны имеют очень сложные функции
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
- ПРИЛОЖЕНИЕ ОТВЕТЫ
 10. Тельца Гольджи — секреторные органеллы
10. Тельца Гольджи — секреторные органеллы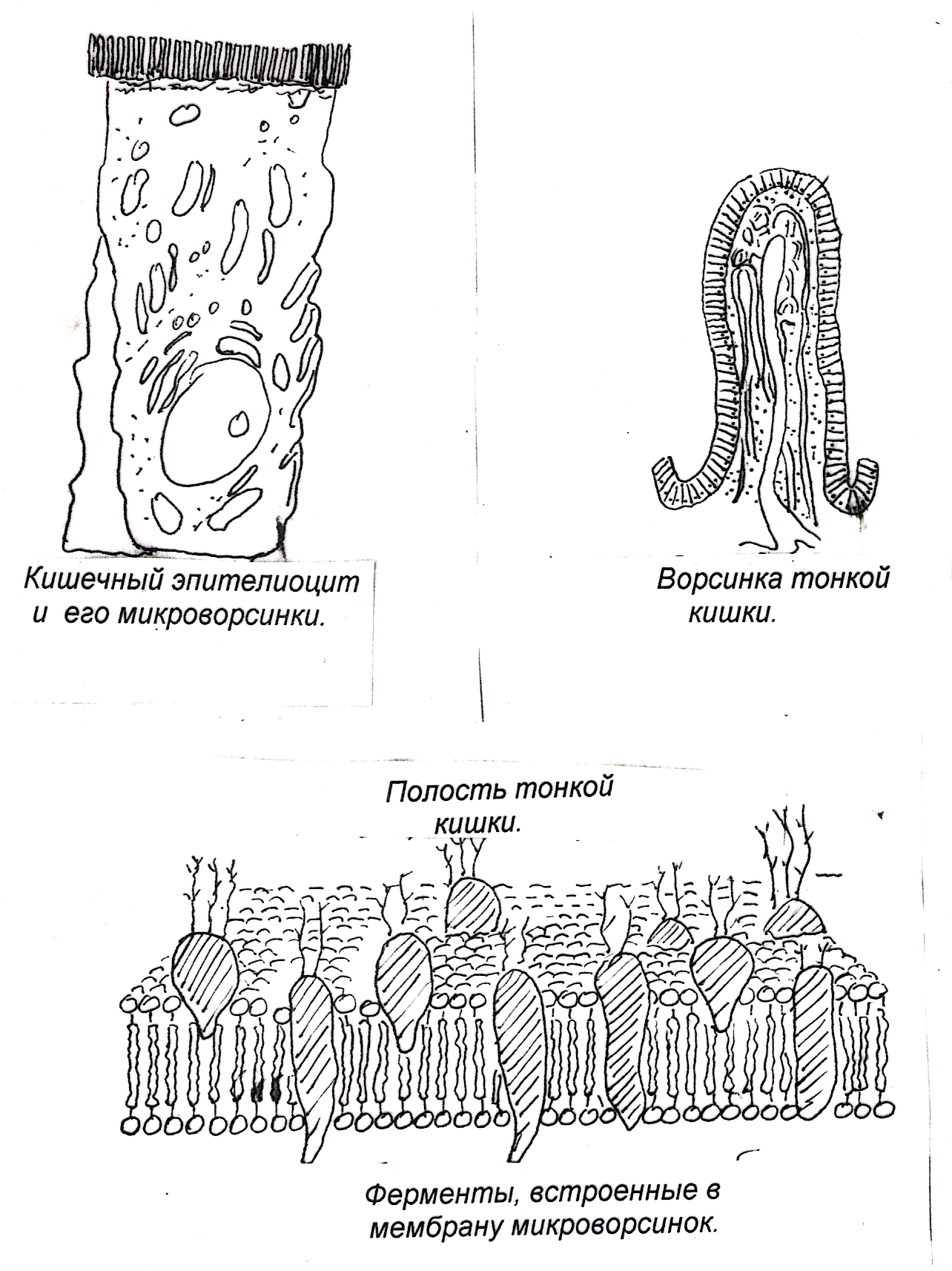 ВОДА
ВОДА 11. В водных растворах аминокислоты ионизированы
11. В водных растворах аминокислоты ионизированы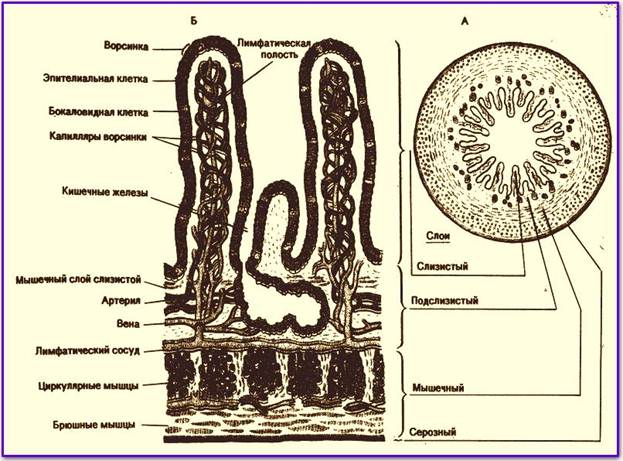 11. Различия между гомологичными белками можно выявить по иммунной реакции
11. Различия между гомологичными белками можно выявить по иммунной реакции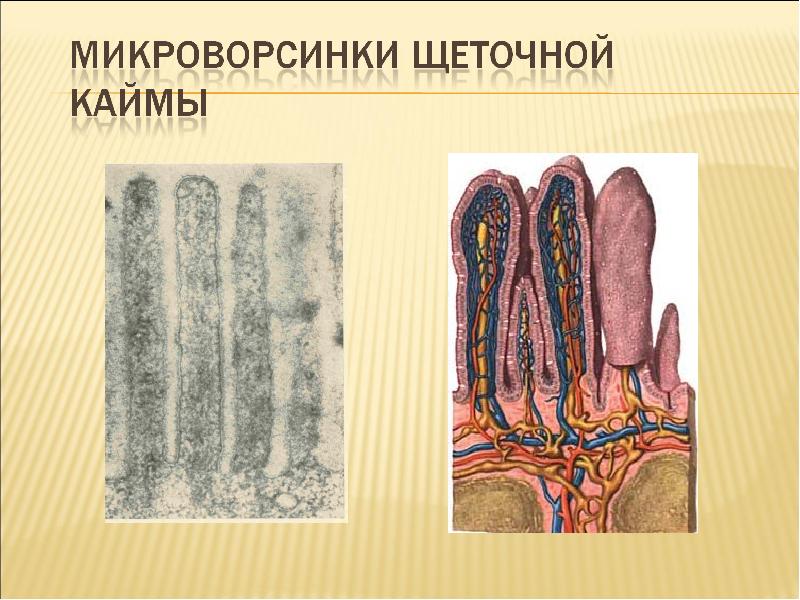 ГЛОБУЛЯРНЫЕ БЕЛКИ: СТРУКТУРА И ФУНКЦИЯ ГЕМОГЛОБИНА
ГЛОБУЛЯРНЫЕ БЕЛКИ: СТРУКТУРА И ФУНКЦИЯ ГЕМОГЛОБИНА ФЕРМЕНТЫ
ФЕРМЕНТЫ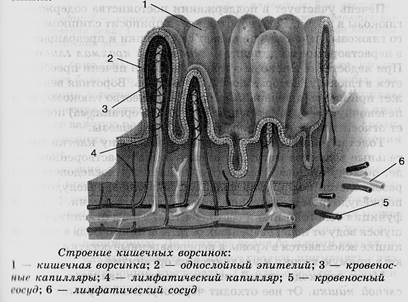 23. Многие ферменты существуют в нескольких формах
23. Многие ферменты существуют в нескольких формах 22. Некоторым ферментам требуются ионы марганца
22. Некоторым ферментам требуются ионы марганцаИсточник: scask.ru
Программа по гистологии, цитологии и эмбриологии
Программа по гистологии, цитологии и эмбриологии
Цели и задачи дисциплины: Обеспечение обучающемуся необходимой информации для овладения определенными знаниями в области общей и частной гистологии, цитологии и эмбриологии с учетом дальнейшего обучения и профессиональной деятельности по специальностям «Лечебное дело», «Педиатрия» и «Медико-профилактическое дело».
Общая гистология и цитология
1.Введение.
1.1.Назначение, содержание, место гистологии, цитологии и эмбриологии в системе подготовки врача. Возникновение и развитие гистологии, цитологии и эмбриологии как самостоятельных наук. Роль отечественных ученых в cоздание самостоятельных кафедр гистологии в России в XIX в. Развитие гистологии, цитологии и эмбриологии в XX в. Современный этап в развитии гистологии, цитологии и эмбриологии.
1.2. Методы изготовления препаратов для световой микроскопии. Сущность и методы фиксации микрообъектов. Способы уплотнения (заливки). Микротомия с использованием салазочных, ротационных микротомов. Метод замораживания. Сущность и методы окраски микропрепаратов и их заключения в бальзам, смолы, желатин. Виды микропрепаратов — срезы, мазки, отпечатки, пленки. Техника микроскопирования в световых микроскопах. Особенности микроскопии в ультрафиолетовых лучах, люминесцентная микроскопия, фазовоконтрастная микроскопия, интерференционная микроскопия, лазерная конфокальная микроскопия.
1.3.Электронная микроскопия (трансмиссионная и сканирующая), методы изготовления микрообъектов для электронной микроскопии. Специальные методы изучения микрообъектов — гистохимия (в том числе электронная гистохимия), радиоавтография, иммуногистохимия, фракционирование клеточного содержимого с помощью ультрацентрифугирования. Методы исследования живых клеток — культуры тканей вне- и внутри организма, клонирование, образование гетерокарионов и гибридов клеток, прижизненная окраска.
1.4. Количественные методы исследования: цитофотометрия, электронная микрофотометрия, спектрофлуорометрия, денситометрия.
1.5. Методы исследования в эмбриологии — особенности фиксации и приготовления тотальных препаратов и срезов органов эмбриона. Серийные срезы и пластическая реконструкция эмбриологических объектов. Методы определения возраста эмбриона человека.
2.Цитология (Клеточная биология)
2.1. Предмет и задачи цитологии, ее значение в системе биологических и медицинских наук. Основные положения клеточной теории на современном этапе развития науки. Понятие о клетке, как основной единице живого. Общий план строения клеток эукариот:
клеточная оболочка, цитоплазма, ядро. Неклеточные структуры как производные клеток. Взаимосвязь формы и размеров клеток с их функциональной специализацией.
2.2. Строение клетки
2.2.1.Биологическая мембрана как основа строения клетки. Строение, основные свойства и функции. Понятие о компартментализации клетки и ее функциональное значение.
2.2.2.Клеточная оболочка. Внешняя клеточная (плазматическая) мембрана. Структурно-химические особенности. Характеристика надмембранного слоя (гликокаликса) и подмембранного (кортикального) слоя. Морфологическая характеристика и механизмы барьерной, рецепторной и транспортной функций. Взаимосвязь плазматической мембраны над- и подмембранного слоев клеточной оболочки в процессе функционирования. Структурные и химические механизмы взаимодействия клеток.
2.2.3.Специализированные структуры клеточной оболочки: микроворсинки, реснички, базальные инвагинации. Их строение и функции. Общая характеристика межклеточных взаимодействий. Классификация. Межклеточные соединения (контакты): простые контакты, соединения типа замка, плотные соединения, десмосомы, щелевидные контакты (нексусы), синаптические соединения (синапсы).
2.2.4.Цитоплазма
2.2.4.1.Гиалоплазма. Физико-химические свойства, химический состав. Участие в клеточном метаболизме.
2.2.4.2.Органеллы. Определение, классификации. Органеллы общего и специального значения. Мембранные и немембранные органеллы.
2.2.4.2.1. Органеллы общего значения.
Мембранные:
Эндоплазматическая сеть. Строение и функции гранулярной и гладкой эндоплазматической сети. Особенности строения в зависимости от специфики метаболических процессов в клетке.
Пластинчатый комплекс (Комплекс Гольджи). Строение и функции. Его роль в выполнении железистыми клетками секреторной функции, в химической модификации поступающих белков. Значение во взаимодействии мембранных структур.
Лизосомы. Строение, химический состав, функции. Понятие о лизосомах, протеосомах, эндосомах, об аутофагосомах и гетерофагосомах.
Пероксисомы. Строение, химический состав, функции.
Митохондрии. Строение, функции. Представление об автономной системе синтеза белка. Особенности митохондриального аппарата в клетках с различным уровнем биоэнергетических процессов.
Немембранные:
Рибосомы. Строение, химический состав, функции. Понятие о полисомах. Роль свободных и связанных с мембранами эвдоплазматической сети рибосом в биосинтезе клеточных белков.
Центриоли. Строение и функции в неделящемся ядре и при митозе.
Фибриллярные структуры цитоплазмы. Цитоскелет. Основные компоненты цитоскелета: микротрубочки, микрофиламенты, тонофиламенты (промежуточные филаменты). Их строение, химический состав.
2.2.4.2.2.Органеллы специального значения: Миофибриллы, микроворсинки, реснички, жгутики. Строение и функциональное значение в клетках, выполняющих специальные функции.
2.2.4.3.Включения.Определение. Классификация. Значение в жизнедеятельности клеток и организма. Строение и химический состав различных видов включений.
2.2.4.4.Ядро. Роль ядра в хранении и передаче генетической информации и в синтезе белка. Форма и количество ядер. Понятие о ядерноцитоплазматнческом отношении. Общий план строения интерфазного ядра: хроматин, ядрышко, ядерная оболочка, кариоплазма (нуклеоплазма).
Кариоплазма (нуклеоплазма). Физико-химические свойства, химический состав. Значение в жизнедеятельности ядра.
Хроматин. Строение и химический состав. Структурно-химическая характеристика хроматиновых фибрилл, перихроматиновых фибрилл, перихроматиновых и интерхроматиновых гранул. Роль основных и кислых белков в структуризации и в регуляции метаболической активности хроматина. Понятие о нуклеосомах; механизм компактизации хроматиновых фибрилл. Понятие о деконденсированном и конденсированном хроматине (эухроматине, гетерохроматине, хромосомах), степень их участия в синтетических процессах. Строение хромосомы. Половой хроматин.
Ядрышко. Ядрышко как производное хромосом. Понятие о ядрышковом организаторе. Количество и размер ядрышек. Химический состав, строение, функция. Характеристика фибриллярных и гранулярных компонентов, их взаимосвязь с интенсивностью синтеза РНК. Структурно-функциональная лабильность ядрышкового аппарата.
Ядерная оболочка. Строение и функции. Структурно-функциональная характеристика наружной и внутренней мембран, перинуклеарного пространства, комплекса поры. Взаимосвязь количества ядерных пор и интенсивности метаболической активности клеток. Связь ядерной оболочки с эндоплазматической сетью; роль наружной мембраны в процессе новообразования клеточных мембран,
2.3. Основные проявления жизнедеятельности клеток
2.3.1.Синтетические процессы в клетке. Взаимосвязь компонентов клетки в процессах анаболизма и катаболизма. Понятие о секреторном цикле; механизмы поглощения и выделения продуктов в клетке.
2.3.2.Внутриклеточная регенерация. Общая характеристика и биологическое значение.
2.3.3.Информационные межклеточные взаимодействия. Гуморальные, синаптические, взаимодействия через внеклеточный матрикс и щелевые контакты.
2.3.4.Реакция клеток на внешние воздействия. Структурные и функциональные изменения клеток и отдельных клеточных компонентов в процессах реактивности и адаптации. Физиологическая и репаративная регенерация: сущность и механизмы.
