Актуальные проблемы травмы периферических нервов | Маргасов А.В.
В статье рассмотрены проблемы, посвященные травме периферических нервов. Обобщены результаты множества исследований, внесены предложения по решению проблем хирургического лечения травмы нерва на всех уровнях оказания медицинской помощи.
Введение
За последние десятилетия отмечается неуклонное увеличение частоты техногенных аварий, травматизма на производстве, травм, связанных с дорожно-транспортными происшествиями, бытового травматизма, несчастных случаев. Крайне остро стоит проблема локальных вооруженных конфликтов, терроризма и других боевых действий. Нередко случаются природные катаклизмы и стихийные бедствия. Все эти обстоятельства создают серьезные проблемы для здоровья человека, приводят к росту сочетанных и множественных повреждений.
Среди множества неврологических заболеваний травматическая патология периферической нервной системы занимает особое место, т. к. составляет примерно половину от всех заболеваний периферической нервной системы человека и занимает 1-е место по степени утраты трудоспособности.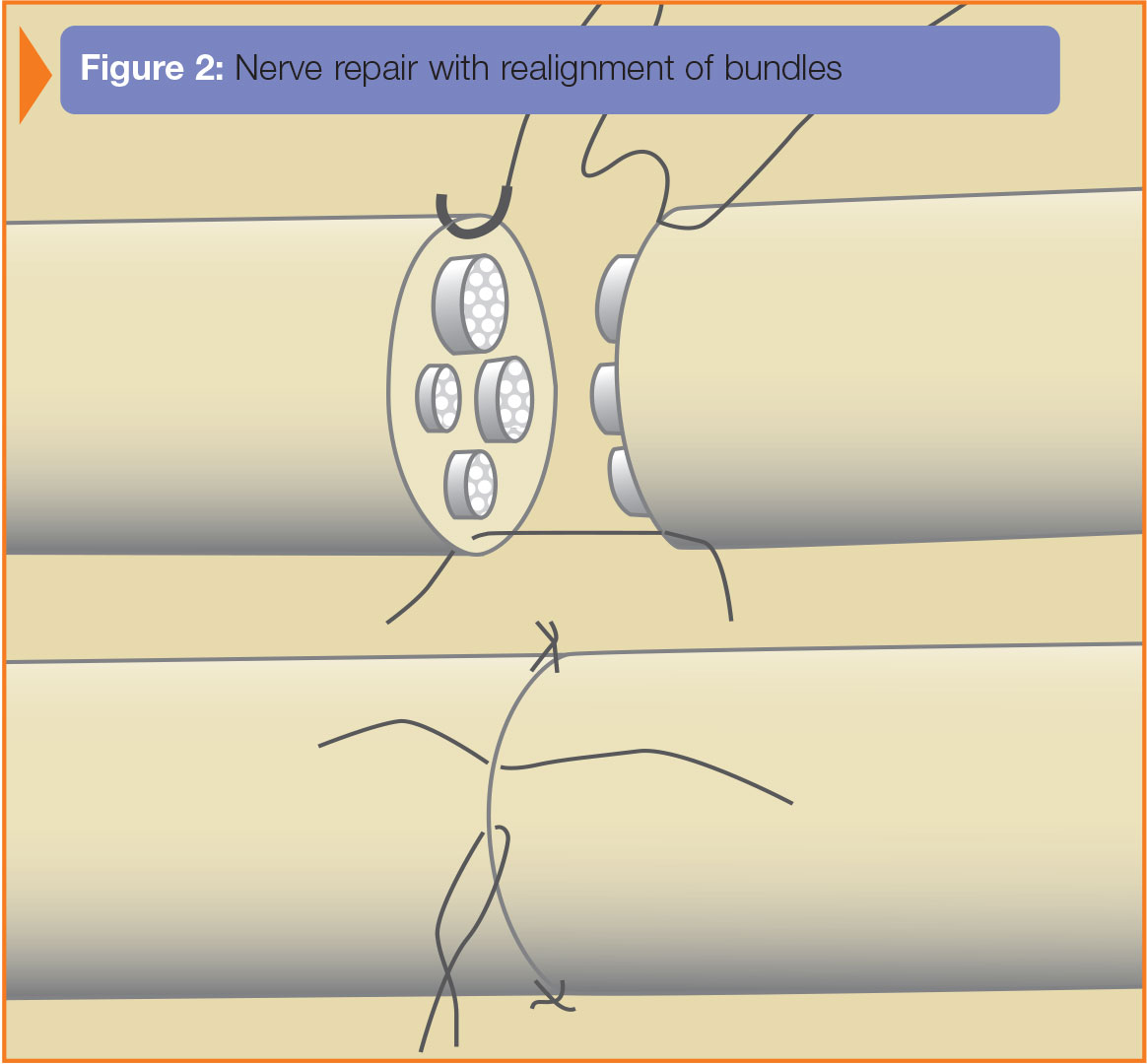 Частота повреждений периферических нервов в мирное время составляет от 1,5 до 13%, а в период боевых действий достигает 20% от числа всех травм. Инвалидизация при всех травмах нерва достигает 60%.
Частота повреждений периферических нервов в мирное время составляет от 1,5 до 13%, а в период боевых действий достигает 20% от числа всех травм. Инвалидизация при всех травмах нерва достигает 60%.
Особенности посттравматических изменений нерва требуют особого организационного, лечебного, функционального подхода к восстановлению поврежденного нерва. Существующие подходы к лечению не позволяют в полном объеме восстанавливать поврежденные нервы, особенно в раннем посттравматическом периоде. В этой связи необходимо указать на основные проблемы оказания помощи при травме нерва и разработать усовершенствованный стандарт лечения травмы нерва. Следует акцентировать внимание на приоритетных задачах, таких как организация лечебного процесса, срочность хирургического лечения, клиническая эффективность, предупреждение осложнений. На настоящий момент существует стандарт лечения травмы нерва, а также разновидности методик лечения, однако их эффективность остается невысокой.
Цель данного теоретического исследования — изучение существующих алгоритмов и уточнение актуальных проблем лечения травмы нерва, создание новых подходов к лечению травмы нерва.
Задачи исследования:
изучение руководящих документов и организационных подходов к лечению травмы нерва;
анализ патоморфологических особенностей при травме нерва;
оценка существующих методов хирургического лечения;
обобщение результатов, внесение предложений по решению проблем лечения травмы нерва.
Методология данного исследования включает эмпирические (наблюдение, опрос), теоретические (аксиоматический метод, в его основе — существующие стандарты оказания медицинской помощи) и общелогические способы познания, к которым относится и структурно-функциональный, позволяющий систематизировать полученные данные и определяющий взаимосвязи между элементами данной системы.