2.4.Воспроизведение клеток
2.4.1.Клеточный цикл. Определение понятия; этапы клеточного цикла для клеток, сохранивших способность к делению, и клеток, утративших способность к делению.
2.4.2.Митотический цикл. Определение понятия. Фазы цикла (интерфаза, митоз). Биологическое значение митоза и его механизм. Преобразование структурных компонентов клетки на различных этапах митоза. Роль клеточного центра в митотическом делении клеток. Морфология митотических хромосом.
2.4.3.Эндомитоз. Определение понятия. Основные формы, биологическое значение. Понятие о плоидности клеток. Полиплоидия; механизмы образования полиплоидных клеток (одноядерных, многоядерных), функциональное значение этого явления.
2.4.4.Мейоз. Его механизм и биологическое значение.
2.4.5.Морфо-функциональная характеристика процессов роста и дифференцировки, периода активного функционирования, старения и гибели клеток.
2.5.Гибель клеток. Дегенерация, некроз. Определение понятия и его биологическое значение. Апоптоз (программированная гибель клеток). Определение понятия и его биологическое значение.
3. Общая гистология (учение о тканях).
3.1.Ткани как системы клеток и их производных — один из иерархических уровней организации живого. Клетки как ведущие элементы ткани. Неклеточные структуры — симпласты и межклеточное вещество как производные клеток. Синцитии. Понятие о клеточных популяциях. Клеточная популяция (клеточный тип, дифферон, клон). Статическая, растущая, обновляющаяся клеточные популяции. Стволовые клетки и их свойства. Детерминация и дифференциация клеток в ряду последовательных делений, коммитирование потенций. Диффероны. Тканевый тип, генез (гистогенез). Закономерности возникновения и эволюции тканей, теории параллелизма А.А.Заварзина и дивергентной эволюции Н.Г.Хлопина, их синтез на современном уровне развития науки.
3.1.2. Принципы классификации тканей. Классификация тканей.
3.1.3. Восстановительные способности тканей — типы физиологической регенерации в обновляющихся, растущих и стационарных клеточных популяциях, репаративная регенерация. Компенсаторно-приспособительные и адаптационные изменения тканей, их пределы.
3.2.Эпителиальные ткани.
3.2.1.Общая характеристика. Источники развития. Морфофункциональная и генетическая классификация эпителиальной ткани.
3.2.2.Покровные эпителии. Пограничность положения. Строение однослойных (однорядных и многорядных) и многослойных эпителиев (неороговевающих, ороговевающих, переходного). Принципы структурной организации и функции. Взаимосвязь морфофункционалъных особенностей эпителиальной ткани с ее пограничным положением в организме.
3.2.2.1.Базальная мембрана: строение, функции, происхождение. Особенности межклеточных контактов в различных видах эпителия. Горизонтальная и вертикальная анизоморфность эпителиальных пластов. Полярность эпителиоцитов и формы полярной дифференцировки их клеточной оболочки. Цитокератины как маркеры различных видов эпителиальных тканей. 3.2.2.2.Физиологическая и репаративная регенерация эпителия. Роль стволовых клеток в эпителиальных клетках обновляющегося типа; состав и скорость обновления их дифферонов в различных эпителиальных тканях.
3.2.3.Железистый эпителий. Особенности строения секреторных эпителиоцитов. Цитологическая характеристика эпителиоцитов, выделяющих секрет по голокриновому, апокриновому и мерокриновому типу.
3.2.3.1.Железы, их классификация. Характеристика концевых отделов и выводных протоков экзокринных желез. Особенности строения эндокринных желез.
3.3.Ткани внутренней среды
3.3.1.Кровь и лимфа. Гемоцитопоэз и лимфоцитопоэз.
3.3.1.2.Кровь Основные компоненты крови как ткани — плазма и форменные элементы. Функции крови. Содержание форменных элементов в крови взрослого человека. Формула крови. Возрастные и половые особенности крови.
3.3.1.2.1.Эритроциты: Размеры, форма, строение и функции, классификация эритроцитов по форме, размерам и степени зрелости. Особенности строения плазмолеммы эритроцита и его цитоскелета. Виды гемоглобина и связь с формой эритроцита. Ретикулоциты.
3.3.1.2.2.Лейкоциты: Классификация и общая характеристика. Лейкоцитарная формула. Гранулоциты — нейтрофилы, эозинофилы, базофилы, их содержание, размеры, форма, строение, основные функции. Особенности строения специфических гранул. Агранулоциты — моноциты, лимфоциты, количество, размеры, особенности строения и функции. Характеристика лимфоцитов — количество, морфофункциональные особенности, типы.
3.3.1.2.3.Кровяные пластинки (тромбоциты): Размеры, строение, функция.
3.3.1.3.Лимфа Лимфоплазма и форменные элементы. Связь с кровью, понятие о рециркуляции лимфоцитов.
3.3.1.4. Гемоцитопоэз и лимфоцитопоэз
3.3.1.4.1.Эмбриональный гемоцитопоэз. Развитие крови как ткани (гистогенез).
3.3.1.4.2.Постэмбриональный гемопоэз: физиологическая регенерация крови. Понятие о стволовых клетках крови (СКК) и колониеобразующих единицах (КОЕ). Характеристика плюрипотентных предшественников (стволовых, коммитированных клеток), унипотентных предшественников, бластных форм. Морфологически неидентифидируемые и морфологически идентифицируемые стадии развития клеток крови (характеристика клеток в дифферонах: эритроцитов, гранулоцитов, моноцитов, Т-лимфоцитов, В-лимфоцитов и кровяных пластинок (тромбоцитов). Особенности Т и В — лимфопоэза во взрослом организме. Регуляция гемопоэза и лимфопоэза, роль микроокружения.
3.3.2.Соединительные ткани
3.3.2.1.Общая характеристика соединительных тканей. Классификация. Источники развития. Гистогенез.
3.3.2.2.Волокнистые соединительные ткани.
3.3.2.2.1. Общая характеристика. Классификация.
3.3.2.2.2.Рыхлая волокнистая соединительная ткань. Клетки рыхлой волокнистой соединительной ткани. Фибробласты, их разновидности, фиброциты, миофибробласты, их происхождение, строение, участие в процессах фибриллогенеза. Макрофаги, их происхождение, виды, строение, роль в защитных реакциях организма. Понятие о системе мононуклеарных фагоцитов. Лейкоциты, их роль в защитных реакциях организма. Адипоциты (жировые клетки) белой и бурой жировой ткани, их происхождение, строение и значение. Перициты, адвентициальные клетки, их происхождение, строение и функциональная характеристика. Плазматические клетки, их происхождение, строение, роль в иммунитете. Тучные клетки, их происхождение, строение, функции. Пигментные клетки, их происхождение, строение, функция.
Межклеточное вещество. Общая характеристика и строение. Основное вещество, его физико-химические свойства и значение. Коллагеновые и эластические волокна, их роль, строение и химический состав. Представление о различных типах коллагена и их локализации в организме. Ретикулярные волокна. Происхождение межклеточного вещества. Возрастные изменения.
3.3.2.2.3.Плотная волокнистная соединительная ткань, ее разновидности, строение и функции. Сухожилие как орган.
3.3.2.2.4.Специализированные соединительные ткани. Ретикулярная ткань, строение, гистофизиология и значение. Жировая ткань, ее разновидности, строение и значение. Пигментная ткань, особенности строения и значение. Слизистая ткань, строение.
3.3.2.3.Скелетные ткани
3.3.2.3.1.Общая характеристика скелетных тканей. Классификация.
3.3.2.3.2.Хрящевые ткани. Общая характеристика. Виды хрящевой ткани (гиалиновая, эластическая, волокнистая). Хрящевые клетки — хондробласты, хондроциты (хондрокласты). Изогенные группы клеток. Гистохимическая характеристика и строение межклеточного вещества различных видов хрящевой ткани. Хондрогенез и возрастные изменения хрящевых тканей. Строение суставного хряща.
3.3.2.3.3.Костные ткани. Общая характеристика. Классификация. Клетки костной ткани: остеоциты, остеобласты, остеокласты. Их цито-функциональная характеристика. Межклеточное вещество костной ткани, его физико-химические свойства и строение. Ретикулофиброзная (грубоволокнистая) костная ткань. Пластинчатая (тонковолокнистая) костная ткань. Их локализация в организме и морфофункциональные особенности. Гистогенез и регенерация костных тканей. Возрастные изменения. Факторы, оказывающие влияние на строение костных тканей. Кость как орган.
3.4.Мышечные ткани
3.4.1.Общая характеристика и гистогенетическая классификация.
3.4.2.Cоматическая поперечно-полосатая (исчерченная) мышечная ткань. Развитие, морфологическая и функциональная характеристики. Микроскопическое и электронно-микроскопическое строение. Строение миофибриллы, ее структурно-функциональная единица (саркомер). Механизм мышечного сокращения. Типы мышечных волокон и их иннервация. Моторная единица. Миосателлитоциты. Регенерация мышечной ткани, значение миосателлитоцитов. Мышца как орган. Связь с сухожилием.
3.4.3.Сердечная поперечно-полосатая (исчерченная) мышечная ткань. Источник развития, этапы гистогенеза. Морфофункциональная характеристика рабочих и проводящих кардиомиоцитов. Возможности регенерации.
3.4.4.Гладкая (неисчерченная) мышечная ткань. Источник развития. Морфологическая и функциональная характеристика. Регенерация,
3.4.5.Мионейральная ткань. Источник развития, строение и функция.
3.4.6.Миоидные и моэпителиальные клетки. Источники развития. Строение. Функции.
3.5.Нервная ткань.
3.5.1.Общая характеристика нервной ткани. Эмбриональный гистогенез. Дифференцировка нейробластов и глиобластов. Понятие о регенерации структурных компонентов нервной ткани.
3.5.2.Нейроциты (нейроны). Источники развития. Морфологическая и функциональная классификация. Общий план строения нейрона. Микро- и ультраструктура перикариона (тела нейрона), аксона, дендритов. Базофильное вещество (субстанция Ниссля). Особенности цитоскелета нейроцитов (нейрофиламенты и нейротрубочки). Роль плазмолеммы нейроцитов в рецепции, генерации и проведении нервного импульса. Транспортные процессы в цитоплазме нейронов. Аксональный транспорт — антероградный и ретроградный. Быстрый и медленный транспорт, роль микротрубочек. Понятие о нейромедиаторах. Секреторные нейроны, особенности их строения и функция. Физиологическая гибель нейронов. Регенерация нейронов.
3.5.3.Нейроглия. Общая характеристика. Источники развития глиоцитов. Классификация.
Макроглия (олигодендроглия, астроглия и эпендимная глия). Микроглия.
3.5.4.Нервные волокна. Общая характеристика. Классификация. Особенности формирования, строения и функции безмиелиновых и миелиновых нервных волокон. Понятие об осевом цилиндре и мезаксоне. Ультрамикроскопическое строение миелиновой оболочки. Дегенерация и регенерация нервных волокон.
3.5.5.Нервные окончания. Общая характеристика. Классификация. Рецепторные (чувствительные) нервные окончания – свободные, несвободные и инкапсулированные, нервно-мышечные веретена, нервно-сухожильные веретена, комплекс клетки Меркеля с нервной терминалью. Эффекторные окончания — двигательные и секреторные. Нервно-мышечное окончание (моторная бляшка) в скелетных мышцах и в гладкой мышечной ткани. Секреторные (нейро-железистые) нервные окончания.
3.5.6.Синапсы. Классификации. Межнейрональные электрические, химические и смешанные синапсы, строение и механизмы передачи возбуждения. Ультраструктура химических синансов — пресинапгическая и постсинаптическая части, синаптические пузырьки, синаптическая щель. Рефлекторные дуги, их чувствительные, двигательные и ассоциативные звенья.
СПЕЦИАЛЬНАЯ ЧАСТЬ
4. Частная гистология
4.1.Нервная система.
4.1.1.Общая характеристика. Источники и ход эмбрионального развития. Нервная трубка и ее дифференцировка на вентрикулярную, субвентрикулярную (камбиальную), промежуточную (плащевую) и маргинальную зоны. Нервный гребень и плакоды, их дифференцировка. Органогенез.
4.1.2.Периферическая нервная система
4.1.2.1.Нерв. Строение, тканевой состав. Реакция на повреждение, регенерация.
4.1.2.2.Чувствительные нервные узлы (спинномозговые и черепные). Строение, тканевой состав. Характеристика нейронов и нейроглии.
4.1.3.Центральная нервная система
4.1.3.1.Строение серого и белого вещества. Понятие о рефлекторной дуге (нейронный состав и проводящие пути) и о нервных центрах. Строение оболочек мозга — твердой, паутинной, мягкой. Субдуральное и субарахноидальное пространства, сосудистые сплетения. Особенности строения сосудов (синусы, гемокапилляры) центральной нервной системы.
4.1.3.2.Спинной мозг. Общая характеристика строения. Строение серого вещества: виды нейронов и их участие в образовании рефлекторных дуг, типы глиоцитов. Ядра серого вещества. Строение белого вещества. Центральный канал спинного мозга и спинно-мозговая жидкость.
4.1.3.3.Головной мозг.
4.1.3.3.1.Мозжечок. Строение и нейронный состав коры мозжечка. Грушевидные клетки, корзинчатые и звездчатые нейроциты, клетки-зерна. Афферентные и эфферентные нервные волокна. Межнейрональные связи, тормозные нейроны. Клубочек мозжечка. Глиоциты мозжечка.
4.1.3.3.2.Ствол мозга. Строение и нейронный состав.
4.1.3.3.3.Головной мозг. Общая характеристика строения, особенности строения и взаимоотношения серого и белого вещества. Кора большого мозга. Эмбриональный и постэмбриональный гистогенез. Цитоархитектоника слоев (пластинок) коры больших полушарий. Нейронный состав, характеристика пирамидных нейронов. Представление о модульной организации коры. Межнейрональные связи, особенности строения синапсов. Тормозные нейроны. Глиоциты коры. Миелоархитектоника — радиальные и тангенциальные нервные волокна. Особенности строения коры в двигательных и чувствительных зонах.
Гематоэнцефалический барьер, его строение и функция.
4.1.4.Автономная (вегетативная) нервная система
4.1.4.1.Общая характеристика строения центральных и периферических отделов парасимпатической и симпатической систем. Строение и нейронный состав ганглиев (экстрамуральных и интрамуральных). Пре- и постганглионарные нервные волокна.
4.2.Сенсорная система (Органы чувств)
4.2.1.Классификация. Общий принцип клеточной организации рецепторных отделов. Нейросенсорные и сенсоэпителиальные рецепторные клетки.
4.2.2.Орган зрения. Общая характеристика. Источники эмбрионального развития и гистогенез.
4.2.2.1.Общий план строения глазного яблока. Оболочки, их отделы и производные, тканевой состав. Основные функциональные аппараты: диоптрический, аккомодационный и рецепторный. Строение и роль составляющих их роговицы, хрусталика, стекловидного тела, радужки, сетчатки. Нейронный состав и глиоциты сетчатки, их морфофункциональная характеристика. Строение и патофизиология палочко- и колбочконесущих нейронов сетчатки. Особенности строения центральной ямки диска зрительного нерва. Пигментный эпителий сетчатки, строение и значение. Особенности кровоснабжения глазного яблока. Морфологические основы циркуляции внутриглазной жидкости. Возрастные изменения.
4.2.2.2. Вспомогательные органы глаза (веки, слезный аппарат).
4.2.3.Орган обоняния. Общая характеристика. Эмбриональное развитие. Строение и клеточный состав обонятельной выстилки: рецепторные, поддерживающие и базальные клетки. Гистофизиология органа обоняния. Возрастные изменения. Вомеро-назальный орган.
4.2.4.Орган вкуса. Общая характеристика. Эмбриональное развитие. Строение и клеточный состав вкусовых почек: вкусовые, поддерживающие и базальные клетки. Иннервация вкусовых почек. Гистофизиология органа вкуса. Возрастные изменения.
4.2.5.Органы слуха и равновесия. Общая характеристика. Эмбриональное развитие.
4.2.5.1.Наружное ухо: строение наружного слухового прохода и барабанной перепонки. Среднее ухо: слуховые косточки, характеристика эпителия барабанной полости и слуховой трубы.
4.2.5.2.Внутреннее ухо: костный и перепончатый лабиринты.
4.2.5.2.1.Вестибулярная часть перепончатого лабиринта: эллиптический и сферический мешочки и полукружные каналы. Их рецепторные отделы: строение и клеточный состав пятна и ампулярных гребешков. Иннервация. Гистофизиология вестибулярного лабиринта.
4.2.5.2.2.Улитковая часть перепончатого лабиринта: строение улиткового канала, строение и клеточный состав спирального органа, его иннервация. Гистофизиология восприятия звуков. Возрастные изменения.
4.3.Сердечно-сосудистая система.
4.3.1.Строение и эмбриональное развитие сердечно-сосудистой системы.
4.3.2.Кровеносные сосуды. Общие принципы строения, тканевой состав. Классификация сосудов. Понятие о микроциркуляторном русле. Зависимость строения сосудов от гемодинамических условий. Васкуляризация сосудов (сосуды сосудов). Ангиогенез, регенерация сосудов. Возрастные изменения в сосудистой стенке.
4.3.2.1.Артерии. Классификация. Особенности строения и функции артерий различного типа: мышечного, мышечно-эластического и эластического. Органные особенности артерий.
4.3.2.2.Микроциркуляторное русло.
4.3.2.2.1.Артериолы, их виды и роль в кровообращении. Строение. Значение эндотелиомиоцитных контактов в гистофизиологии артериол.
4.3.2.2.2.Гемокапилляры. Классификация, функция и строение. Морфологические основы процесса проницаемости капилляров и регуляции их функций. Органные особенности капилляров.
4.3.2.2.3.Венулы. Их виды, функциональное значение, строение.
4.3.2.2.4.Артериоловенулярные анастомозы. Значение для кровообращения. Классификация. Строение артериоловенулярных анастомозов различного типа.
4.3.2.3.Вены. Строение стенки вен в связи с гемодинамическими условиями. Классификация. Особенности строения вен различного типа (мышечного и безмышечного). Строение венозных клапанов. Органные особенности вен.
4.3.2.4.Лимфатические сосуды. Строение и классификация. Строение лимфатических капилляров и различных видов лимфатических сосудов. Понятие о лимфангионе. Участие лимфатических капилляров в системе микроциркуляции.
4.3.3.Сердце. Эмбриональное развитие. Строение стенки сердца, его оболочек, их тканевой состав. Эндокард и клапаны сердца. Миокард, рабочие, проводящие и секреторные кардиомиоциты. Особенности кровоснабжения, регенерации. Проводящая система сердца, ее морфофункциональная характеристика. Эпикард и перикард. Внутриорганные сосуды сердца. Иннервация сердца. Сердце новорожденного. Перестройка и развитие сердца после рождения. Возрастные изменения сердца.
4.4.Система органов кроветворения и иммунной защиты.
4.4.1.Общая характеристика системы кроветворения и иммунной защиты. Основные источники и этапы формирования органов кроветворения в онтогенезе человека. Мезобластический, гепатомсленотимический и медуллярный этапы становления системы кроветворения.
4.4.2.Центральные органы кроветворения и иммуногенеза
4.4.2.1.Костный мозг. Строение, тканевой состав и функции красного костного мозга. Особенности васкуляризации и строение гемокапилляров. Понятие о микроокружении. Желтый костный мозг. Развитие костного мозга во внутриутробном периоде. Возрастные изменения. Регенерация костного мозга.
4.4.2.2.Тимус. Эмбриональное развитие. Роль в лимфоцитопоэзе. Строение и тканевой состав коркового и мозгового вещества долек. Васкуляризация. Строение и значение гематотимического барьера. Временная (акциденталъная) и возрастная инволюция тимуса.
4.4.3. Периферические органы кроветворения и иммуногенеза
4.4.3.1.Селезенка. Эмбриональное развитие. Строение и тканевой состав (белая и красная пульпа, Т- и В- зависимые зоны). Кровоснабжение селезенки. Структурные и функциональные особенности венозных синусов.
4.4.3.2.Лимфатические узлы. Эмбриональное развитие. Строение и тканевой состав. Корковое и мозговое вещество. Их морфофункциональная характеристика, клеточный состав. Т- и В-зависимые зоны. Система синусов. Васкуляризация. Роль кровеносных сосудов в развитии и гистофизиологии лимфатических узлов. Возрастные изменения.
4.4.3.3.Лимфоидные образования в составе слизистых оболочек: лимфоидные узелки и диффузные скопления в стенке воздухоносных путей, пищеварительного тракта (одиночные и множественные) и других органов. Их строение, клеточный состав и значение.
4.4.4. Морфологические основы защитных реакций организма.
4.4.4.1.Воспаление, заживление, восстановление. Клеточные основы воспалительной реакции (роль нейтрофильных и базофильных лейкоцитов, моноцитов) и процесса заживления ран.
4.4.4.2. Иммунитет. Виды. Характеристика основных клеток, осуществляющих иммунные реакции — нейтрофильных лейкоцитов, макрофагов, антигенпредставляющих клеток, Т-лимфопитов, В-лимфодитов, плазмоцитов. Понятие об антигенах и антителах. Антигеннезависимая и антигензависимая пролиферация лимфоцитов. Процессы лимфоцитопоэза в Т- и В-зависимых зонах периферических лимфоидных органов. Понятие о циркуляции и рециркуляции Т- и В-лимфопитов. Гуморальный и клеточный иммунитет — особенности кооперации макрофагов, антигенпредставляющих клеток, Т- и В-лимфоцитов. Эффекторные клетки и клетки памяти в гуморальном и клеточном иммунитете. Естественные киллеры. Плазматические клетки и стадии их дифференциации. Регуляция иммунных реакций: цитокины, гормоны.
4.5. Эндокринная система.
4.5.1.Общая характеристика и классификация эндокринной системы. Центральные и периферические звенья эндокринной системы. Понятие о гормонах, клетках-мишенях и их рецепторах к гормонам. Механизмы регуляции в эндокринной системе. Классификация эндокринных желез.
4.5.2.Гипоталамо-гипофизарная нейросекреторная система
4.5.2.1.Гипоталамус. Нейроэндокринные нейроны крупноклеточных и мелкоклеточных ядер гипоталамуса. Гипоталамоаденогипофизарная и гипоталамонейрогипофизарная системы. Либерины и статины, их роль в регуляции эндокринной системы. Регуляция функций гипоталамуса центральной нервной системой.
4.5.2.2.Гипофиз. Эмбриональное развитие. Строение и функции аденогипофиза. Цитофункциональная характеристика аденоцитов передней доли гипофиза. Гипоталамоаденогипофизарное кровообращение, его роль во взаимодействии гипоталамуса и гипофиза. Средняя (промежуточная) доля гипофиза и ее особенности у человека. Строение и функция нейрогипофиза, его связь с гипоталамусом. Васкуляризация и иннервация гипофиза. Возрастные изменения.
4.5.3.Эпифиз мозга. Строение, клеточный состав, функция. Возрастные изменения.
4.5.4.Периферические эндокринные железы
4.5.4.1.Щитовидная железа. Источники развития. Строение. Фолликулы как
морфофункциональные единицы, строение стенки и состав коллоида фолликулов. Фолликулярные эндокриноциты (тироциты), их гормоны и фазы секреторного цикла. Роль гормонов тироцитов. Перестройка фолликулов в связи с различной функциональной активностью. Парафолликулярные эндокриноциты (кальцитониноциты, С-клетки). Источники развития, локализация и функция. Фолликулогенез. Васкуляризация и иннервация щитовидной железы.
4.5.4.2.Околощитовидные железы. Источники развития. Строение и клеточный состав. Роль в регуляции минерального обмена. Васкуляризация, иннервация и механизмы регуляции околощитовидных желез. Структура околощитовидных желез у новорожденных и возрастные изменения.
4.5.4.3.Надпочечники. Источники развития. Фетальная и дефинитивная кора надпочечников. Зоны коры и их клеточный состав. Особенности строения корковых эндокриноцитов в связи с синтезом и секрецией кортикостероидов. Роль гормонов коры надпочечников в регуляции водно-солевого равновесия, развитии общего адаптационного синдрома, регуляции белкового синтеза. Мозговое вещество надпочечников. Строение, клеточный состав, гормоны и роль мозговых эндокриноцитов (эпинефроцитов). Возрастные изменения надпочечника.
4.5.5.Эндокринные структуры желез смешанной секреции.
4.5.5.1.Эндокринные островки поджелудочной железы. Эндокринная функция гонад (яичек, яичников), плаценты.
4.5.6.Одиночные гормонопродуцирующие клетки.
4.5.6.1.Представление о диффузной эндокринной системе (ДЭС), локализация элементов, их клеточный состав. Нейроэндокринные клетки. Представления о АПУД системе.
4.6. Пищеварительная система.
4.6.1.Общая характеристика пищеварительной системы. Основные источники развития тканей пищеварительной системы в эмбриогенезе. Общий принцип строения стенки пищеварительного канала — слизистая оболочка, подслизистая основа, мышечная оболочка, наружная оболочка (серозная или адвентициальная), их тканевой и клеточный состав. Понятие о слизистой оболочке, ее строение и функция. Иннервация и васкуляризапия стенки пищеварительной трубки. Эндокринный аппарат пищеварительной системы. Лимфоидные структуры пищеварительного тракта. Строение брюшины.
4.6.2.Передний отдел пищеварительной системы Особенности строения стенки различных отделов, развитие.
4.6.2.1.Ротовая полость. Строение слизистой оболочки в связи с функцией и особенностями пищеварения в ротовой полости.
4.6.2.1.1.Строение губы, щеки, твердого и мягкого неба, языка, дёсны, миндалины.
4.6.2.1.2.Большие слюнные железы. Классификация, источники развития, строение и функции. Строение секреторных отделов выводных протоков. Эндокринная функция.
4.6.2.1.3.Язык. Строение. Особенности строения слизистой оболочки на верхней и нижней поверхностях органа. Сосочки языка, их виды, строение, функции.
4.6.2.1.4.Зубы. Строение. Эмаль, дентин и цемент — строение, функция и химический состав. Пульпа зуба — строение и значение. Периодонт — строение и значение. Кровоснабжение и иннервация зуба. Развитие и смена зубов. Возрастные изменения.
4.6.2.2.Глотка и пищевод. Строение и тканевой состав стенки глотки и пищевода в различных его отделах. Железы пищевода, их гистофизиология.
4.6.3.Средний и задний отделы пищеварительной системы. Особенности строения стенки
различных отделов. Развитие.
4.6.3.1.Желудок. Строение слизистой оболочки в различных отделах органа. Цитофизиологическая характеристика покровного эпителия, слизеобразование. Локализация, строение и клеточный состав желез в различных отделах желудка. Микро- и ультрамикроскопические особенности экзо- и эндокринных клеток. Регенерация покровного эпителия и эпителия желез желудка. Кровоснабжение и иннервация желудка. Возрастные особенности строения желудка.
4.6.3.2.Тонкая кишка. Характеристика различных отделов тонкой кишки. Строение стенки, ее тканевый состав. Система «крипта-ворсинка» как структурнофункциональная единица. Виды клеток эпителия ворсинок и крипт, их строение и цитофизиология. Гистофизиология процесса пристеночного пищеварения и всасывания. Роль слизи и микроворсинок энтероцитов в пристеночном пищеварении. Цитофизиология экзо- и эндокринных клеток. Регенерация эпителия тонкой кишки. Кровоснабжение и иннервация стенки тонкой кишки. Возрастные изменения стенки тонкой кишки. Лимфоидные образования в стенке кишки.
4.6.3.3.Толстая кишка. Характеристика различных отделов. Строение стенки, ее тканевый состав. Особенности строения слизистой оболочки в связи с функцией. Виды эпителиоцитов и эндокриноцитов, их цитофизиология. Лимфоидные образования стенки. Кровоснабжение.
4.6.3.3.1.Червеобразный отросток.Особенности строения и функции.
4.6.3.3.2.Прямая кишка. Строение стенки.
4.6.4.Поджелудочная железа. Общая характеристика. Строение экзокринного и эндокринного отделов. Цитофизиологическая характеристика ацинарных клеток. Типы эндокриноцитов островков и их морфофункциональная характеристика. Кровоснабжение. Иннервация. Регенерация. Особенности гистофизиологии в разные периоды детства. Изменения железы при старении организма.
4.6.5.Печень. Общая характеристика. Особенности кровоснабжения. Строение классической дольки как структурно-функциональной единицы печени. Представления о портальной дольке и ацинусе. Строение внутридольковых синусоидных сосудов, цитофизиология их клеточных элементов: эндотелиоцитов, макрофагов. Перисинусоидальные пространства, их структурная организация. Липоциты, особенности строения и функции. Гепатопиты — основной клеточный элемент печени, представления об их расположении в дольках, строение в связи с функциями печени. Строение желчных канальцев (холангиол) и междольковых желчных протоков. Механизмы циркуляции по ним желчи. Иннервация. Регенерация. Особенности строения печени новорожденных. Возрастные особенности.
4.6.5.1.Желчный пузырь и желчевыводящие пути. Строение и функция.
4.7. Дыхательная система.
4.7.1. Общая характеристика дыхательной системы. Воздухоносные пути и респираторный отдел. Развитие. Возрастные особенности. Регенерация.
4.7.2. Внелегочные воздухоносные пути. Особенности строения стенки воздухоносных путей: носовой полости, гортани, трахеи и главных бронхов. Тканевой состав и гистофункциональная характеристика их оболочек. Клеточный состав эпителия слизистой оболочки.
4.7.3.Легкие. Внутрилегочные воздухоносные пути: бронхи и бронхиолы, строение их стенок в зависимости от их калибра.
4.7.3.1.Ацинус как морфофункциональная единица легкого. Структурные компоненты ацинуса. Строение стенки альвеол. Типы пневмоцитов, их цитофункциональная характеристика. Структурно-химическая организация и функция сурфактантно-альвеолярного комплекса. Строение межальвеолярных перегородок. Аэрогематический барьер и его значение в газообмене. Макрофаги легкого. Кровоснабжение легкого.
4.7.3.2.Плевра. Морфо-функциональная характеристика.
4.8. Кожа и ее производные.
4.8.1.Кожа. Общая характеристика. Тканевый состав. развитие. Регенерация.
4.8.1.1.Эпидермис. Основные диффероны клеток в эпидермисе. Слои эпидермиса. Их клеточный состав. Антигенпредставляющие клетки кожи. Особенности строения эпидермиса «толстой» и «тонкой» кожи. Понятие о процессе кератинизации, его значение. Клеточное обновление эпидермиса и представление о его пролиферативных единицах и колонковой организации. Местная система иммунного надзора эпидермиса – клетки Лангерганса и лимфоциты, их гистофункциональная характеристика. Пигментные клетки эпидермиса, их происхождение, строение и роль. Осязательные клетки. Базальная мембрана, дермальноэпидермальное соединение.