Организация оказания медицинской помощи при повреждениях периферических нервов
Основным документом, регламентирующим порядок оказания помощи пострадавшим с травмой нерва, является Приказ Министерства здравоохранения Российской Федерации от 7 ноября 2012 г. № 616н «Об утверждении стандарта специализированной медицинской помощи при поражениях отдельных нервов, нервных корешков и сплетений» [1]. В данном стандарте указано, что условием и формой оказания помощи является плановое стационарное лечение, вне зависимости от осложнений. Несмотря на то, что при травме нерва чаще используют первичный отсроченный шов, более оправданной тактикой является хирургическое лечение нерва в раннем периоде, после получения повреждения, при соблюдении ряда условий. Однако эффективное хирургическое лечение травмы нерва невозможно без соответствующих специалистов, условий пребывания пострадавшего, технического оснащения стационара и специальных хирургических материалов. В связи с отсутствием в арсенале нейрохирургов специальных материалов для хирургического лечения нерва реконструктивные операции откладываются на поздние сроки от 3 мес. до года, к тому моменту, когда уже сформируются необратимые дегенеративные изменения в тканях.
В стандартах специализированной помощи при травме нерва Минздрава России указаны усредненные значения частоты предоставления помощи, которые на практике весьма вариабельны, в данных стандартах также отсутствует метод тубулизации нерва (табл. 1).
В перечне используемых имплантатов имеется ограниченное количество изделий, которые редко используются или не применяются вовсе при хирургическом лечении поврежденного нерва (табл. 2).
Это связано в первую очередь с тем, что на рынке медицинских изделий отсутствуют специальные и высокоэффективные материалы для восстановления периферических нервов. Помимо этого, на настоящий момент не существует государственной программы финансирования разработок медицинских изделий для оказания экстренной специализированной и высокотехнологичной помощи, а частные инвесторы не готовы вкладывать денежные средства в рискованные и долгосрочные проекты, в результате чего ученые используют полученные гранты и премии только для первичной разработки, тогда как для сертифицированных доклинических и клинических исследований требуются существенные финансовые поступления. Следует отметить, что на настоящий момент обеспечение государственных медицинских учреждений не позволяет в полном объеме выполнять все требования стандартов оказания медицинской помощи.
Большинство повреждений нерва восстанавливается без оперативного вмешательства, с ранним восстановлением функции. Восстановление функции зависит от времени и, в большей степени, от основных патологических процессов в нерве. Несмотря на то, что некоторые повреждения нерва восстанавливаются самостоятельно, в некоторых случаях хирургия — единственный способ лечения для устранения неврологического дефицита или невропатической боли [2]. Рекомендованные средние сроки лечения составляют 10 дней, что является только хирургическим этапом, тогда как необходимо контролировать поздний послеоперационный период, проводить лечебные мероприятия, раннюю реабилитацию. Нетрудоспособность может составлять от 3 мес. до года, в ряде случаев клиническим исходом становится инвалидность.
На настоящий момент существует возможность организовать в травмоцентрах первого и второго уровня оказание специализированной высокотехнологичной помощи при травме нерва в раннем периоде, используя современные разработки в этой области, а также применив неотложную форму оказания помощи: выполнение радикальных операций в первые 12–24 ч после поступления пострадавшего в медицинское учреждение. Это позволит сократить сроки лечения, затраты на длительную реабилитацию, предотвратит утрату нетрудоспособности, что окажет положительный экономический и социальный эффект.
Особенности патоморфологии травм нерва
Дегенеративные изменения, происходящие в нерве после травмы, значительно отличаются от повреждений других органов и тканей и требуют применения специфических методов и материалов для эффективного лечения. Аксональная дегенерация развивается в зависимости от последовательности процессов в зоне травмы, распространяющихся и проксимально и дистально. Разъединенные аксоны и клетки дегенерируют путем апоптоза, названным «хроматолизис» [3]. Эта центральная дегенерация подобна тому, что происходит в других травмированных тканях, включая кожу и мышцу. Однако существенное различие нерва с другими тканями заключается в том, что валлеровская дегенерация дистального аксона начинается от зоны травмы моторного или сенсорного рецептора на отдаленном расстоянии. Валлеровская дегенерация начинается спустя 24–48 ч после повреждения периферического нерва, и дистальные аксоны и окружающий миелин перерождаются. Ближайший аксональный сегмент также дегенерирует назад к перехвату Ранвье, к месту последующего аксонального начала роста [4]. Чем раньше концы поврежденного нерва будут сопоставлены, тем меньше вероятность отставания валлеровской дегенерации от регенерации проросших аксонов. В ряде экспериментов на седалищном нерве животного установлено, что сшивание нерва сразу после пересечения препятствует развитию трофических изменений кожных покровов конечности.
Для обеспечения беспрепятственной регенерации нерва и профилактики послеоперационных осложнений нужны определенные условия, которые позволят восстанавливать поврежденные нервы в раннем периоде. Регенерировавшие волокна первоначально меньше здоровых и отличаются диаметром осевого цилиндра и толщиной миелиновой оболочки. Достижение ими исходных размеров является необходимым условием восстановления нормальной функции. Если существует препятствие для врастания аксонов центрального отрезка нерва в тяжи нейролеммоцитов периферического отрезка (обширная травма, воспалительный процесс, гематома, наличие рубца), то аксоны центрального отрезка растут беспорядочно и могут образовать клубок, называемый невромой. При ее раздражении возникает сильная боль, которая воспринимается как происходящая из первоначально иннервируемой области, например как боль в ампутированной конечности (фантомные боли).
Анализируя имеющиеся данные, можно сделать вывод, что для достижения положительных результатов лечения травмы нерва в раннем периоде необходимо создать условия для формирования и продвижения конуса роста, такие как: защита от окружающих тканей, сохранение собственного аксоплазматического матрикса в пределах зоны роста, локальный противовоспалительный и антибактериальный эффект, отсутствие натяжения нерва. Кроме того, при наложении шва необходимо идентифицировать двигательные волокна и сопоставить пучки как можно точнее друг с другом без перегибов пучков. Концы поврежденных нервов не должны иметь обширных повреждений эпиневрия и нервных структур, а также внутристволовых гематом и признаков нагноения.
Лечение травм периферических нервов
Большинство травм периферических нервов требуют хирургического лечения. Тактика хирургического лечения определяется видом травмы, формой повреждения как патогистологической, так и морфологической. Важную роль в выборе тактики лечения играет точная диагностика уровня повреждений. Для восстановления функции поврежденного нерва применяют разные оперативные приемы: эпиневральный и межпучковый шов, межпучковую аутопластику, внешний и внутренний невролиз, невротизацию поврежденного нерва. Наиболее распространенным способом реконструкции нерва является эпиневральный шов [5].
Лечение резаных и размозженных повреждений периферического нерва представляет основную проблему в восстановительной хирургии и регенеративной медицине. Хотя периферические нервы известны свойственной им способностью восстанавливаться, степень функционального восстановления зависит от ряда факторов, таких как: возраст пациента и общее состояние, тип нерва, срок, прошедший от момента повреждения до хирургического вмешательства, навыки хирурга, а также локализация и протяженность повреждения нерва [6].