4.8.1.2.Дерма, Сосочковый и сетчатый слои, их тканевой состав. Особенности строения дермы в коже различных участков тела — стопы, ладоней, лица, суставов и др. Гистофункциональная характеристика иммунной системы в дерме. Васкуляризация кожи. Гиподерма.
4.8.2.Железы кожи. Сальные и потовые железы (меро- и апокриновые), их развитие, строение, гистофизиология. Молочные железы — см. в разделе «Женская половая система». Возрастные особенности кожи и ее желез.
4.8.3.Придатки кожи. Волосы. Развитие, строение, рост и смена волос, иннервация. Ногти. Развитие, строение и рост ногтей.
4.9. Система органов мочеобразования и мочевыведения.
4.9.1.Общая характеристика системы мочевых органов. Развитие.
4.9.2.Почки, Корковое и мозговое вещество почки. Нефрон — как морфофункциональная единица почки, его строение. Типы нефронов, их топография в корковом и мозговом веществе. Васкуляризация почки — кортикальная и юкстамедуллярная системы кровоснабжения. Почечные тельца, их основные компоненты. Строение сосудистых клубочков. Мезангий, его строение и функция. Структурная организация почечного фильтра и роль в мочеобразовании. Юкстагломерулярный аппарат. Гистофизиология канальцев нефронов и собирательных трубочек в связи с их участием в образовании окончательной мочи. Строма почек, ее гистофункциональная характеристика. Понятие о противоточной системе почки. Морфофункциональные основы регуляции процесса мочеобразования. Эндокринный аппарат почки (ренин-ангиотензиновая, интерстициальная простагландиновая и калликреин-кининовая системы), строение и функция. Иннервация почки. Регенеративные потенции. Особенности почки у новорожденного. Последующие возрастные изменения почки.
4.9.3. Мочевыводящие пути. Строение стенки почечных чашечек и лоханки. Строение мочеточников. Строение мочевого пузыря. Понятие о цистоидах. Особенности строения мужского и женского мочеиспускательного канала.
4.10. Половые системы.
4.10.1.Развитие. Первичные гоноциты, начальная локализация, пути миграции в зачаток гонады. Половая дифференцировка.
4.10.2.Мужские половые органы. Гистогенетические процессы в зачатке гонады, ведущие к развитию яичка. Развитие семявыносящих путей.
4.10.2.1.Яичко. Строение. Извитые семенные канальцы, строение стенки. Сперматогенез. Цитологическая характеристика его основных фаз. Роль сустентоцитов в сперматогенезе. Гематотестикулярный барьер. Эндокринная функция яичка: мужские половые гормоны и синтезирующие их гландулоциты (клетки Лейдига), их цитохимические особенности, участие в регуляции сперматогенеза. Гистофизиология прямых канальцев, канальцев сети и выносящих канальцев яичка. Регуляция генеративной и эндокринной функций яичка. Возрастные особенности.
4.10.2.2.Семявыносящие пути. Придаток яичка. Семявыносящий проток. Семенные железы.
Семяизвергательный канал. Бульбо-уретральные железы. Предстательная железа. Их строение и функции. Возрастные изменения. Половой член. Строение.
4.10.3.Женские половые органы.
4.10.3.1.Яичник. Развитие. Общая характеристика строения. Особенности строения коркового и мозгового вещества. Овогенез. Отличия овогенеза от сперматогенеза. Строение и развитие фолликулов. Овуляция. Понятие об овариальном цикле и его регуляции. Развитие, строение и функции желтого тела в течение овариального цикла и при беременности. Атрезия фолликулов. Эндокринная функция яичника: женские половые гормоны и вырабатывающие их клеточные элементы. Возрастные особенности.
4.10.3.2.Матка. Развитие. Строение стенки матки в разных ее отделах. Менструальный цикл и его фазы. Особенности строения эндометрия в различные фазы цикла. Связь циклических изменений эндометрия и яичника. Перестройка матки при беременности и после родов. Васкуляризация и иннервация матки. Возрастные изменения.
4.10.3.3.Маточные трубы. Развитие, строение и функции.
4.10.3.4.Влагалище. Развитие. Строение его стенок. Изменение в связи с менструальным циклом.
4.10.4.Молочная (грудная) железа. Происхождение. Развитие. Строение. Постнатальные изменения. Функциональная морфология лактирующей и нелактирующей (нефункционирующей и после лактации) молочной железы. Нейроэндокринная регуляция функций молочных желез. Изменение молочных желез в ходе овариально-менструального цикла и при беременности.
5. Эмбриология человека (Ранний эмбриогенез).
5.1.Эмбриология млекопитающих как основа для понимания особенностей эмбрионального развития человека. Периодизация развития человека и животных. Представление о биологических процессах, лежащих в основе развития зародыша — индукция, детерминация, деление, миграция клеток, рост, дифференцировка, взаимодействие клеток, гибель клеток. Особенности эмбрионального развития человека. Критические периоды в развитии. Нарушение процессов детерминации как причина аномалий и уродств.
5.2.Прогенез. Сперматогенез. Овогенез. Особенности структуры половых клеток.
5.3.Оплодотворение. Биологическое значение оплодотворения, особенности и хронология процесса. Дистантные и контактные взаимодействия половых клеток. Преобразования в спермии: капацитация, акросомальная реакция, пенетрация спермием прозрачной зоны и плазмолеммы овоцита, сброс цитоплазматической оболочки спермия, поворот спермия, формирование мужского пронуклеуса.
5.4.Преобразования в овоците: рассеивание клеток лучистого венца, кортикальная реакция, выброс ферментов кортикальных гранул, преобравание прозрачной зоны (зонная реакция), активация цитоплазматических процессов, окончание мейоза, полярные тельца.
5.5.Мужской и женский пронуклеусы, распад их оболочек, установление связи хромосом пронуклеусов с центриолью спермия.
5.6.Первая неделя развития. Зигота — одноклеточный зародыш, ее геном, активация внутриклеточных процессов.
5.6.1.Дробление. Специфика дробления у человека и хронология процесса. Строение зародыша на разных стадиях дробления. Роль прозрачной зоны. Характеристика темных и светлых бластомеров, их межклеточных контактов. Уменьшение размеров бластомеров, их взаимодействие. Морула. Бластоциста. Внутренняя клеточная масса (эмбриобласт) и трофобласт. Стадия свободной бластоцисты. Состояние матки к началу имплантации. Начало 1-й фазы гаструляпии.
5.6.2.Имплантация. Хронология процесса имплантации. Дифференцировка трофобласта на цитотрофобласт и синцитиотрофобласт. Активация синцитиотрофобласта. Образование лакун и их соединение с кровеносными сосудами эндометрия. Гистиотрофный тип питания. Формирование первичных и вторичных ворсин хориона.
5.7.Вторая неделя развития. Гаструляция. Разделение эмбриобласта на эпибласт и гипобласт. Преобразование гипобласта, формирование первичного желточного мешка.
5.7.1.Преобразование эпибласта: образование амниотической полости и выделение амниотической эктодермы, формирование амниотического пузыря; начало 2-й фазы гаструляпии путем эмиграции — формирование первичной полоски и первичного узелка, образование зародышевой мезодермы, головного отростка, энтодермы зародыша, образование прехордальной пластики. Образование внезародыщевой мезодермы.
5.8.Третья неделя развития. Дифференцировка зародышевой мезодермы (сомиты, нефрогонотомы, висцеральный и париетальный листки спланхнотома, эмбриональный целом). Рост головного отростка, образование хорды. Формирование нервной трубки и нервных гребней, асинхронность развития головного и каудального отделов. Туловищная складка, образование первичной кишки.
5.8.1.Дифференцировка внезародышевой мезодермы аллантоиса, амниотического пузыря, желточного стебля, соединительной ножки, слоя, подстилающего трофобласт.
5.8.2.Формирование первичных кровеносных сосудов и первичных клеток крови в мезодерме желточного мешка, соединительной ножки. Формирование первых кровеносных сосудов в мезодерме зародыша. Зачаток первичного сердца, начало функции. Закладка предпочки, легкого.
5.8.3.Образование третичных ворсин хориона. Гемотрофный тип питания.
5.9.Четвертая неделя развития. Изменение формы зародыша (образование поперечных и продольных складок). Завершение процессов нейруляции и сегментации мезодермы. Ушная и хрусталиковая плакоды. Развитие мезонефроса. Миграция гоноцитов из желточной энтодермы каудального конца зародыша. Образование рта (прорыв орофарингеальной мембраны), формирование позвоночного столба. Закладка аденогипофиза, щитовидной и околощитовидной желез, желудка, печени, дорзальной части поджелудочной железы.
5.9.1.Эмбриональный органогенез.
5.10.Внезародышевые органы.
5.10.1.Плацента, формирования, особенности организации материнского и фетального компонентов на протяжении беременности. Опережающее развитие соединительной ткани плаценты и других внезародышевых органов. Структурные отличия терминальных и дифенитивных ворсинок в разных триместрах беременности, функции плаценты.
5.10.2.Амнион, его строение и значение.
5.10.3.Пуповина, ее образование и структурные компоненты: студенистая (слизистая) ткань, сосуды, рудименты желточного мешка и аллантоиса. Система мать-плацента-плод и факторы, влияющие на ее физиологию.
5.11.Особенности организма новорожденного. Общая характеристика и периодизация постнатального развития.
5.12.Факторы, влияющие на развитие: генетические, материнские, внешние (радиация, алкоголь, курение, наркотики, инфекция, химические и лекарственные вещества, пестициды и др.).
Дополнение к программе
по профильным разделам частной гистологии для стоматологических факультетов
Гистология и эмбриология органов полости рта
Общая морфофункциональная характеристика. Строение стенки пищеварительного канала: слизистая оболочка, подслизистая основа, мышечная, серозная и адвентициальная оболочка. Васкуляризация и иннервация пищеварительного канала.
Развитие. Эмбриональная кишечная трубка. Ротовая и анальная бухты. Развитие и тканевые источники оболочек кишки в ее различных отделах. Формирование передней, средней и задней кишки.
Полость рта. Гистофункциональная характеристика слизистой оболочки; структурные и гистохимические особенности клеток эпителия его слизистой оболочки, кровоснабжение и иннервация. Ороговение в эпителии слизистой оболочки ротовой полости. Возрастные особенности. Собственная пластинка слизистой оболочки, ее состав. Разновидности слизистой оболочки ротовой полости (жевательная, выстилающая, специализированная). Подслизистая основа.
Десна. Строение и гистохимическая характеристика. Многослойный плоский ороговевающий эпителий и собственная пластинка слизистой оболочки десны. Десна свободная и прикрепленная. Межзубные сосочки десны. Десневой желобок, десневая щель (бороздка) и карман. Эпителиальное прикрепление.
Твердое небо. Особенности железистой и жировой части твердого неба. Краевая зона и небный шов. Особенности слизистой оболочки на их ротовой и носовой поверхностях неба.
Губы. Характеристика кожной, переходной и слизистой частей. Губные железы. Строение спайки губ. Возрастные изменения.
Щеки. Их максилярная, мандибулярная и промежуточные зоны. Щечные железы. Жировое тело щеки. Слизистая оболочка альвеолярных отростков челюстей. Мягкое небо. Язычок.
Дно ротовой полости. Переходная складка губы и щеки. Строение уздечек губ, подъязычной складки, подъязычного мясца и сосочка околоушной железы.
Язык. Его развитие и строение. Слизистая оболочка языка, особенности ее строения на нижней, верхней и боковых поверхностях, сосочках. Нитевидные, грибовидные, окруженные валом и листовидные сосочки языка. Вкусовой аппарат. Вкусовые луковюцд. Железы языка. Апоневроз и перегородки языка. Мышечное тело языка. Уздечка языка.
Особенности иннервации и кровоснабжения языка.
Специализированные лимфоидные образования ротовой полости. Лимфозпителиальное глоточное кольцо. Миндалины, Язычная миндалина, небные, глоточная и другие миндалины. Их локализация, особенности строения и развитие. Ямки и крипты миндалины. Лимфатические фолликулы миндалины и их клеточные элементы. Дольки и капсула миндалины. Иннервация и кровоснабжение миндалин. Гистофизиология лимфоэпителиального глоточного кольца. Возрастные изменения миндалин.
Железы ротовой полости. Слюнные железы. Мелкие слюнные железы. Особенности и распределение. Большие слюнные железы рта. Строение, развитие и гистофизиология. Микроскопическое и улырамикроскогшческое строение концевых отделов и выводных протоков. Особенности белковых, слизистых и смешанных концевых отделов. Исчерченные слюнные протоки и их значение в процессах секреции и реабсорбции. Слюна, ее химический состав и значение. Особенности развития и строения околоушных, подчелюстных и подъязычных слюнных желез. Эндокринная функция слюнных желез. Кровоснабжение и иннервация слюнных желез. Возрастные изменения и регенерация желез.
Зубы. Общая морфофункциональная характеристика зубов. Понятие о твердых и мягких тканях зуба. Эмаль. Ее микроскопическое и улырамикроскогшческое строение, физико-химические свойства. Форма и строение эмалевых призм. Радиальные светлые и темные полосы эмали и тангенциальные линии. Эмалевые пучки и пластинки, эмалевые веретена. Межпризматическое вещество. Апризматическая эмаль. Особенности обызвествления и обмена веществ в эмали.
Особенности строения эмали различных зубов. Эмалево-дептинные и эмалево-цементные соединения. Кутикула, пелликула и их роль в проникновении неорганических веществ в эмаль. Строение поверхностного слоя эмали у детей до 1 года жизни и взрослого человека. Возрастные изменения эмали. Дентин, его микроскопическая и ультрамикроскопическая характеристика. Основное вещество дентина. Дентинные волокна, радиальные и тангенциальные. Дентинные трубочки и обызвествление дентина. Дентинные шары.
Интерглобулярный дентин. Зернистый слой. Плащевой и околопульпарный дентин. Контурные линии дентина. Предентин. Питание и иннервация дентина. Первичный и вторичный дентин. Прозрачный дентин. Реакция дентина на повреждение. «Мертвые пути» в дентине. Цемент. Его расположение, химический состав, обызвествление. Эмалево-цементная и цементно-дентинная граница. Строение цемента. Клеточный и безклеточный цемент. Цементоциты. Межклеточное вещество, его основное вещество и волокнистый остов. Связь цемента с периодонтом.
Топография различных видов цемента в однокорневых и многокорневых зубах. Питание цемента. Отличия от кости. Возрастные особенности. Гиперцементоз. Мягкие ткани зуба. Особенности строения и морфофункциональное значение пульпы зуба. Межклеточное вещество пульпы, его гистохимическая характеристика. Клетки пульпы. Особенности строения слоев пульпы. Одонтобласты, их структура и роль. Пульпа коронки и пульпа корня. Иннервация и кровоснабжение пульпы зуба. Иннервация, кровоснабжение и лимфатические сосуды пульпы.
Чувствительность дентина и пульпы. Значение пульпы в жизнедеятельности зуба. Реактивные свойства, асептическое воспаление и регенерация пульпы зуба. Дентикли. Возрастные и регрессивные изменения пульпы.
Поддерживающий аппарат зубов. Периодонт, клетки и коллагеновый остов. Циркулярная связка. Особенности расположения волокон в разных отделах периодонта. Маргинальный периодонт. Эпителиальные включения в периодонте. Кровоснабжение и иннервация периодонта. Зубная альвеола, строение и фунющональная характеристика. Особенности расположения и строения межальвеолярных и межкорневых перегородок.
Перестройка периодонта, зубных альвеол и альвеолярных частей верхней и нижней челюсти в ответ на изменения функциональной нагрузки. Десна. Зубо-десневое соединение. Десневая щель и десневой карман и ею роль в патологии. Пародонт: десна, цемент, периодонт, кость альвеолы. Его возрастные изменения и функциональная перестройка.
Развитие лица, ротовой полости и челюстей. Ротовая ямка. Первичная ротовая полость. Жаберный аппарат. Его части и производные. Жаберные карманы, щели и дуги. Развитие лица, развитие неба и разделение первичной роговой полости на окончательную роговую и носовую полости. Развитие преддверия полости рта. Развитие челюстного аппарата. Развитие языка. Пороки развития.
Развитие зубо-челюстной системы. Одонтогенез. Развитие и рост выпадающих (молочных) зубов. Образование щечно-зубной и первичной зубной пластинок. Закладка зубного зачатка. Дифференцировка зубного зачатка. Эпителиальный зубной орган, зубной сосочек, зубной мешочек. Их строение, развитие и производные. Нарушения ранних стадий развития зуба. Гистогенез зуба. Одонтобласты и их значение в образовании дентина в коронке и корне зуба. Образование радиальных и тангенциальных дентинных волокон. Плащевой и околопульпарный дентин.
Предентин. Нарушения дентиногенеза. Энамелобласты, изменение их полярности. Энамелогенез. Возникновение эмалевых призм. Обызвествление эмали. Неонатальная линия. Созревание эмали. Нарушения энамелогенеза. Развитие корня зуба. Цементобласты и их значение в образовании цемента. Формирование клеточного и бесклеточного цемента. Дифференцировка зубных сосочков. Развитие пульпы зуба. Васкуляризация и иннервация развивающегося зуба. Развитие периодонта и костной альвеолы. Прорезывание выпадающих (молочных) зубов. Теории прорезывания зубов. Нарушения прорезывания зубов. Закладка, развитие и прорезывание постоянных зубов. Смена зубов. Физиологическая и репаративная регенерация тканей зуба. Возрастные изменения зубов. Особенности развития многокорневых зубов.
Органоиды специального назначения
К органоидам специального назначения относятся микроворсинки, реснички, жгутики, миофибриллы, псевдоподии.
Реснички и жгутики –это подвижные цитоплазматические выросты, в которых находится осевая нить (или аксонема), а у основания – базальное тельце. Базальное тельце служит матрицей для организации аксонемы. Само базальное тельце состоит из 9 триплетов микротрубочек, располагается в основании реснички или жгутика и напоминает по строению центриоль.
Аксонема состоит из 9 периферических и двух центрально расположенных пар микротрубочек. Центральная пара окружена центральной оболочкой, от которой к периферическим парам в виде лучей расходятся радиальные спицы. Периферические пары микротрубочек связаны между собой белком нексином. Кроме того, от микротрубочек соседних пар отходят ручки из белка динеина, обладающего активностью АТФ-азы. Аксонема формируется путем самосборки белковых субъединиц. Матрицей для самосборки служит базальное тельце.
Микроворсинки – это цитоплазматические выросты длиной около 1 мкм. Они многократно увеличивают поверхность клетки. Каждая микроворсинка имеет внутренний каркас, образованный пучком около 40 микрофиламентов, расположенных вдоль клеточной оси и фиксирующихся на внутренней поверхности плазмолеммы.
Миофибриллы– органоиды специального назначения мышечных волокон. Они имеют вид тонких нитей, вытянутых вдоль мышечных волокон и отделяющихся друг от друга рядами вытянутых митохондрий и эндоплазматической сети. Сами миофибриллы состоят из сократительных белков актина, миозина и других.
Псевдоподии (ложноножки) –это временные цитоплазматические выпячивания у некоторых одноклеточных организмов и некоторых клеток (например, лейкоцитов) многоклеточных животных. Служат для амебоидного движения и фагоцитоза.
Цитоскелет
Цитоскелет – это трехмерный цитоплазматический комплекс волокнистых и трубчатых структур, придающий клетке определенную форму и выполняющий другие функции. Он представлен микротрубочками, микрофиламентами и промежуточными филаментами.
Микротрубочки – неветвящийся структурный компонент цитоплазмы, присутствующий практически во всех типах эукариотических клеток. Имеют диаметр 24 нм, а в длину –несколько мкм. Это динамические структуры, постоянно растущие с одного конца и деполяризующиеся с другого. Они построены из белка тубулина. Имеют вид полых цилиндров.
Их главная функция заключается в создании эластичного и устойчивого внутриклеточного каркаса, необходимого для поддержания формы клеток. Кроме того, они принимают участие в транспорте макромолекул и органоидов, обеспечивают подвижность жгутиков и ресничек. Входя в состав веретена деления, обеспечивают расхождение хромосом при делении клеток. Они могут быть факторами ориентированного движения клетки в целом.
Разрушение микротрубочек колхицином приводит к нарушениям транспорта веществ (например, к блокаде секреции) и механического переноса отдельных внутриклеточных компонентов. Кроме того, при делении клеток блокируется веретено деления.
Микрофиламенты – относятся к фибриллярным компонентам эукариотических клеток. Они располагаются в кортикальном слое цитоплазмы, прямо под плазмолеммой, в виде пучков или слоями. В среднем их длина составляет 6 нм.
В зависимости от химического состава микрофиламенты могут выполнять функции цитоскелета и участвовать в обеспечении движения. В их состав входят сократительные белки: актин, миозин, тропомиозин и др.
Основные функции микрофиламентов – это создание внутриклеточного сократительного аппарата, который обеспечивает амебоидное движение клеток, большинство движений органоидов и деление клеток.
Промежуточные филаменты(микрофибриллы) – это тонкие неветвящиеся, нередко лежащие пучками нити, которые присутствуют в клетках разных тканей и состоят из разных веществ, например, из кератина – в эпителии, десмина – в мышечных волокнах и др. Они выполняют опорно-каркасную функцию.
Включения
Включения – это непостоянные компоненты, которые могут появляться и исчезать в зависимости от функционального состояния клетки. Их классифицируют на: трофические, секреторные, экскреторные, пигментные, кристаллы. Трофические включения – это, как правило, резервные скопления гранул белка, гликогена, капельки жиров. Секреторные гранулы и пузырьки содержат биологически активные вещества и поступают по протокам желез в те или иные органы. Экскреторные включения не являются биологически активными веществами. Обычно это продукты метаболизма клеток, подлежащие удалению. Пигментные включения могут изменить цвет ткани временно или постоянно (примеры: гемоглобин, меланин, липофусцин – пигмент изнашивания, билирубин и др.). В растительных клетках встречаются включения кристаллов (чаще всего это оксалат кальция).
Узнать еще:
Немембранные органоиды
Задание 1. 37. Сравните строение и функции микроворсинок и микротрубочек. Ответ поясните.
Задание 2. 34. Объясните, где синтезируются рибосомы? Какое строение они имеют? Ответ поясните.
Задание 3. 37. В чем разница строение базальных телец и ресничек? Как они связаны? Ответ поясните.
Задание 4. 37. В чем разница строения жгутика и центриолей? Зачем нужны центриоли? Ответ поясните.
Задание 5. 34. Какое строение имеет клеточный центр? Могут ли его структуры увеличиваться в количестве? Ответ поясните.
Задание 6. 34. Какие организмы не имеют центриолей? Есть ли центриоли у бактерий? Ответ поясните.
Ответы.
Задание 1.
1. Микроворсинка — это вырост плазматической мембраны клетки, под которым имеются актиновые микрофиламенты.
2. Функции микроворсинок — увеличение площади поверхности клетки, передвижение.
3. Микротрубочки образованы из тубулина, находятся внутри клетки и представляют собой полые цилиндры.
4. Функции микротрубочек — перемещение органоидов в клетке и формирование веретена деления.
Задание 2.
1. Рибосомы синтезируются в ядрышке клетки; основа рибосом — рРНК, которая создается на основании определенной ДНК.
2. Рибосомы состоят из большой и малой субъединиц, на 50% состоят из рРНК, на 50% — из белка.
Задание 3.
1. Базальные тельца — «корни», «якоря» ресничек, которые укрепляют их в цитоплазме.
2. У ресничек внутри есть пучки микротрубочек; из них 9 дуплетов микротрубочек на периферии, один дуплет в центре; снаружи реснички покрыты плазмалеммой.
3. У базальных телец внутри по периферии расположены 9 триплетов микротрубочек.
Задание 4.
1. Жгутик — длинная структура, закреплённая в цитоплазме базальным тельцем, необходимая для передвижения клетки.
2. Центриоли — два перпендикулярных цилиндра из микротрубочек.
3. Центриоли играют важную роль в процессе деления клетки — участвуют в процессе формирования веретена деления.
Задание 5.
1. Клеточный центр — это центриоли с отходящими от них микротрубочками.
2. Да, они могут делиться, и формировать два полюса при делении материнской клетки.
3. После деления должно восстановиться изначальное число органоидов, в том числе центриолей.
Задание 6.
1. Высшие растения не имеют центриолей, как и все прокариоты.
2. Бактерии центриолей не имеют, так как для них не характерен митоз и отсутствует веретено деления, центром которого у других организмов являются центриоли.
Смотреть еще: подготовка к ЕГЭ по биологии онлайн, репетитор по биологии в Москве, онлайн тесты по биологии.
обыкновенные пили, F-пили. Химический состав, строение, функции.
В клетке насчитывается от мало до дохуя. Построены из белка пилина и представляют из себя цилиндры отходящие от клеточной оболочки. Тоньше жгутиков, расположены пертрихально или полярно. У кишечки их больше всего. Сам пилин имеет консервативные и вариабельные участки. Являются необходимым антигеном к фактору колонизации. Обеспечивают адгезию.
Обыкновенные – придают гидрофобность, обеспечивают приврепление к поверхности и участвуют в транспорте метаболитов. Через пили могут проникать фаги.
F-пили – необходимы для соединения клеток и передачи ДНК, образуют коньюгационный тонель. (каким образом?)
1. Основные типы движения прокариот (плавание, «роение», скольжение, подтягивающее движение и др.). Таксисы у прокариот.
Присущая спирохетам локализация двигательного аппарата интересна тем, что позволяет сделать вывод о возможности его работы в условиях нахождения в «закрытом» клеточными структурами состоянии. Это может служить ключом к пониманию еще одного вида движения, присущего части прокариот, — скольжения. Последнее определяют как способность организма передвигаться по твердому или полужидкому субстрату без помощи наружных локомоторных структур — жгутиков.
Способность к скольжению
обнаружена у разных групп прокариот, как одноклеточных, так и многоклеточных, имеющих нитчатое строение: некоторых микоплазм, миксобактерий, цитофаг, нитчатых серобактерий, цианобактерий и др. Скорость этого типа движения невелика: 2–11 мкм/с. Общим для всех скользящих организмов является способность к выделению слизи. Кроме того, у ряда скользящих форм в составе клеточной стенки между пептидогликановым слоем и наружной мембраной обнаружен тонкий слой, состоящий из белковых фибрилл. Например, у нитчатой цианобактерий Oscillatoria к наружной поверхности пептидогликанового слоя примыкают параллельные ряды фибрилл диаметром 5–7 нм: на 1 мкм2 поверхности приходится до 55 таких фибрилл. У нитчатых цианобактерий фибриллы формируют единую систему, непрерывно в виде спирали обволакивающую весь трихом (нить). Скольжение нитчатых форм сопровождается и одновременным их вращением, так что любая точка на поверхности трихома описывает при движении спираль. Направление вращения является видоспецифическим признаком и коррелирует с направлением хода спирали белковых фибрилл.
Механизм скользящего движения не ясен. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в клеточной стенке, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Однако анализ этой модели привел к заключению, что для обеспечения скольжения по «реактивному» механизму клетке необходимо в течение 1 с выделять такой объем слизи, который во много раз превосходит ее цитоплазматическое содержимое.
По другой гипотезе, получившей распространение в последние годы, скользящее движение связано с особенностями строения клеточной стенки подвижных безжгутиковых форм — наличием белкового слоя, состоящего из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков, с той разницей, что находятся фибриллы «внутри» клеточной стенки. У некоторых скользящих бактерий описаны структуры, весьма напоминающие базальные тела жгутиковых форм. Вращательное движение фибрилл, «запускаемое» этими структурами, приводит к появлению на поверхности клетки так называемой «бегущей волны», т. е. движущихся микроскопических выпуклостей клеточной стенки, в результате чего клетка отталкивается от твердого или вязкого субстрата. На скольжение расходуется около 5% энергии от общего объема клеточных энергетических затрат. Скользящее движение в разных группах бактерий обеспечивается энергией в форме АТФ или DmH+.
Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
Необходимость для скольжения слизи пока не ясна. Скольжение может происходить в среде подходящей консистенции без какого-либо выделения слизи. Более того, выделение больших количеств слизи, как правило, затрудняет движение клетки и приводит к потере ею подвижности. Согласно гипотезе «бегущей волны» выделение слизи не является абсолютно необходимым для скольжения, но облегчает в определенных условиях отталкивание клетки от субстрата.
Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами.
В зависимости от фактора различают:
хемотаксис (частный случай — аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. Наибольшее внимание привлекает изучение хемотаксиса, т. е. движения в определенном направлении относительно источника химического вещества. Для каждого организма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы (эффекторы). Среди последних выделяют аттрактанты
(вещества, привлекающие бактерий) и репелленты (вещества, отпугивающие бактерий).
Аттрактантами могут быть сахара, аминокислоты, витамины, нуклеотиды и другие химические молекулы; репеллентами — некоторые аминокислоты, спирты, фенолы, неорганические ионы. Аттрактантом для аэробных и репеллентом для энаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны.
Фототаксис, т. е. движение к свету или от него, свойствен прежде всего фототрофным бактериям. Способность перемещаться по силовым линиям магнитного поля Земли или магнита — магнитотаксис — обнаружен у разных бактерий, обитающих в пресной и морской воде. В клетках этих бактерий найдены непрозрачные частицы определенной геометрической формы — магнитосомы, заполненные железом в форме магнетита (Fe3O4) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4% сухого вещества бактерий. В северном полушарии такие магниточувствительные бактерии плывут в направлении северного полюса Земли, в южном — в направлении южного. У ряда бактерий обнаружен вискозитаксис — способность реагировать на изменение вязкости раствора и перемещаться в направлении ее увеличения или уменьшения.
За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы. Изучение хемотаксиса у Escherichia coli позволило обнаружить свыше 30 различных хеморецепторов, представляющих собой белки, синтезируемые независимо от присутствия индуктора или только в результате индукции. Рецептор реагирует на эффектор и передает сигнал по определенному пути, конкретный механизм которого неизвестен, на «мотор» жгутика.
У бактерий с перитрихиальным жгутикованием выявлены два вида двигательного поведения: прямолинейное движение и кувырканье, т. е. периодические и случайные изменения направления движения. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, ее прямолинейное движение, ориентированное по отношению к химическому веществу, становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
1) Скользящее движение неравномерным выделением слизи (встречается у миксобактерий, цианобактерий).
2) Плавающее передвижение с помощью жгутиков – наиболее распространенный тип движения. Жгутики – очень тонкие образования диаметром 10-20 нм, редко до 60 нм (у сложных жгутиков). Количество их варьирует от одного до тысячи.
Подвижность бактерий определяют микроскопией препаратов в «раздавленной» или «висячей» капле. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные — по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют её), либо определяют способность бактерий давать «феномен роения».
В частности, Proteus vulgaris распространяется по поверхности агара, образуя тонкий налёт (напоминающий таковой при выдохе на холодное стекло), а неподвижные штаммы протея лишены такой способности. Это явление получило название «феномен роения», а наблюдение за ним легло в основу некоторых понятий бактериальной серодиагностики.