Для сближения концов поврежденного нерва применяются следующие способы и приемы: придание конечности определенного положения; мобилизация центрального и периферического отрезков нерва на значительном протяжении; соединение разноименных нервов; этапный шов, перемещение нерва в новое ложе по более короткому пути; резекция кости для укорочения конечности. При повреждениях периферических нервов, которые сопровождаются диастазом более 3 см, на настоящий момент «золотым стандартом» лечения является аутотрансплантация нерва [7]. Для восстановления повреждения периферического нерва используют собственный нерв пациента или синтетический имплантат. Сила тракции в анастомозе влияет на физиологическое натяжение и тем самым на окончательный успех восстановления [8].
Основной задачей операции является более точное соединение пучков нерва и удерживание их в таком положении на срок, необходимый для прочного срастания эпиневрия. Основной причиной отказа от хирургического лечения травмы нерва в раннем периоде являются возможные осложнения: формирование невромы в области шва нерва, несостоятельность шва, вовлечение в рубец соединительной ткани, нагноение в области шва нерва, неточное сопоставление концов нерва.
Наиболее перспективным методом хирургического лечения является тубулизация ушитого нерва при соблюдении определенной методики имплантации и правильном подборе материала имплантата. Трубочки показаны больным, у которых имеется небольшой диастаз нерва или аутогенный нерв недоступен для пересадки (или невозможно полностью восстановить дефект). Вместе с тем трубочки могут быть сделаны из кровеносных сосудов, мышцы и разных синтетических материалов, таких как силикон или полигликолевая кислота [5]. Техника тубулизации с неабсорбируемыми или абсорбируемыми трубками показала многообещающие результаты экспериментально и клинически, когда использовалась для устранения диастаза нерва или при сшивании нерва [2]. Трубка нерва — трубчатая структура, разработанная для устранения диастаза нерва, защиты нерва от окружающей ткани (например, от формирования невромы) и направления аксональной регенерации в дистальную культю нерва [9].
Коллаген является более приемлемым (поскольку он доступен и им легко изолировать и укрывать), он обеспечивает адгезивность для различных типов клеток, что было эффективно продемонстрировано в лаборатории и клинически. Основной его недостаток — продолжительное время резорбции, необходимое для полного биологического распада, в пределах от 3 до 8 мес. Синтетические кондуиты нерва включают алифатические полиэфиры и основанные на сополиэфире, такие как эпсилон-поликапролактон, полигликолевая и полимолочная кислоты [3].
В то время как функциональные результаты от восстановительной трансплантации ограничены иммунологическими последствиями аллотрансплантации, такими как отторжение, регенерация нерва представляет собой дополнительную проблему. Клеточные методы лечения, особенно использование мезенхимальных стволовых клеток, представляются возможным вариантом лечения для усиления регенерации нерва [10]. Применение 3D-моделирования в медицине предпослало возможность использовать данную технологию для создания объемных моделей нейрокондуита в местах разветвления нервных стволов. Микроканалы, напечатанные 3D-способом, направляют рост и образуют аксональные пути через все каналы. Это приводит к отдельным аксональным путям в каналах, где шванновские клетки связываются в аксональные пути в середине канала и соединяются с эпителиальными клетками периферического конца аксона [11].
Перечисленные методики имеют ряд недостатков и не могут применяться при всех видах травм периферических нервов. В связи с вышесказанным становится очевидно, что существует реальная потребность в быстром и простом способе восстановления нерва, который можно применять на этапе квалифицированной медицинской помощи и который обеспечит возможность соединения идентичных нервных пучков, условия для восстановления проводимости и направленного роста нерва, исключит возможные осложнения.
Заключение
В ходе изучения разноуровневых аспектов травматической патологии нерва проанализированы существующие алгоритмы и уточнены актуальные проблемы лечения травмы нерва. Изучены руководяще документы и организационные подходы к лечению травмы нерва, проведен анализ патоморфологических особенностей при травме нерва, оценены существующие методы хирургического лечения, обобщены результаты и проблемы лечения травмы нерва.
В целях решения основных проблем организации оказания помощи пострадавшим с травмой нерва необходимо создать на базе ведущей медицинской организации, оказывающей неотложную нейрохирургическую помощь, центра лечения травмы нерва для оказания высокотехнологичной медицинской помощи в зоне ответственности, куда направлялся бы контингент пострадавших.
Быстрые дегенеративные процессы, развивающиеся после травмы нерва, морфологические особенности строения нерва, механические свойства составляют проблему при выборе сроков и объема оказания помощи.
После изучения имеющихся новых материалов и методов хирургического лечения нерва выявлено, что практически каждый метод в отдельности имеет свои недостатки и узкие показания как по морфологии, так и по патогенезу травмы нерва. Проблема оперативного лечения в раннем периоде может быть решена путем тубулизации шва нерва или установки аутотрансплантата, что позволит предупредить развитие послеоперационных осложнений, обеспечит раннее восстановление нарушенных функций конечности. Биоинертный биополимерный состав трубки снабжен противовоспалительными веществами, которые уменьшат воспалительные реакции в послеоперационном периоде. Отличительной особенностью данной методики является то, что сроки деградации трубки из биополимеров позволят восстанавливать нервы при больших дефектах и обеспечивать быстрый и беспрепятственный рост нерва [12].
Необходимо понимать, что сложные механизмы посттравматической дегенерации нерва требуют постоянного изучения, проведения экспериментов, применения новых материалов и методов. Несмотря на некоторые успехи в отдельных случаях, восстановление нерва не всегда происходит должным образом. Сохраняются определенные риски развития осложнений, на предупреждение которых направлены все современные методы лечения. Большое значение имеют организация оказания помощи, человеческий фактор, наличие необходимого оборудования и материалов.
Таким образом, для решения всех основных проблем лечения травмы нерва необходим комплексный, разноуровневый организационный и методический подход. Наличие узкоспециализированных центров лечения травмы нерва, эффективных методик хирургического лечения позволит оказывать эффективную высокотехнологичную медицинскую помощь в неотложном порядке, что сократит сроки лечения, окажет положительный социальный и экономический эффект.
.
Поражение периферических нервов — симптомы, признаки и лечение
ПОРАЖЕНИЕ ПЕРИФЕРИЧЕСКИХ НЕРВОВ – ВИДЫ
Виды поражений могут быть самыми разнообразными, поэтому в профессиональной неврологии существует специальная классификация, позволяющая разделить их по этиологии и топографо-анатомическому принципу.
По происхождению выделяют следующие виды поражений периферических нервов: инфекционно-аллергические (корь, краснуха и прочие), инфекционные, компрессорно-ишемические, токсические, вертеброгенные (мышечные, суставные, сухожильно-связочные), дисметаболические (недостаток витаминов, ряд заболеваний, например, диабет), идиопатические (наследственные), аллергические (сыворотки, вакцины).
По топографо-анатомическому принципу: полиневриты (ряд воспалений в периферийных нервах), радикулиты (воспалительные процессы в корешках), мононевриты (единичные воспаления в периферийных нервах), фуникулиты (воспалительные процессы в канатиках), плекситы (воспаление сплетений).
Также, поражение периферийных нервов можно разделить на три основных вида, по патоморфологии и патогенезу: невралгии, радикулиты (невриты) и радикулопатии (невропатии).