Если бактерия находится в жидкой среде, то жгутик помогает ей плыть. Плавание — это самый быстрый способ передвижения. Причем, бактерия может неплохо управлять своим движением, меняя направление вращения базального тела: вращение базального тела по часовой стрелке толкает клетку в направлении от жгутика, а биения против часовой стрелки тянут клетку вслед за жгутиком.
А теперь представьте размахивание жгутиками на твердой поверхности, смоченной жидкостью. Бактерии будут не плыть, а расползаться в одной плоскости. Такое движение называется роением. Роение чаще бывает у бактерий, живущих в крупных колониях, — подвижные бактерии, находящиеся с краю, пытаются отодвинуться как можно дальше и основать свои собственные колонии.
Бактерии могут также создавать более короткие и просто устроенные нити, чем жгутики, — пили. Клетка может с помощью пили прикрепиться к чему-нибудь твердому, а потом подтянуться к месту крепления, разбирая эту нить, начиная от места крепления пили к клетке (рис. 2). Можно сказать, что клетка перемещается рывками. Подобный способ движения у одной клетки называется твитчинг (англ. twitch — дергать, тащить). А если так действует несколько скрепленных друг с другом бактерий, то они дружно скользят по твердой поверхности.
В оболочках клетки могут быть разнообразные белковые комплексы, например, обхватывающие клетку кольца из белков. Эти кольца крутятся, как гусеницы у гусеничного трактора, и помогают бактерии скользить по твердой поверхности. Такой способ подвижности есть у бактерии Flavobacterium johnsoniae.
У других бактерий есть белки, расположенные вдоль всей поверхности клетки. Эти белки создают продольные волны, и бактерия извивается и плывет в водной толще или скользит на твердом субстрате.
Поможем написать любую работу на аналогичную
тему
Получить выполненную работу или консультацию специалиста по вашему
учебному проекту
Узнать стоимость
Микроворсинка
ⓘ Микроворсинка
Микроворсинка — вырост эукариотической клетки, имеющий пальцевидную форму и содержащий внутри цитоскелет из актиновых микрофиламентов. Из микроворсинок состоит воротничок у клеток хоанофлагеллят и у воротничково-жгутиковых клеток губок и других многоклеточных животных. В организме человека микроворсинки имеют клетки эпителия тонкого кишечника, на которых микроворсинки формируют щеточную кайму, а также механорецепторы внутреннего уха — волосковые клетки.
Микроворсинки нередко путают с ресничками, однако они резко отличаются по строению и функциям. Реснички имеют базальное тело и цитоскелет из микротрубочек, способны к быстрым движениям кроме видоизмененных неподвижных ресничек и служат у крупных многоклеточных обычно для создания потоков жидкости или восприятия раздражителей, а у одноклеточных и мелких многоклеточных животных также для передвижения. Микроворсинки не содержат микротрубочек и способны лишь к медленным изгибаниям в кишечнике либо неподвижны.
За упорядочение актинового цитоскелета микроворсинок отвечают вспомогательные белки, взаимодействующие с актином — фимбрин, спектрин, виллин и др. Микроворсинки также содержат цитоплазматический миозин нескольких разновидностей.
Микроворсинки кишечника не путать с многоклеточными ворсинками во много раз увеличивают площадь поверхности всасывания. Кроме того, у позвоночных на их плазмалемме закреплены пищеварительные ферменты, обеспечивающие пристеночное пищеварение.
Микроворсинки внутреннего уха стереоцилии интересны тем, что образуют ряды с различной, но строго определенной в каждом ряду длиной. Вершины микроворсинок более короткого ряда соединены с более длинными микроворсинками соседнего ряда с помощью белков — протокадгеринов. Их отсутствие или разрушение может приводить к глухоте, так как они необходимы для открывания натриевых каналов на мембране волосковых клеток и, следовательно, для преобразования механической энергии звука в нервный импульс.
Хотя микроворсинки сохраняются на волосковых клетках в течение всей жизни, каждая из них постоянно обновляется за счёт тредмиллинга актиновых филаментов.
Microvilli: определение и функции — видео и стенограмма урока
Где они обитают?
Микроворсинки чаще всего встречаются в тонком кишечнике, на поверхности яйцеклеток, а также на лейкоцитах. В кишечнике они работают вместе с ворсинками, поглощая больше питательных веществ и материала, поскольку они увеличивают площадь поверхности кишечника. Как вы увидите через секунду, они очень важны для нас из-за этого факта.
Они также играют роль в яйцеклетках, поскольку помогают закрепить сперматозоиды на яйцеклетке, облегчая тем самым оплодотворение.В лейкоцитах микроворсинки снова действуют как якорная точка. Они позволяют лейкоцитам, которые мчатся по телу, хвататься за все, что им нужно в данный момент, и прилипать к ним.
Почему они так важны?
Микроворсинки чрезвычайно важны, потому что они увеличивают площадь поверхности клетки, на которой они находятся. Имея эти крошечные складки в мембране любой клетки, на которую мы смотрим, вы получаете больше от той же упаковки. Представьте на секунду, что у вас на полу лежит веревка длиной шесть футов.Он занимает шесть футов только что раскинутой недвижимости. Теперь, если вы сделаете небольшие изгибы струны от конца до конца, вы сможете значительно сократить занимаемое пространство. Вы, вероятно, могли бы даже заставить эту веревку поместиться в пространстве длиной всего один фут или меньше. Я имею в виду, посмотрите на эти браслеты выживания!
Смысл, который мы пытаемся здесь показать, заключается в том, что у вас есть только определенное количество места для чего-то, но, сложив это и обернув, вы можете вместить большое количество места в небольшое пространство.Это важно в организме, потому что большая часть того, что составляют клетки, находится только в определенном пространстве. Имея складки в мембране, вы можете увеличить количество пространства для абсорбции питательных веществ, и других материалов, но при этом вы по-прежнему занимает только то же пространство, что и раньше. Как указывалось ранее, мы видим это везде, где расположены микроворсинки, но одна из самых заметных — в тонком кишечнике.
В тонком кишечнике пища расщепляется, и наш организм использует ее для получения энергии.Позволяя мембране кишечника иметь ворсинки, а затем и микроворсинки, мы стремительно увеличиваем площадь, на которой пища может прикрепляться и поглощаться. Это позволяет получить гораздо большую площадь поверхности, чем если бы у нас не было микроворсинок. Большая площадь поверхности означает большее усвоение питательных веществ, что означает больше энергии и счастливых тел.
Итоги урока
Давайте рассмотрим. Microvilli , по сути, представляют собой плазматическую мембрану, поддерживаемую сетью микрофиламентов и цитоплазмы. Это крошечные выступы, которые существуют внутри клеток и вокруг них.Микроворсинки встречаются в различных структурах и участках организма. Одним из таких мест является тонкий кишечник, где микроворсинки и ворсинки работают над увеличением площади поверхности кишечника, что позволяет лучше усваивать жизненно важные питательные вещества для организма.
Определение Microvilli и факты о нем
- Microvilli : плазматические мембраны, поддерживаемые микрофиламентами и цитоплазмой
- Ворсинки : крошечные складки в тонкой кишке
- Поглощение питательных веществ : Если на клеточной мембране есть складки, это означает, что имеется увеличенное пространство.Это позволяет усваивать питательные вещества.
Результаты обучения
Узнайте, подготовил ли вас этот урок к следующим действиям:
- Переопишите микроворсинки и обсудите их состав
- Укажите расположение микроворсинок на теле
- Признать важность микроворсинок
Microvillus — обзор | Темы ScienceDirect
Врожденная
Болезнь включения микроворсинок (врожденная атрофия микроворсинок), хотя и встречается редко, но, по-видимому, является наиболее частой причиной врожденной трудноизлечимой водянистой диареи.Диагноз ставится на основании биопсии кишечника, демонстрирующей атрофию ворсинок, гипоплазию крипт и, при электронной микроскопии, включение микроворсинок в энтероциты. Младенцам с этим заболеванием требуется полное парентеральное питание.
Тафтинговая энтеропатия проявляется в первые недели жизни трудноизлечимой водянистой диареей, но ее начало наступает несколько позже, чем при болезни включения микроворсинок. Замечательным открытием является то, что большая часть эпителиальной поверхности содержит очаговые эпителиальные пучки (группы плотно упакованных энтероцитов в форме капли с апикальным закруглением плазматической мембраны).
Врожденная мальабсорбция глюкозы и галактозы — редкое заболевание с дефектом системы котранспортеров глюкозы и натрия, приводящее к увеличению осмотической нагрузки в просвете кишечника. Однако пораженные люди способны абсорбировать фруктозу, которая становится источником пищевых углеводов.
Врожденная хлоридная диарея — редкое аутосомно-рецессивное заболевание. Часто встречается многоводие по материнской линии. При рождении у младенца развивается быстрое обезвоживание с заметным вздутием живота.Хлорид в стуле составляет 120 ммоль / л –1 , имеется метаболический алкалоз. При врожденной натриевой диарее клинические проявления аналогичны таковым при врожденной хлоридной диарее, за исключением того факта, что уровень натрия в стуле достигает 145 мЭкв / л -1 , а pH стула является щелочным, в отличие от того, что наблюдается при врожденной хлоридной диарее .
Первичная мальабсорбция желчных кислот — еще один чрезвычайно редкий дефект транспорта в дистальной части подвздошной кишки. Неизлечимая диарея начинается рано после рождения, за ней следует задержка развития и мальабсорбция жира.
Гипобеталипопротеинемия клинически неотличима от абеталипопротеинемии (синдром Бассена – Корнцвейга) и проявляется стеатореей, задержкой роста, гиполипидемией и акантоцитозом, которые наблюдаются с младенчества. Дефект — это неспособность образовывать мицеллы, важный механизм переноса жира в энтероциты. Болезнь задержки хиломикронов может проявляться сходным образом со стеатореей и задержкой развития. Дети с кишечной лимфангиэктазией могут страдать от стеартореи, энтеропатии с потерей белка и лимфопении.
Дефицит фермента дисахаридазы может быть диагностирован с помощью дыхательного водородного теста и подтвержден биопсией кишечника, демонстрирующей нормальную гистологию и почти полное отсутствие дисахаридазной активности. Врожденный дефицит сахаразы-изомальтазы становится очевидным при добавлении в рацион сахарозы или крахмала. Врожденная лактазная недостаточность — редкое состояние. Сообщалось о врожденной недостаточности глюкоамилазы у детей с хронической диареей, которая усугублялась при пероральном введении крахмала.
Нарушение переваривания жиров, как правило, вызывает частую дефекацию.Стул имеет неприятный запах, объемный, жирный и плавает на поверхности туалетной воды. Это одна из самых распространенных причин, по которой дети не могут развиваться. Тест на хлорид пота подтверждает диагноз муковисцидоза у некоторых из этих детей. Однако внешнесекреторная недостаточность поджелудочной железы может возникать и при других заболеваниях.
Ячейка. Больше информации. Микровиллы. Атлас гистологии растений и животных
Микроворсинки — это короткие тонкие выступы, обычно обнаруживаемые на свободных поверхностях некоторых дифференцированных клеток, чаще встречаются в эпителиальных клетках (Рисунки 1 и 2).Они представляют собой нитевидные структуры длиной от 1 до 2 мкм и толщиной около 100 нм. Внутри каждая микроворсинка содержит несколько десятков актиновых нитей, ориентированных параллельно длинной оси. Микроворсинки обычно плотные и плотно упакованы, как бордюр. На виде сверху микроворсинки пространственно организованы в виде шестиугольников.
Рис. 1. Эпителий тонкой кишки при световой микроскопии (изображение слева) и при сканирующей электронной микроскопии (изображение справа), демонстрирующих микроворсинки на свободной поверхности клеток.
Рис. 2. Изображение поверхности эпителия тонкой кишки, полученное с помощью просвечивающей электронной микроскопии. Терминальная сеть актиновых филаментов представляет собой более темную полосу, наблюдаемую у основания микроворсинок.
Есть много типов клеток, несущих микроворсинки, но они чаще обнаруживаются в эпителиальных клетках, таких как энтероциты кишечника, извитый эпителий протоков почек и придаток яичка. Микроворсинки также можно найти в некоторых специализированных сенсорных клетках, таких как нейроны обонятельных рецепторов, орган рецепторных клеток Кортиева, клетки плаценты и в движущихся клетках.Хотя микроворсинки демонстрируют сходную внешнюю морфологию в разных типах клеток, они содержат различный молекулярный состав и обнаруживают некоторые различия во внутренней структуре. Ниже мы рассмотрим особенности микроворсинок энтероцитов эпителия тонкой кишки.
1. Формация
Микроворсинки образуются после того, как актиновые филаменты связываются с плотной пластиной, находящейся на цитозольной поверхности плазматической мембраны. Также необходимы белки для сбора актиновых филаментов в пучки и их стабилизации.Похоже, это последовательный процесс. Во-первых, ворсин облегчает формирование пучков актиновых нитей и стабилизирует их дистальные концы, подавляя рост нитей, поэтому ворсинки могут быть важны для определения длины микроворсинок. Эзрин, в меньшем количестве, помогает соединить актиновые филаменты пучка с плазматической мембраной. Вторая фаза формирования микроворсинок включает достижение конечной длины и распределение по поверхности клетки в виде шестиугольников. Фимбрин и спин могут способствовать этим особенностям.Боковые связи протокадгеринов между актиновыми филаментами различных микроворсинок также происходят во время этой второй стадии. Важно отметить, что при формировании микроворсинок необходимо большое количество мембранных и мембранных белков, обеспечиваемых интенсивным экзоцитозом. Отклонения плазматической мембраны, по одному на каждую микроворсинку, являются следствием полимеризации актиновых филаментов.
2. Строение
Микроворсинки состоят из 6 основных белков: актина, фимбрина, виллина, миозина (Myo1A), кальмодулина и спектрина (неэритроцитарного) (рис. 3).Каркас микровыступов представляет собой пучок из 30-40 актиновых филаментов, параллельных длинной оси и их минусовые концы к вершине микровысоких. Фимбрин и виллин связывают вместе актиновые филаменты, тогда как Myo1A и кальмодулин образуют связи между пучком и мембраной. Положительный конец актиновых филаментов простирается в цитоплазму, и там актиновые филаменты различных микроворсинок перемешиваются между собой и образуют паутину с гексагональным узором. Эта сеть известна как конечная сеть, и она распространяется по периферии клетки под микроворсинками.Терминальная паутина содержит большое количество неэритроцитарного спектрина.
Рис. 3. Молекулярная организация микроворсинок (адаптировано из Crawley et al., 2014)
Каждая микровышка стабильна и не перемещается, но ее белковый каркас постоянно обновляется за счет обмена актина и других белков каркаса между микровысотой и цитозолем. Это означает, что существует баланс между входящими и исходящими белками. Подсчитано, что каждые 20 минут каркас каждой микровысоты полностью обновляется.Микроворсинки могут быть полностью дезорганизованы и исчезать после увеличения концентрации кальция, например, при клеточном стрессе. Кальций изменяет поведение ворсинок от стабилизации до разъединения актиновых нитей. Микроворсинки исчезают в клетках, вступающих в митоз. Терминальная перемычка также имеет пластиковую конструкцию.
3. Функция
Обмен веществ между полостями или внешней средой и тканями является одной из основных функций эпителия, такого как эпителий кишечника и почек.В основном это происходит на апикальной (свободной) поверхности эпителиальных клеток, где расположены большинство молекулярных переносчиков, ионных насосов и происходят эндоцитозные процессы. Чем больше поверхность клетки, тем больше места для размещения молекулярных механизмов обмена. Микроворсинки представляют собой шиповидные выступы, которые значительно увеличивают поверхность плазматической мембраны и, следовательно, содержание мембранного белка. Это особенно важно для абсорбирующих и секреторных клеток эпителия. Микроворсинки могут увеличивать поверхность клетки до 100 раз по сравнению с плоской мембраной.Микроворсинки энтероцитов несут множество ферментов, которые участвуют не в молекулярном обмене, а в пищеварении.
Микроворсинки могут влиять на молекулярные пути трансдукции. Их мембраны работают как относительно изолированные домены с определенным набором молекул, таких как переносчики глюкозы, ионные насосы и рецепторы. Длина микроворсинок хорошо подходит для выполнения локальной трансдукции молекулярных путей независимо от цитозоля. Кроме того, актиновые филаменты и каркас структурных белков образуют плотную сеть молекул, которая каким-то образом фильтрует или контролирует молекулы, приходящие или покидающие внутреннюю часть каждой микровысоты.Этот каркас также может служить временным хранилищем кальция.
Высокая плотность микроворсинок создает физический барьер, защищающий от патогенов. Это важно для эпителия кишечника. Кроме того, большое количество мембраны, хранящейся в микроворсинках, является резервуаром, который может противостоять гипертоническим воздействиям и предотвращать разрыв плазматической мембраны.
Стереоцилии — это специализированные микроворсинки, выполняющие сенсорные функции. Несмотря на название стереоцилии, на самом деле они представляют собой модифицированные микроворсинки, преобразованные в сенсорные структуры, и некоторые авторы предлагают вместо этого название эстереомикровеллы.Их можно найти в эпителии придатка яичка и во внутреннем ухе, работая как механорецепторы, которые ощущают движение окружающей жидкости. Стероцилии внутреннего уха млекопитающих обнаруживаются как часть кортиевского органа. Они имеют длину от 10 до 50 мкм и содержат внутри более 3000 актиновых нитей. Звуковые волны воздуха преобразуются в жидкие волны в ухе, которые стереоцилии воспринимают и преобразуют в электрические сигналы, которые проходят через слуховой нерв в мозг. Стереоцилии похожи на антенны, принимающие внешние сигналы, которые преобразуются в электрическую информацию.
Некоторые микровысоты специализируются на восприятии света. Фоторецепторы — это светочувствительные клетки, обнаруженные в органах зрения, таких как сетчатка. Клеточные сенсорные структуры на самом деле представляют собой модифицированные реснички или микровыступы. Фоторецепторы с измененной микровысотой для обнаружения света можно найти в зрительных структурах беспозвоночных. Эти фоторецепторы содержат микровысоты, расположенные в так называемых рабдомерах. Каждая микровысадка содержит визуальные пигменты, которые могут реагировать на низкую интенсивность света и эффективны при высокой интенсивности света.Микровысоты организуются, образуя так называемые рабдомеры, а молекулярный путь трансдукции делает световой ответ, основанный на микроворсинах, более чувствительным, чем светочувствительные системы ресничек позвоночных.
Предполагается, что большое количество внеклеточных пузырьков высвобождается из микроворсинок. Кончики микроворсинок энтероцитов способны выпускать пузырьки в окружающую среду. Связь между плазматической мембраной и актин-миозиновой сетью может способствовать образованию этих везикул, перетаскивая мембрану к дистальной части каждого микровыступа, где небольшие участки мембраны трансформируются в пузырьки.Эти внеклеточные везикулы содержат ферменты и обогащены щелочной фосфатазой.
Библиография
Коричневый J W, McKnight C J . 2010. Молекулярная модель микровиллярного цитоскелета и организация щеточной каймы. PLoS One. 5: e940.
Crawley SW, Mooseker MS, Tyska MJ . 2014. Формирование щеточной каймы кишечника. J Cell Biol. 207: 441-451.
Файн Дж. Л., Харди Р., Лафлин С. Б. . 2010 г.Фототрансдукция и эволюция фоторецепторов. Curr Biol. 20: R114-R124.
Ланге К . 2011. Фундаментальная роль микроворсинок в основных функциях дифференцированных клеток: Схема универсальной регулирующей и сигнальной системы на периферии клетки. J. Cell Physiol. 226: 896-92
МакКоннелл Р. Э., Хиггинботэм Дж. Н., Шифрин-младший Д. А., Табб Д. Л., Коффи Р. Дж., Тайска М. Дж. . 2009. Микроворсинки энтероцитов — органеллы, образующие пузырьки. J Cell Biol.185: 1285-129.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A . 2015. Структура, регуляция и функциональное разнообразие микроворсинок апикального домена эпителиальных клеток. Ежегодный обзор клеточной биологии и биологии развития. 31: 593-621.
Цитоскелет, жгутики и реснички, а также плазматическая мембрана
Если бы вы удалили все органеллы из клетки, оставались бы только плазматическая мембрана и цитоплазма? Нет. Внутри цитоплазмы все еще будут ионы и органические молекулы, а также сеть белковых волокон, которая помогает поддерживать форму клетки, закрепляет определенные органеллы в определенных положениях, позволяет цитоплазме и везикулам перемещаться внутри клетки и дает возможность одноклеточные организмы передвигаться самостоятельно.В совокупности эта сеть белковых волокон известна как цитоскелет. Внутри цитоскелета есть три типа волокон: микрофиламенты, также известные как актиновые филаменты, промежуточные филаменты и микротрубочки (рис. 1).
Рис. 1. Микрофиламенты, промежуточные волокна и микротрубочки составляют цитоскелет клетки.
Микрофиламенты — самые тонкие из волокон цитоскелета, они участвуют в перемещении клеточных компонентов, например, во время деления клеток. Они также поддерживают структуру микроворсинок, обширную складку плазматической мембраны, обнаруженную в клетках, предназначенных для абсорбции.Эти компоненты также распространены в мышечных клетках и отвечают за сокращение мышечных клеток.
Промежуточные филаменты имеют средний диаметр и выполняют структурные функции, такие как поддержание формы клетки и закрепление органелл. Кератин, соединение, укрепляющее волосы и ногти, образует промежуточные волокна одного типа.
Микротрубочки — самые толстые из волокон цитоскелета. Это полые трубки, которые быстро растворяются и восстанавливаются. Микротрубочки направляют движение органелл и представляют собой структуры, которые притягивают хромосомы к своим полюсам во время деления клетки.Они также являются структурными компонентами жгутиков и ресничек. В ресничках и жгутиках микротрубочки организованы в виде круга из девяти двойных микротрубочек снаружи и двух микротрубочек в центре.
Центросома — это область около ядра клеток животных, которая функционирует как центр организации микротрубочек. Он содержит пару центриолей, две структуры, которые лежат перпендикулярно друг другу. Каждая центриоль представляет собой цилиндр из девяти троек микротрубочек. Центросома реплицируется до деления клетки, и центриоли играют роль в притяжении дублированных хромосом к противоположным концам делящейся клетки.Однако точная функция центриолей в делении клеток не ясна, так как клетки, у которых удалены центриоли, все еще могут делиться, а клетки растений, у которых нет центриолей, способны к делению клеток.
Жгутики и реснички
Жгутики (единичное число = flagellum ) представляют собой длинные волосовидные структуры, которые отходят от плазматической мембраны и используются для перемещения всей клетки (например, сперматозоидов, Euglena ). Когда присутствует, клетка имеет только один жгутик или несколько жгутиков.Однако, когда присутствуют реснички (singular = cilium ), их много, и они проходят по всей поверхности плазматической мембраны. Это короткие, похожие на волосы структуры, которые используются для перемещения целых клеток (например, парамеций) или перемещения веществ по внешней поверхности клетки (например, ресничек клеток, выстилающих фаллопиевы трубы, которые перемещают яйцеклетку к матке, или реснички, выстилающие клетки дыхательных путей, которые перемещают твердые частицы в горло, в которые попала слизь).
Плазменная мембрана
Подобно прокариотам, эукариотические клетки имеют плазматическую мембрану (рис. 2), состоящую из фосфолипидного бислоя со встроенными белками, которые отделяют внутреннее содержимое клетки от окружающей среды. Фосфолипид — это молекула липида, состоящая из двух цепей жирных кислот и фосфатной группы. Плазматическая мембрана регулирует прохождение некоторых веществ, таких как органические молекулы, ионы и вода, предотвращая прохождение одних для поддержания внутренних условий, при этом активно вводя или удаляя другие.Другие соединения пассивно перемещаются через мембрану.
Рис. 2. Плазматическая мембрана представляет собой бислой фосфолипидов со встроенными белками. Есть и другие компоненты, такие как холестерин и углеводы, которые могут быть обнаружены в мембране в дополнение к фосфолипидам и белку.
Плазматические мембраны клеток, которые специализируются на абсорбции, сложены в виде пальцевидных выступов, называемых микроворсинками (единственное число = микроворсинки). Эта складка увеличивает площадь поверхности плазматической мембраны.Такие клетки обычно выстилают тонкий кишечник — орган, который поглощает питательные вещества из переваренной пищи. Это отличный пример соответствия формы функциям конструкции. Люди с глютеновой болезнью имеют иммунный ответ на глютен, белок, содержащийся в пшенице, ячмене и ржи. Иммунный ответ повреждает микроворсинки, и, таким образом, пораженные люди не могут усваивать питательные вещества. Это приводит к недоеданию, спазмам и диарее. Пациенты, страдающие глютеновой болезнью, должны соблюдать безглютеновую диету.
У вас была идея улучшить этот контент? Нам очень понравится ваш вклад.
ответов
Какая субклеточная органелла отвечает за привлечение основного красителя? Шероховатая эндоплазматическая сеть.
# 115 Желчный пузырь, H&E
Почему микроворсинки видны не на всех клетках, выстилающих просвет? В зависимости от ориентации секции некоторые сотовые компоненты могут быть видны не во всех ячейках.
ВОПРОСОВ
1.Что такое серийные разделы и почему они важны? Непрерывный ряд секций, которые показывают структуры в трех измерениях. Серийные срезы важны для визуализации трехмерной структуры ткани, чтобы отличить артефакт от патологии.
2. Какая связь между гетерохроматином и синтетической активностью ДНК? Гетерохроматин является транскрипционно неактивной ДНК. Эухроматическое ядро относительно неокрашено, потому что генетический материал считывается.
3. Где находится сайт синтеза рибосомальной РНК? Ядрышко.
4. Каковы функции митохондриальных крист? Сайт ферментов, связанных с производством АТФ.
5. Каково функциональное значение комплекса Гольджи? Сайт посттрансляционных модификаций, упаковки и сортировки белков.
6. Сколько мембран составляет ядерная оболочка? Два. Стенка митохондрии? Два.
7. Каковы цитологические и функциональные различия между ресничками и микроворсинками? Реснички представляют собой структуры на основе подвижных микротрубочек; микроворсинки представляют собой неподвижные структуры на основе актина.
8. Знать структурные характеристики и функциональное значение следующих органелл и включений: ядро, ядрышко, рибосомы, эндоплазматический ретикулум (два типа), митохондрии, аппарат Гольджи, лизосомы, микротрубочки, реснички, микроворсинки, гликоген, липид, пероксисомы.Какие из них требуют для идентификации чего-то большего, чем просто световая микроскопия? Какие дополнительные техники можно использовать?
Сдвиг жидкости вызывает образование микроворсинок за счет механочувствительной активации TRPV6
Конструкция и изготовление устройства
Микрожидкостное устройство состоит из двух микрофлюидных каналов, имитирующих кровообращение матери (верхний слой) и плода (нижний слой), разделенных мембраной ВК. (Толщина 10 мкм, Asahi Techno Glass, Сидзуока, Япония).Размер поперечного сечения микроканалов составлял 2 мм (ширина) × 200 мкм (высота), а длина канала составляла 20 мм для канала плода и 15 мм для канала матери. Микрожидкостные каналы были приготовлены путем монтажа реплики в полидиметилсилоксане (PDMS; SYLPOT 184, Dow Corning Toray, Токио, Япония) с соотношением сшивающего агента 10: 1 (мас. / Мас.) С использованием SU-8 (SU-8 100, MicroChem Corp., MA) мастер, изготовленный стандартными фотолитографическими методами. После термического отверждения узорчатых микроканалов структура материнской камеры была введена в центр материнского канала с помощью биопсийного штампа ( φ = 4 мм).Затем мембрану VC (части 7 × 7 мм) собирали с каналами PDMS плода и матери, чтобы полностью покрыть материнскую камеру. Оба слоя PDMS обрабатывали плазмой O 2 (SAMCO) и инкубировали в течение 90 мин при 75 ° C для облегчения постоянного связывания.
Клеточные культуры и эксперименты по протоку
Трофобластные клетки BeWo, полученные из хориокарциномы человека 48,51 , были предоставлены RIKEN BRC через Национальный проект биоресурсов MEXT, Сайтама, Япония.Клетки культивировали в питательной смеси Ham’s F12 (Sigma, St Louis, MO) с добавлением 10% фетальной бычьей сыворотки и 50 мкг мл -1 сульфата канамицина (Sigma). HVT (ScienCell, Карлсбад, Калифорния) поддерживали в среде трофобласта, дополненной фетальной бычьей сывороткой и факторами роста трофобласта (ScienCell), и использовали для экспериментов в пассаже 3. Перед посевом на устройство микроканалы заполняли культуральной средой для регидратации VC мембрана на 30 мин. Клетки BeWo собирали с использованием 0.05% трипсин / ЭДТА и 20 мкл клеточной суспензии (3,0 × 10 4 клеток) добавляли в материнскую камеру. Затем материнскую камеру закрывали тонкой мембраной PDMS, и устройство инкубировали в инкубаторе CO 2 , чтобы обеспечить адгезию клеток на мембране VC. Через 1-2 часа культивирования устройство было подключено к перистальтическому насосу (GILSON, Middleton, WI) для контроля скорости потока. Все культуры клеток были выполнены при 37 ° C в увлажненном инкубаторе с 5% CO 2 . На основе компьютерного моделирования скорости сдвига с помощью COMSOL Multiphysics (COMSOL AB, Стокгольм, Швеция) (дополнительный рис.10), FSS в материнской камере оценивается в диапазоне от 0,001 (в центре камеры; низкий FSS) до 0,12 дин см -2 (на входе или выходе камеры; высокий FSS; когда скорость потока составляет 5 мкл мин (-1 ), следуя уравнению: τ = μ × δu / δy , где μ — вязкость среды для культивирования клеток со значением вязкости 9,5 × 10 −4 Па · с при 37 ° C, и δu / δy — скорость сдвига.Мы подтвердили, что ночная культура в статических условиях в нашем устройстве не вызывала серьезного недостатка кислорода или питательных веществ в клетках BeWo (дополнительный рис. 11).