ЛЕЧЕНИЕ ПОРАЖЕНИЙ ПЕРИФЕРИЧЕСКИХ НЕРВОВ
Процесс лечения достаточно сложный, требует очень качественной предварительной диагностики, позволяющей врачу-неврологу оценить вид и степень повреждения нервов, а также общее физиологическое состояние пациента.
При сложных повреждениях, сочетающихся с политравмой, в первую очередь, проводят восстановление жизненно важных функций организма. Стабилизировав состояние больного, врачи переходят к лечению поврежденных нервов – эта процедура должна проводиться только профильными специалистами с привлечением современного диагностического и манипуляционного оборудования.
В зависимости от сложности проблемы, используют два основных метода лечения: консервативный и хирургический – актуальность применения той или иной методики определяет только лечащий врач, на основании результатов анализов.
Необходимо провести лечение периферических нервов в специализированной клинике или остались какие-либо вопросы? Обратитесь в «ИМПУЛЬС», наши профильные специалисты проведут комплексную консультацию и помогут в решении вашей проблемы!
Периферические нервы — нейрохирургическое лечение и способы блокады периферических нервов
Описание
Поражение периферических нервов может приводить к очень тяжелым последствиям для здоровья человека.
Наиболее часто причиной повреждений становится травма или заболевание. Опыт показывает, что оказать реальную помощь в таких ситуациях может только нейрохирургия.
Типичные причины травматического повреждения периферических нервов:
- удары тупыми предметами;
- травмы от колюще-режущих предметов;
- огнестрельные ранения;
- чрезмерные тракции.
Следствием травматичных воздействий подчас становится частичный или полный перерыв периферического нерва. При этом происходит разрыв оболочек нерва, что чревато развитием дегенеративных процессов. В результате ткани нервов начинают рассасываться, и со временем остаются лишь их оболочки.
Восстановление целостности нервных тканей – чрезвычайно сложная задача, и эффективность ее решения во многом зависит от уровня квалификации хирурга. Тем более, что после разрыва концы нерва начинают расходиться в разные стороны, и в отдельных случаях это расстояние может составлять до 10-15 сантиметров. Способность к проводимости у восстановленного нервного ствола возвращается достаточно медленно – примерно по 1 мм в сутки.
Характерные виды повреждений периферических нервов
- Сдавление периферического нерва – это нарушение, которое проявляется вследствие сдавления нервного ствола рубцовым процессом при переломах, травмах, вывихах, ожогах, ударах и огнестрельных ранениях. Нередко повреждения нервов возникают при хирургических операциях.
- Огнестрельное поражение периферического нерва. В зависимости от скорости снаряда и характера повреждений, здесь возможны два варианта: размозжение участка нервного ствола или его полный перерыв.
- Опухоли периферических нервов – достаточно редкие случаи. Здесь следует помнить важное правило: никаких биопсий (ни пункционных, ни открытых). Или опухоль удаляется полностью, или не нужно трогать вообще.
- Периферический порез лицевого нерва – лечение возможно, как консервативным, так и хирургическим способом. Оперативный метод приносит более быстрые и эффективные результаты, и зачастую является единственно правильным.
- Повреждение плечевого сплетения – один из наиболее сложных и прогностически неблагоприятных случаев. Следствием может стать потеря трудоспособности, косметический дефект, болевой синдром, снижение социальной адаптации.
У нас используются передовые методики нейрохирургического лечения и современные способы блокады периферических нервов.
Оперативное лечение периферических нервов в центре «Новые технологии»
В нейрохирургическом центре им. профессора Г.С. Тиглиева «Новые технологии» осуществляется оперативное и консервативное лечение различных повреждений периферических нервов. В том числе проводится лечение поражений, полученных вследствие колотых, резаных и огнестрельных ран, переломов костей, компрессионных повреждений, лечение опухоли периферических нервов и т.д.
В центре «Новые технологии» выполняются следующие хирургические операции с полным курсом послеоперационной реабилитации:
- шов нерва;
- аутопластика нерва;
- невролиз;
- декомпрессия нервов;
- реинервации;
- операции, связанные с перемещением нерва в новое ложе.
Медицинский центр «Консультант» — Нейрохирургия
Причины нарушения целостности или проводимости периферических нервов
Нарушение целостности, проводимости периферических нервов может наступить вследствие травмы бытового, производственного порядка, ранения или спортивной тренировки. Повреждения нервов конечностей встречается у новорожденных в случаях родовой травмы. Появление гематом, опухолей, аневризмы, рубцевания после травм, образование костных мозолей также приводит к нарушению чувствительности. Нервы сдавливаются, надрываются, истончаются, что приводит к потере целостности, проводимости.
Отсутствие лечения провоцирует замещение нерва соединительной тканью, которая усугубляет процесс, нервные клетки устраняются, конечности полностью теряют чувствительность, снижается функциональность, появляются хронические боли. Оперативное вмешательство «невролиз» направлено на устранение причин сдавливания нерва, восстановление чувствительности ткани, конечности.
Методики диагностирования травмы
Первичный осмотр позволяет обнаружить повреждение периферических нервов у пациентов любого возраста. В процессе оказания первой помощи необходимо выявить возможные нарушения костной системы, мышц и исключить травмы нервных окончаний. Признаки повреждения нервов периферического типа:
- нарушения двигательной функции;
- утрата чувствительности;
- слабость или потеря двигательного рефлекса.
Первичный осмотр подразумевает пальпацию, анализ сложности повреждения.
Невролиз периферических нервов конечностей
Невролиз – выделение нервного волокна с устранением факторов, мешающих его проводимости.
Хирургическое вмешательство показано на разных сроках течения заболевания.
Различается три периода: первичный, отсроченный, поздний – на любом из них может проводиться операция. Невролиз является эффективной методикой нового поколения, которая восстанавливает поврежденные нервные окончания и возвращает конечностям чувствительность, полноценную функциональность.
В некоторых случаях при операции невролиза требуется транспозиция нерва. Процедура смещения или переноса фрагмента нерва из другой части тела в зону нарушения для вживления в место разрыва успешно применяется на практике. Операция проводится для удаления таких факторов, как рубцы, костные мозоли – причины, вызывающие не только нарушение чувствительности, но и мешающие нормальной работоспособности, вызывающие болевой синдром за счет постоянного раздражения нервных окончаний.
Хирургическое вмешательство проводится со значительным разрезом тканей. Это требуется для выделения нерва из окружающих тканей. Хирург определяет необходимость проекционного, внепроекционного окольного или углового доступа на сгибах, уделяя особое внимание сохранению мышечного соединения с нервными волокнами.
Операция подразумевает удаление участков нерва, которые невозможны к восстановлению. После проведения процедуры концы волокон сшиваются, проводится транспозиция фрагмента. Проведение невролиза требует опыта, особой хирургической техники, специальных шовных материалов и микроинструментов.
Послеоперационный период и реабилитация
Восстановление после невролиза включает:
- фиксацию конечности лонгетой для ограничения движения;
- ношение послеоперационных повязок;
- курсы парафинотерапии;
- лечение электростимуляцией;
- прохождение восстановительного курса лечебной физкультуры;
- прием прописанных витаминов, ноотропных препаратов.