РНК-интерференция
Клетки BeWo выращивали до 50% конфлюэнтности и инкубировали с реагентом для трансфекции DharmaFect1 (2 мкл в 1 мл культуральной среды) в комплексе с 25 нМ siGENOME SMARTpool против TRPV6 человека (5′-GGAAACAGCGCUACACAUA-3 ‘, 5′-GGAAACAGCGCUACACAUA-3’, 5 ‘ -GGACAAAGACUCAGUGGAA-3 ‘, 5′-UGAACAAGUUGCUCAAGUA-3′, 5’-CCAUAUAUCUGCUGUACAU-3 ‘), TRPV2 (5′-GGUAAGACGUGCCUGAUGA-3′, 5’-GAUCCUGAC-3 ‘, 5′-GAUCCUGUCU-3′, 5’-GAUCCUGU ‘ ′ -GCGAGACCGUCAACAGUGU-3 ′) или 25 нМ siGENOME Non-targeting siRNA Pool # 1 (5′-UAGCGACUAAACACAUCAA-3 ′, 5′-UAAGGCUAUGAAGAGAUAC-3 ′, 5′-AUGAUUGGCCU ) (Дхармакон, Thermo Scientific, Лафайет, Колорадо, США).В некоторых экспериментах олигонуклеотид миРНК TRPV6 против единственной целевой последовательности (# 1: 5’-GGACAAAGACUCAGUGGAA-3 ‘, # 2: 5′-UGAACAAGUUGCUCAAGUA-3’, соответственно) использовали для подтверждения специфичности действия миРНК. Олигонажи TRPV6 siRNA были синтезированы Sigma-Aldrich (Сент-Луис, Миссури, США). Через 7–12 ч среду для трансфекции заменяли свежей. Клетки, трансфицированные миРНК, культивировали в течение ночи, а затем высевали на устройство.
Иммунофлуоресценция
клеток BeWo засевали в устройство на 3.0 × 10 4 клеток на материнскую камеру и культивировали в течение ночи с перфузией среды или без нее. В некоторых экспериментах клетки BeWo, выросшие до субконфлюента на чашке со стеклянным дном, покрытой коллагеном типа I (IWAKI, Токио, Япония), инкубировали в течение ночи с капсаицином (Sigma). Клетки промывали PBS и фиксировали 4% параформальдегидом в PBS (4% PFA / PBS) в течение 20 мин при комнатной температуре. После фиксации клетки пермеабилизировали 0,2 или 0,01% Triton X-100 в течение 5-10 минут и инкубировали с 5% обезжиренным молоком (BD Difco, Sparks, MD, UAS) в PBS в течение 20 минут, чтобы заблокировать неспецифическое связывание антитело.Эти препараты затем инкубировали при 4 ° C в течение ночи с первичными антителами: мышиным моноклональным антителом против эзрина (1: 300, ab4069, Abcam, Кембридж, Массачусетс, США), мышиным моноклональным антителом против переносчика глюкозы GLUT1 (1: 300, clone SPM498, ab40084, Abcam) или фаллоидин, конъюгированный с AlexaFluor 568 (1: 300, A12380, Molecular Probes, Юджин, Орегон, США). За инкубацией первичных антител следовали 1 час инкубации с конъюгированными с Alexa Fluor488 вторичными антителами козы против IgG мыши (1: 300, A11017, Molecular Probes) и 1 мкг мл -1 4 ‘, 6-диамидино-2- фенилиндол (Sigma) в 5% обезжиренном молоке / PBS.Окрашенные образцы помещали в среду Fluoromount / Plus (Diagnostic BioSystems, Pleasanton, CA, USA) для подавления фотообесцвечивания. Поперечные и совмещенные изображения получали с помощью конфокальной лазерной сканирующей микроскопии LSM780 и программного обеспечения для визуализации ZEN (Carl Zeiss Microscopy, Йена, Германия).
ОТ-ПЦР и анализ экспрессии генов TaqMan
Тотальную РНК получали с использованием набора PureLink RNA Mini Kit (Life Technologies, Карлсбад, Калифорния, США). Комплементарную ДНК первой цепи синтезировали из 1 мкг (для ОТ-ПЦР) или 100 нг (для анализа TaqMan) общей РНК с использованием обратной транскриптазы SuperScript III (Life Technologies) и праймеров Oligo-dT (Life Technologies) для ОТ-ПЦР или случайным образом. гексамеры (New England BioLabs Inc., Ипсвич, Массачусетс, США) для анализа TaqMan. Последовательности праймеров, используемые для анализов RT-PCR, были следующими: прямой праймер глицеральдегид-3-фосфат-дегидрогеназы человека: 5′-ACCACAGTCCATGCCATCAC-3 ‘; обратный праймер глицеральдегид-3-фосфат-дегидрогеназы человека: 5’-TCCACCACCCTGTTGCTGTA-3 ‘; прямой праймер TRPV6 человека: 5’-CTCTGCCTATGGAGCAAGTTCTGC-3 ‘; Обратный праймер TRPV6 человека: 5’-GAGAGTCGAGGTCAGTGGTCC-3 ‘. Пары праймеров, используемые для TRPV1, TRPV2, TRPV3, TRPV4 и TRPV5 человека, были, как сообщалось ранее, 52 .Исходное изображение электрофореза ДНК в геле показано на дополнительном рисунке 12. Количественная ОТ-ПЦР в реальном времени была проведена с помощью системы ПЦР в реальном времени StepOne (Applied Biosystems / Life Technologies) с использованием TaqMan Fast Advanced Master Mix и TaqMan Gene Expression Assays. ( SLC2A1 , Hs00892681_m1 и RN18S1 , Hs03928985_g1 в качестве внутреннего контроля) в соответствии с инструкциями производителя. Реакции проводили в трех повторностях и анализировали сравнительным методом C T .
Поглощение и транспорт 2-NBDG в канал плода
клеток BeWo (3,0 × 10 4 клеток на материнскую камеру) высевали на устройство и культивировали в течение 20 часов с перфузией среды или без нее (2 мкл мин. -1 ). Фетальные и материнские каналы промывали сбалансированным солевым раствором (BSS; 136 мМ NaCl, 5 мМ KCl, 1 мМ CaCl 2 , 1 мМ MgCl 2 и 18 мМ HEPES, pH 7,4), а материнский канал, включая камеру часть осторожно перфузировали 2 мМ 2-NBDG (Институт пептидов, Осака, Япония).Затем устройство инкубировали в инкубаторе CO 2 в течение 20 минут без потока жидкости, чтобы обеспечить поглощение глюкозы клетками. После инкубации буфер BSS в фетальном канале извлекали и разбавляли до объема 100 мкл буфером BSS. Флуоресценцию в этом буфере измеряли для определения переноса глюкозы в канал плода. Для количественной оценки поглощения 2-NBDG клетками BeWo клетки тщательно промывали BSS и лизировали буфером для лизиса (1% TritonX-100, 50 мМ Tris-HCl (pH 8.0), 150 мМ NaCl и 1 мМ ЭДТА). Лизат извлекали и разбавляли до 50 мкл буфером для лизиса. Флуоресценцию измеряли с помощью флуороспектрометра Nanodrop 3300 (Thermo Scientific, Уолтман, Массачусетс, США).
Сканирующая электронная микроскопия
клеток BeWo (3,0 × 10 4 клеток на материнскую камеру) высевали на устройство, культивировали в течение 20 часов с перфузией среды или без нее (5 мкл мин. -1 ), а затем фиксировали 2 % глутарового альдегида (NACALAI TESQUE, Киото, Япония) в течение 1 ч при комнатной температуре.В некоторых экспериментах клетки культивировали с перфузией среды в присутствии хелаторов кальция, EGTA (Sigma) и BAPTA-AM (Sigma). Фиксированные образцы обезвоживали серией разбавлений этанола (Wako Pure Chemicals, Токио, Япония), заменяли т -бутилового спирта (Wako), сушили вымораживанием и подвергали осмиевому проводящему металлическому покрытию с помощью Plasma Coater HPC-30. W (Shinku Device, Мито, Япония). Наблюдение с помощью SEM выполняли с использованием FE-SEM (сканирующая электронная микроскопия с полевой эмиссией) SU8000 (Hitachi, Tokyo, Japan) при 5 кВ.
Измерение длины микроворсинок
СЭМ-изображения микроворсинок на поверхности клеток получали с помощью FE-SEM SU8000 (Hitachi) при 5 кВ. Морфологический анализ был выполнен с использованием ImageJ 1.48v и плагина Analyze Skelton 53 . Чтобы выделить сигналы микроворсинок, мы сначала применили порог интенсивности к отдельным изображениям. Затем с помощью команды «Скелетонизировать» были созданы каркасные изображения микроворсинок. Расстояние между отдельными конечными точками и точками ветвления скелетонизированных микроворсинок рассчитывали с помощью Analyze Skelton (2D / 3D).Чтобы избежать переоценки, мы приняли минимальную длину ветви за минимальную расчетную длину микроворсинок. Поскольку в зоне с низким FSS наблюдалось обильное образование микроворсинок, было трудно измерить длину отдельной микроворсинки. Таким образом, общая длина микроворсинок / поля была рассчитана путем суммирования длины отдельных микроворсинок.
Иммуноэлектронная микроскопия
Клетки иммуноокрашивали мышиным моноклональным антителом против GLUT1 (1: 300, клон SPM498, ab40084, Abcam) и антимышиным IgG, конъюгированным с частицами нанозолота (1:50, предоставлено Tokai Electron Microscopy, Nagoya, Япония), а затем фиксировали 2% глутаровым альдегидом в 0.1 М фосфатный буфер (pH 7,4) при 4 ° C в течение ночи. Подготовка ультратонких срезов и визуализация были выполнены с помощью Tokai Electron Microscopy. Вкратце, фиксированные образцы обрабатывали составом GoldEnhance EM (Nanoprobes, Яфанк, Нью-Йорк, США) для усиления сигналов золотых частиц и пост-фиксировали 2% тетроксидом осмия при 4 ° C в течение ночи. После дегидратации серией разбавлений этанолом образцы пропитывали пропиленоксидом, залитым в смолу (Quetol-812, Nisshin EM, Tokyo, Japan).Из блоков делали ультратонкие срезы при 90 нм алмазным ножом с использованием ультрамикротома (Ultracut UCT, Leica, Mannheim, Germany) и окрашивали уранилацетатом с последующим окрашиванием раствором свинца (Sigma-Aldrich). Образцы наблюдали с помощью просвечивающей электронной микроскопии (JEM-1400Plus, JEOL Ltd, Токио, Япония) при ускоряющем напряжении 80 кВ. Цифровые изображения были получены с помощью камеры устройства с зарядовой связью (Veleta, Olympus Soft Imaging Solutions GmbH, Мюнстер, Германия).
Ca
2+ визуализация
Ca 2+ визуализация была выполнена с материнским каналом PDMS, связанным со стеклянной подложкой (покрытой коллагеном типа I), вместо многослойного микроканала PDMS, чтобы избежать автофлуоресценции от мембраны VC.Клетки BeWo (3,0 × 10 4 клеток на материнскую камеру) высевали на устройство и оставляли для прикрепления в течение 3–6 часов при 37 ° C, инкубировали в течение 1 часа с 2 мкМ кальциевого индикатора Fura-2-AM (Life Technologies ) и трижды осторожно промыли свежей средой. Флуоресцентные изображения получали под инвертированным микроскопом (IX71, Olympus, Токио, Япония). Динамику интенсивности флуоресценции в клетках контролировали с помощью программного обеспечения Aquacosmos и камеры CCD (ORCA-R2, Hamamatsu Photonics, Сидзуока, Япония).
Иммуноблоттинг
Клетки, культивированные в области камеры устройства, лизировали 100 мкл 1 × SDS буфера для образцов (50 мМ трис-HCl (pH 6,8), 2% SDS, 50 мМ дитиотреитол и 10% глицерин). . Образцы обрабатывали ультразвуком, кипятили, разделяли электрофорезом в SDS-полиакриламидном геле с использованием 10% акриламидного геля и переносили на нитроцеллюлозную мембрану (Bio-Rad, Hercules, Калифорния, США). Мембраны предварительно инкубировали с 3,2% обезжиренным молоком в трис-буферном солевом растворе (20 мМ Трис, pH 7,4 и 150 мМ NaCl) в течение 30 минут и инкубировали при 4 ° C в течение ночи с первичным антителом, которое включало мышиное моноклональное антитело против эзрина (1: 200, ab4069, Abcam), кроличьи поликлональные антитела против эзрина (фосфо-T567) (1: 1000, ab47293, Abcam), кроличьи антитела против Akt (1: 1000, # 9272, Cell Signaling Technology, Беверли, Массачусетс, UAS ), кроличьи моноклональные антитела против фосфо-Akt (Ser473) (1: 1000, # 4060, Cell Signaling Technology), кроличьи поликлональные антитела против TRPV6 (1: 100, sc-28763, Santa Cruz Biotech, Santa Cruz, CA, США) или мышиное моноклональное антитело против β-актина (1: 5,000, клон AC15, A1978, Sigma-Aldrich).Блоты промывали трис-буферным физиологическим раствором, содержащим 0,05% Твин 20, и инкубировали с антимышиными антителами, конъюгированными с пероксидазой хрена (1: 5,000, NA9310, GE Healthcare Bio-Sciences, Упсала, Швеция) или антителами против кроликов (1: 5,000, NA9340, GE Healthcare) вторичные антитела IgG. Обнаружение антител выполняли с помощью ImmunoStar Zeta (Wako). Исходные изображения иммуноблотов показаны на дополнительном рис. 13.
Кишечные ворсинки | BioNinja
Понимание:
• Структура клеток эпителия ворсинок адаптирована к поглощению пищи
После того, как переваренная пища прошла через желудок, она попадает в тонкий кишечник для всасывания в кровь
- Тонкая кишка также высвобождает пищеварительные ферменты для обеспечения полного гидролиза молекул пищи
Внутренняя эпителиальная выстилка тонкой кишки Кишечник сильно сложен в виде пальцевидных выступов, называемых ворсинками (единственное число: ворсинки)
- Многие ворсинки будут выступать в просвет кишечника, значительно увеличивая доступную площадь поверхности для абсорбции материала
Характеристики ворсинок
Кишечник ворсинки содержат несколько структурных особенностей, которые способствуют всасыванию продуктов пищеварения:
- M icrovilli — взъерошивание эпителиальной мембраны дополнительно увеличивает площадь поверхности
- R ich кровоснабжение — плотная капиллярная сеть быстро всасывается товаров 900 40
- S однослойный эпителий — Минимизирует расстояние диффузии между просветом и кровью
- L acteals — Поглощает липиды из кишечника в лимфатическую систему
- I crypts кишечные железы — Экстракты Либеркуна) выделяют пищеварительные соки
- M эмбрановые белки — Облегчает транспортировку переваренных материалов в эпителиальные клетки
Мнемоника: MR SLIM
Особенности кишечных ворсинок
Навык:
• Идентификация клеток эпителия ворсинок, которые поглощают переваренную пищу, по электронным микрофотографиям
Эпителиальная выстилка ворсинок содержит несколько структурных особенностей, которые оптимизируют ее способность абсорбировать переваренные материалы:
Плотные соединения
- Окклюзионные ассоциации между плазматической мембраной двух соседних клеток, создавая непроницаемый барьер
- Они сохраняют пищеварительные жидкости отделяются от тканей и поддерживают градиент концентрации, обеспечивая одностороннее движение
Microvilli
- Границы микроворсинок значительно увеличивают площадь поверхности плазматической мембраны (> 100 ×), что способствует большему поглощению
- В мембрану будут встроены иммобилизованные пищеварительные ферменты и канальные белки, чтобы способствовать поглощению материала
Митохондрии
- Эпителиальные клетки ворсинок кишечника будут иметь большое количество митохондрий, обеспечивающих АТФ для активных транспортных механизмов
- АТФ может потребоваться для первичного активного транспорта (против градиента), вторичного активного транспорта (совместного транспорта) или пиноцитоза
Пиноцитотические пузырьки
- Пиноцитоз («питье клеток») — это неспецифическое поглощение жидкости и растворенные растворенные вещества (быстрый способ перемещения в большом количестве)
- Эти материалы будут поглощены через разрыв и преобразование мембраны и, следовательно, содержаться в пузырьке
Электронная микрофотография эпителия ворсинок
.