Современные тенденции лечения повреждений периферических нервов Текст научной статьи по специальности «Биотехнологии в медицине»
УДК 616.833-001-08
Современные тенденции лечения повреждений периферических нервов
р.Ф. масгутов, а.а. ризванов, а.а. богов (мл.), ар. галлямов, а.п. киясов, а.а. богов
Республиканская клиническая больница МЗ РТ, г. Казань Казанский (Приволжский) федеральный университет
В обзоре освещены современные методы терапии травмы периферических нервов в сочетании с классическими хирургическими способами лечения данной патологии. Представлены потенциальные стимуляторы посттравмати-ческой регенерации периферических нервов, ключевыми из которых являются сосудистый эндотелиальный фактор роста и основной фактор роста фибробластов, играющих роль узловых молекул в патофизиологии нейрорегенерации.
Ключевые слова: травма периферического нерва, ней-ротрофические факторы.
Current trends for treatment of peripheral nerves injuries
R.F. MASGUTOV, A.A. RIZVANOV, A.A. BOGOV (jr.), A.R. GALLYAMOV, A.P. KIYASOV, A.A. BOGOV
Republican Clinical Hospital of the Ministry of Health of the Republic of Tatarstan, Kazan Kazan (Volga region) Federal University
The review describes current treatment methods of peripheral nerve injuries in combination with classical surgical treatments of this pathology. Are presented potential stimulators of posttraumatic regeneration of peripheral nerves, where the key stimulators are vascular endothelial growth factor and basic fibroblast growth factor, which are basic molecules in the pathophysiology of neuroregeneration.
Key words: peripheral nerve injury, neurotrophic factors.
масгутов руслан Фаридович
старший научный сотрудник Травматологического центра 420064, г. Казань, ул. Оренбургский Тракт, д. 138 тел. 8-950-314-02-93, е-mail: [email protected]
Повреждения периферических нервов остаются одной из актуальных проблем современной травматологии и составляют 3-10% от всех травм опорнодвигательного аппарата [1]. Между тем травмы и заболевания периферической нервной системы приводят к стойкой утрате трудоспособности у 60-63% пациентов, более того, почти 80% этой группы составляют лица молодого трудоспособного возраста [2]. Повреждения периферических нервов оказывают существенное влияние на качество жизни в связи с потерей функции и повышения риска развития вторичной инвалидности от падений, переломов и других травм.
В представленном обзоре литературе дан анализ исследований последних лет, которые открывают новую страницу в понимании патофизиологии посттрав-матической регенерации периферического нерва и направляют как специалистов фундаментальных направлений, так и практикующих врачей на изучение и возможности клинического применения ростовых факторов, которые непосредственно принимают участие в восстановлении периферического нерва после его травмы.
Патогенез травмы периферического нерва
При повреждении нерва основным звеном патогенеза является деструкция нейронов.
Нейроны, формирующиеся в процессе развития, соединены в сложные коммуникационные сети для передачи информации от периферических рецепторов сенсорных нейронов в ЦНС (головного и спинного мозга), а также для передачи команд из центральной нервной системы на эффекторные органы, такие как скелетные мышцы, иннервируемые мотонейронами.
Периферический нерв состоит из аксона нейрона, шванновских клеток, фибробластов, а также элементов кровоснабжения нерва. Соединительная ткань известная как эндоневрий окружает периферические аксоны. В периферических нервах аксоны сгруппированы в фасцикулы, окруженные соединительной тканью. Ключевой клеткой для периферических нервов является шванновская клетка. Шванновские клетки образуют осевые цилиндры, их мембрана богата липидами, которые покрывают аксон и называется миелиновой оболочкой. Шванновские клетки с миелиновой оболочкой аксонов поддерживают и направляют аксоны
ьныЕ проблемы медицины том 2 I
Таблица 1.
Влияние нейротрофических факторов на выживание чувствительных нейронов в постнатальном периоде
Нейротрофический фактор Наличие (+) или отсутствие (-) эффекта Авторы
NGF + Horie, Akahori, 1994; Lindsay, 1996
BDNF + Lindsay, 1996; Acheson et al., 1995
NT-3 + Acheson et al., 1995
GDNF + Leclere et al., 1997
во время регенерации нерва после его травмы. Тело нервной клетки является местом синтеза практически всех белков и органелл в клетке. Сложный процесс, известный как антероградный транспорт, непрерывно транспортирует вещества из тела клетки нейрона через аксон к его периферическому концу — в синапс. Эти транспортируемые вещества включают нейротрансмиттеры, которые обеспечивают связь между нейроном и тканями органа по узкой синаптической щели или, как в случае иннервации двигательных нейронов мышц, нервно-мышечном синапсе. С другой стороны, клетки-мишени, например, миоциты, экспрессируют вещества, которые действуют как нейротрофические факторы. Некоторые из этих веществ упаковываются и передаются путем ретроградного транспорта от синапса через аксон к телу нейрона. Таким образом, нейрон и его клетка-мишень постоянно информированы о статусе связи между ними. Было высказано предположение, что информация от клеток-мишеней принимает форму факторов, которые поддерживают существующие соединения нейронов и способствуют регенерации поврежденных нервных клеток. Несмотря на регенерацию, обширные травмы периферического нерва могут привести к параличам всей конечности или дистальных отделов конечностей. Прогноз восстановления при травмах периферических нервов лучше там, где повреждение шванновских клеток минимально [3].
Нейротрофические факторы в механизме регенерации периферического нерва
За полвека исследований выявлено, что нейротрофические факторы способствуют выживанию и процессу роста нейронов. Восстановление нейрона критически зависит от количества нейротрофического фактора. После травмы нервов, периферическая нервная система млекопитающих реагирует путем экспрессии нейротрофических факторов, которые доступны из ау-токринных или паракринных источников. Известны три основные группы нейротрофических факторов: 1) ней-ротрофины; 2) нейротрофические факторы семейства GDNF (англ. Glial cell line-derived neurotrophic factor) и 3) нейротрофические цитокины [4].
Повреждение периферического нерва вызывает ретроградную дегенерацию спинальных ганглиев, что сказывается преимущественно на кожных нейронах малого диаметра. Результаты исследований показывают, что кожные нейроны более чувствительны к повреждениям периферического нерва, чем мышечные, но их регенеративные способности не отличаются друг от друга [5].
Выживаемость нейронов после аксотомии является необходимым условием для регенерации, которой способствуют множество трофических факторов из нескольких источников, в том числе нейротрофины, ней-
ропоэтические цитокины, инсулиноподобный фактор роста (англ. Insulin Growth Factor, IGF), факторы из глиальных клеток — семейства GDNF и др.
Так, после перерезки нерва увеличивается количество нейронов, экспрессирующих м-РНК мозгового нейротрофического фактора (англ. Brain Derived Neurotrophic Factor [6]. Выработка м-РНК BDNF в ответ на травму нерва показана не только в нейронах, но и в клетках-сателлитах [7].
После лигирования спинномозговых нервов уровень м-РНК фактора роста нервов (англ. Nerve Growth Factor, NGF) в спинальных ганглиях возрастает в 4 раза и поддерживается в течение 3 недель [8]. При этом подъем уровня м-РНК BDNF длится не более трех суток и менее выражен по сравнению с NGF.
Трансформирующий фактор роста р (TGFp) служит митогеном для шванновских клеток и нейротрофиче-ским фактором для некоторых дифференцирующихся нейронов. В интактных спинальных ганглиях TGFp и его рецептор экспрессируются преимущественно в малых нейронах и в клетках-сателлитах, окружающих средние и большие нейроны [7]. После повреждения седалищного нерва крысы в эксперименте в клетках-сателлитах увеличивается экспрессия TGFp, что сопровождается возрастанием уровня экспрессии его рецепторов практически во всех субпопуляциях нейронов [7].
In vitro GDNF стимулирует рост аксонов чувствительных нейронов, преимущественно малых, не экспрессирующих рецепторы нейротрофинов и выявляемых при помощи изолектин-В4 [9]. Эти авторы установили, что в той же культуре GDNF поддерживает выживание чувствительных нейронов, но не выявили их принадлежности к конкретным субпопуляциям (табл. 1).
Установлено, что часть нейронов, экспрессирующих trkA и реагирующих, таким образом, на NGF, в конце эмбрионального и в постнатальном периоде перепрограммиру-ется на действие другого нейротрофического фактора, а именно GDNF. В нейронах этой субпопуляции начинают экспрессироваться оба компонента рецептора GDNF — GFRa и тирозинкиназа c-ret [10]. Происходит ли подобное перепрограммирование при регенерации чувствительных нейронов, остается неясным.
Регенераторный рост центральных отростков нейронов различного фенотипа поддерживают конкретные нейро-трофические факторы. Так, рост аксонов, экспрессирующих NF200 и принадлежащих субпопуляции больших про-приоцептивных нейронов, стимулирует только один представитель семейства нейротрофинов — нейротрофин-3 (NT-3) [11] (табл. 2). Из тех же нейротрофинов только NGF поддерживает рост аксонов пептидергических ноцицеп-тивных нейронов. И только один нейротрофический фактор является универсальным стимулятором роста аксонов больших, малых пептидергических и малых непептидер-гических нейронов. Этим фактором является GDNF [11].
актуальные проблемы медицины том 2
Таблица 2.
Экспрессия нейротрофинов и их рецепторов нейронами спинальных ганглиев (по результатам экспериментов на нокаутных мышах, [12]
Нейротрофин/ Рецептор Численность популяции нейронов в % Модальность
NGF 80 Ноцицепторы, терморецепторы, низкопороговые механорецепторы
trkA 80 Ноцицепторы,терморецепторы, низкопороговые механорецепторы
trkB 30 Механорецепторы (Майсснера)
NT-3 60 Проприорецепторы, рецепторы волос и механорецепторы
trkC 20-30 Проприорецепторы
После перерезки седалищного нерва усиливается экспрессия рецептора GDNF, но количество нейронов, реагирующих на этот нейротрофический фактор, не изменяется [13]. При этом объем популяции нейронов, содержащих trkA и отвечающих на действие nGf, после перерезки нерва уменьшается. Эти данные указывают на различный характер регуляции активности генов рецепторов нейротрофических факторов в субпопуляциях нейронов спинальных ганглиев в ответ на аксотомию.
Для судьбы отдельных нейронов важно присутствие нейротрофических факторов, которые способны активировать антиапоптозные внутриклеточные пути [14]. В списке факторов, влияющих на регенерацию нервных волокон, важное место занимают цитокины. Имеются данные о нейропротекторном действии интерлейкина-1b в отношении нейронов коры, которое может реализоваться через влияние этого цитокина на выработку нейротрофических факторов, например, NGF [15]. С другой стороны, in vitro показано, что интерлейкин-1Ь не влияет на выживание нейронов спинальных ганглиев, но ускоряет рост их отростков [16] и вместе с оксидом азота поддерживает в пептидергических нейронах экспрессию циклооксигеназы-2, что усиливает секрецию вещества «Р» [17]. Высказано предположение, что данный цитокин стимулирует секрецию нейротрофиче-ских факторов ненервными (шванновскими) клетками [16], расположенными в потенциальном пространстве роста аксонов.
Представление об активации интерлейкином-1Ь регенерации нервных проводников основано на данных о стимулирующем влиянии продуцируемого макрофагами интерлейкина-1 на образование NGF шваннов-скими клетками in vitro и in vivo. Шванновские клетки in vitro и в регенерирующем нерве сами синтезируют интерлейкины и экспрессируют их рецепторы, что свидетельствует о возможной аутокринной регуляции. Нейроны также способны синтезировать цитокины. У интактных крыс до 80% нейронов спинального ганглия экспрессируют интерлейкин-1Ь [18]. Это преимущественно большие и средние нейроны. Данный цито-кин экспрессируется в незначительной части малых пептидергических нейронов. Присутствие рецепторов интерлейкина-1 выявлено в большинстве нейронов спинального ганглия, а также в окружающих нейроны клетках-сателлитах [18], что свидетельствует об участии интерлейкина-1Ь в ауто/паракринной регуляции функции чувствительных нейронов.
Травмированные нейроны должны перейти от режима передачи к режиму роста и начать продуцировать белки, такие как, например, GAP-43 (англ. Growth
Associated Protein), тубулин и актин, а также другие нейропептиды и цитокины, которые имеют потенциал для стимуляции аксональной регенерации. Аксоны должны достигать дистального отрезка нерва при условии, когда поддержка их роста осуществляется в достаточной степени. Шванновские клетки в дистальном отрезке нерва проходят через фенотипические изменения, которые должны быть благоприятны для аксональной регенерации. Шванновские клетки играют незаменимую роль в содействии регенерации за счет увеличения ими синтеза поверхностных молекул клеточной адгезии, таких как N-CAM (англ. Neural Cell Adhesion Molecule), Ng-CAM/L1 (англ. Neuron Glia Cell Adhesion Molecule), N-кадгерин и L2/HNK-1, путем изменения базальной мембраны, которая содержит многие белки внеклеточного матрикса, таких как ламинин, фибронектин и тенасцин. Аксональной регенерации могут способствовать факторы, которые повышают потенциал роста аксонов нейронов и оптимизируют регенерацию дистального конца нерва в сочетании с его оперативным лечением [19].
Основные методы лечения повреждения нервов являются: нейрорафия, нейрорафия с последующей дистракцией нерва, аутонервная пластика, эндопротезирование биодеградирующими материалами (например: neuragen® nerve guide).
Перспективными потенциальными стимуляторами регенерации периферических нервов являются сосудистый эндотелиальный фактор роста (англ. Vascular Endothelial Growth Factor, VEGF), основной фактор роста фибробластов (англ. basic Fibroblast Growth Factor, bFGF или FGF2) и NGF.
Эффект NGF иллюстрирует исследование по трансплантации в поврежденный седалищный нерв крысы микрокапсул, содержащих NGF в чистом виде. В группе с NGF количество восстановленных аксонов было большим, расположение нервных волокон было более упорядоченным, проводимость нерва восстанавливалась в разы быстрее, чем в контрольной группе. Также в сравнении с контрольной группой животных было отмечено, что в послеоперационном периоде раны заживали значительно быстрее [20].
Основной фактор роста фибробластов считается мощным средством для улучшения восстановления после повреждения нейронов, однако экзогенно применяемый bFGF неэффективен из-за кратковременного эффекта. Для усиления эффекта в естественных условиях разработана новая система доставки потенциальных стимуляторов путем внедрения bFGF в желатин гидрогель, который медленно биодеградирует. В этом исследовании эффекты bFGF-гидрогеля при
ьные проблемы медицины том 2 I
Рисунок 1.
Плазмида, содержащая сосудистый эндотелиальный фактор роста (VEGF) и основной фактор роста фибробластов (FGF2)
Масгутов РФ. и др. Клеточная трансплантология и тканевая инженерия, 2011
травматическом повреждении лицевого нерва были исследованы на морских свинках. Лицевой нерв был обнажен и зажат в вертикальной части с использованием микрощипцов. Животные были подвергнуты одной из следующих трех процедур: группа А — контроль, без введения bFGF; группа В, одноразовое введение bFGF в нерв; группа С, применение bFGF-гидрогеля. Шесть недель спустя функции лицевого нерва были оценены тремя тестами: наблюдения за мимикой, электро-физиологические тестирования и гистологические исследования. Результаты показали, что одноразовое применение bFGF не влияло посттравматическое восстановление нерва. В группе с применением гидрогеля достигнуты лучшие результаты во всех тестах [21].
Местное применение VEGF индуцирует, по крайней мере, два события: пролиферацию шванновских клеток и образование новых сосудов, которые играют важную роль в процессе регенерации нервов. Однако результаты показывают, что эффект от предварительной обработки факторами роста является локальным и ограниченным и не влияет на синтез нейропептидов [22].
Преодоление диастаза периферического с помощью биодеградирующией трубки из политетрафторэтилена (ПТФэ) является относительно новым, но хорошо зарекомендовавшим хирургическим подходом для восстановления дефектов нервных стволов до 4 см между его концами на разных уровнях верхней конечности [23].
Восстановление периферического нерва с помощью трубки из ПТФЭ и введением факторов роста фибро-бластов также подтверждается в другом исследовании. В данном исследовании bFGF был помещен в эндопротез для седалищного нерва крысы. Кроме того, в послеоперационном периоде, животные системно получали 1 мг/кг/сут FK506 (такролимус) — мощный иммунодепрессант с нейротрофическими свойствами. Регенерация нерва оценивалась с помощью стандартных двигательных (индекс седалищного нерва) тестов и тестов на чувствительность, начиная со 2-й недели после операции. Животные, которые получали bFGF и FK506, показали значительно более быстрое восстановление после травмы, чем у контрольной группы [24].
После повреждения периферического нерва Шван-новские клетки начинают экспрессировать bFGF, что нехарактерно для интактного нерва. Данный фактор рассматривается в качестве узловой молекулы в патофизиологии травмы периферических нервов [25].
В исследовании по восстановлению лицевого нерва после его перерезки и дальнейшего сшивания на крысах было показано, что у животных с введением bFGF восстановление нерва произошло раньше, а количество миелинизированных волокон было большим, чем у животных без введения bFGF [26].
Кроме того, на модели преодоления диастаза седалищного нерва крысы при помощи аутонервной вставки показано, что введение мультицистронной плазмиды pBUD-VEGF-FGF2 (Рис 1), экспрессирующей клонированные гены VEGF и bFGF человека, в центральный и периферический отрезки нерва, а также непосредственно в аутонервную вставку стимулирует реваскуляризацию и регенерацию периферического нерва. В эксперименте с аутонервной вставкой введение pBUD-VEGF-FGF2 в область повреждения нерва приводит к двукратному увеличению количества S100-иммунопозитивных (шванновских) клеток в прилежащих к диастазу участке периферического отрезка нерва [27, 28].
Заключение
Таким образом, сочетание классических хирургических способов восстановления нервных стволов и методов прямой терапии ростовыми факторами ускоряет аксональный рост, стимулирует реваскуляризацию нерва, что подтверждается многочисленными экспериментальными исследованиями и, в итоге, улучшает результаты посттравматического восстановления функции реиннервации поврежденного органа или ткани.
Благодарность
Выполнение данного научного исследования финансируется за счет темы государственного автономного учреждения здравоохранения «Республиканская клиническая больница Министерства здравоохранения Республики Татарстан» «Усовершенствование и разработка новых методов лечения у больных с повреждением плечевого сплетения и периферических нервов».
ЛИТЕРАТУРА
1. Берснев В.П. Хирургия позвоночника, спинного мозга и периферических нервов / Е.А. Давыдов, Е.Н. Кондаков // Специальная литература.
— СПб, 1998. — С. 368.
2. Кубицкий А.А. Хирургическое лечение повреждений периферических нервов верхней конечности методами тракционного удлинения и аутонервной пластики: автореф. дис. … канд. мед. наук / А.А. Кубицкий.
— Казань, 2002. — 24 с.
3. Massing M.W., Robinson G.A., Marx C.E., Alzate O., Madison R.D. Frontiers in Neuroscience. Alzate O., editor // Source Neuroproteomics. — Boca Raton (FL): CRC Press; 2010. — Chapter 15.
4. Boyd J.G., Gordon T. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury // Mol Neu-
robiol. — 2003 Jun. — Vol. 27, № 3. — P. 277-324.
5. Welin D., Novikova L.N., Wiberg M., Kellerth J.O., Novikov L.N. Survival and regeneration of cutaneous and muscular afferent neurons after peripheral nerve injury in adult rats // Exp Brain Res. — 2008 Mar. — Vol. 186, № 2. — P. 315-23.
6. Zhou X.F. et al. Injured primary sensory neurons switch phenotype for brain-derived neurotrophic factor in the rat // Neuroscience. — 1999. — Vol. 92, № 3. — P. 841-853.
7. Zhou X.F. et al. Satellite-cell-derived nerve growth factor and neurotrophin-3 are involved in noradrenergic sprouting in the dorsal root ganglia following peripheral nerve injury in the rat // Eur. J. Neurosci. — 1999. — Vol. 11, № 5. — P. 1711-1722.
8. Shen H., Chung J., Chung K. Expression of neurotrophin mRNAs in
актуальные проблемы медицины том 2
the dorsal root ganglion after spinal nerve injury // Brain Res. Mol. — 1999.
— Vol. 64. — P. 186-192.
9. Leclere P. et al. Effects of glial cell line-derived neurotrophic factor on axonal growth and apoptosis in adult mammalian sensory neurons in vitro. Neuroscience. — 1997. — Vol. 5. — P. 545-558.
10. Molliver D.C. et al. IB4 binding DRG neurons switch from NGF to GDNF dependence in early postnatal life // Neuron. — 1997. — Vol. 19, № 4. — P. 4S49-4S61.
11. Ramer M., Priestley J., McMahon S. Functional regeneration of sensory axons into adult spinal cord // Nature. — 2000. — Vol. 403. — P. Э12-Э16.
12. Farinas I., Cano-Jaimeza M., Bellmunta E., Soriano M. Regulation of neurogenesis by neurotrophins in developing spinal sensory ganglia // Brain Research Bulletin. — 2002. — Vol. 57, № 6. — P. S09-S16.
13. Kashiba H., Hyon B., Senba E. Glial cell line-derived neurotrophic factor and nerve growth factor receptor mRNAs are expressed in distinct subgroups of dorsal root ganglion neurons and are differentially regulated by peripheral axotomy in the rat // Neuroscience Letters. — 1998. — Vol. 4.
— P. 107-110.
14. Mattson M.P., Lindvall O. Neurotrophic factor and cytokine signaling in the aging brain // Greenwich. JAI Press. — 1997. — P. 299-345.
15. Strijbos P., Rothwell N. Interleukin-1 beta attenuates excitatory amino acid-induced neurodegeneration in vitro: Involvement of nerve growth factor // J. Neurosci. — 1995. — Vol. 15, № 5. — P. 3468-3474.
16. Horie H., Sakai I., Akahori Y., Kadoya T. IL-1 beta enhances neurite regeneration from transected-nerve terminals of adult rat DRG // Neuroreport. — 1997. — Vol. 8, № 8. — P. 1955-1959.
17. Morioka N. et al. Nitric oxide synergistically potentiates interleukin-1 beta-induced increase of cyclooxygenase-2 mRNA levels, resulting in the facilitation of substance P release from primary afferent neurons: involvement of cGMP-independent mechanisms // Neuropharmacology. — 2002. — Vol. 43, № 5-10. — P. 868-876.
18. Copray J.C. et al. Expression of interleukin-1 beta in rat dorsal root ganglia // Neuroimmunology. — 2001. — Vol. 118, № 8. — P. 203-211.
19. Fu S.Y., Gordon T. The cellular and molecular basis of peripheral
nerve regeneration // Mol Neurobiol. — 1997. — Vol. 1-2. — P. 67-116.
20. Song M., Chen S.Z., Han H., Xiong Y. An experimental study on repair of peripheral nerve injury by transplantation of microcapsulated NGF-expressing NIH 3T3 cells // Zhonghua Zheng Xing Wai Ke Za Zhi. — 2005.
— Vol. 1. — Р. 53-7.
21. Komobuchi H., Hato N., Teraoka M., Wakisaka H., Takahashi H., Gyo K., Tabata Y., Yamamoto M. Basic fibroblast growth factor combined with biodegradable hydrogel promotes healing of facial nerve after compression injury: an experimental study // Acta Otolaryngol. — 2010. — Vol. 130, № 1. — Р. 173-8.
22. Sondell M., Lundborg G., Kanje M. Vascular endothelial growth factor stimulates Schwann cell invasion and neovascularization of acellular nerve grafts // Brain Res. — 1999 Nov 6. — Vol. 846, № 2. — Р. 219-28.
23. Stanec S., Stanec Z. Reconstruction of upper-extremity peripheral-nerve injuries with ePTFE conduits // J Reconstr Microsurg. — 1998 May.
— Vol. 14, № 4):227-32.
24. Lanzetta M., Gal A., Wright B., Owen E. Effect of FK506 and basic fibroblast growth factor on nerve regeneration using a polytetrafluoroethyl-ene chamber for nerve repair // Int Surg. 2003. — Vol. 88, № 1. — Р. 47-51.
25. Borin A., Cruz O.L., Ho P.L., Testa J.R., Fukuda Y. The action of topical basic fibroblast growth factor in facial nerve regeneration // Otol Neurotol. — 2010 Apr. — Vol. 31, № 3. — Р 498-505.
26. Toledo R.N., Borin A., Cruz O.L., Ho P.L., Testa J.R., Fukuda Y. The action of topical basic fibroblast growth factor in facial nerve regeneration // Otol Neurotol. 2010. — Vol. 31, № 3. — Р 498-505.
27. Масгутов Р.Ф., Салафутдинов И.И., Богов А.А. (мл.), Трофимова А.А., Ханнанова И.Г., Муллин Р.И., Исламов Р.Р., Челышев Ю.А., Богов А.А., Ризванов А.А. Стимуляция посттравматической регенерации седалищного нерва крысы с помощью плазмиды, экспрессирующей сосудистый эндотелиальный фактор роста фибробластов. Клеточная трансплантология и тканевая регенерация. — 2011. — Т. VI, № 3.
28. Челышев Ю.А., Мухамедшина Я.О., Шаймарданова Г.Ф., Николаев С.И. Прямая доставка терапевтических генов для стимулирования посттравматической нейрорегенерации // Неврологический вестник. — 2012. — T. XLIV, вып. 1. — С. 76-83.
Казанские врачи лечат повреждения нервов уникальным методом
В Казани медики разработали уникальный метод лечения пациентов, у которых повреждены периферические нервы и сосуды. Операция состоит всего из одной инъекции и длится не больше часа. При этом восстановительный период сокращается в разы.
Жир спасет от инвалидности. В Казани начали делать уникальные операции. Пациентам с ишемической болезнью нижних конечностей, с повреждением периферических нервов и сосудов больше не приходится проходить длительный курс лечения. Подвижность суставов и чувствительность возвращаются уже через пару месяцев. Секрет – в клетках жировой ткани.
Рядовой осмотр перед нерядовой операцией: кисть левой руки Александра Евсеева ничего не чувствует больше восьми лет, после травмы был поврежден нерв. Годы реабилитации и даже хирургическое вмешательство не смогли вернуть былого крепкого рукопожатия.
«Я открыл кухонную дверь, стекло треснуло, руку порезал. Обратился к врачам, мне нерв сшили. И вот такие последствия – ненормальная чувствительность», — рассказывает он.
Теперь Александр Евсеев — рука об руку с инновациями. В республиканской клинической больнице Татарстана испытывают новый метод лечения.
У пациента забирают фрагмент жировой ткани, в лаборатории извлекают клеточный концентрат и в тот же день вводят в поврежденный участок. Операция длится не больше часа. Концентрат работает как своеобразная фабрика, выделяя биологически активные вещества для скорого восстановления организма.
«Мы получаем совершенно новый феномен в лечении периферического нерва. Это невиданное ранее восстановление за несколько месяцев, когда нерв должен был бы дорасти до этого уровня за полтора-два года», — говорит руководитель травматологического отделения номер 2 Республиканской клинической больницы министерства здравоохранения Республики Татарстан Андрей Богов.
Этот метод пока не применяют в России, да и зарубежных публикаций на эту тему нет. Для лечения подобных травм и заболеваний в мире используются стволовые клетки. В Татарстане решили мыслить шире: в концентрате, что выделяют из жировой ткани, не только стволовые клетки. В этом и новизна, и секрет успеха. Он не вызывает отторжения, аллергических реакций, и не ведет к онкологии. В республике эффект исследовали более чем на полусотне пациентов. Результаты удивили даже авторов.
«В разрабатываемой нами технологии применяются клетки без дополнительного культивирования. Это концентрат клеток, который, конечно, содержит и стволовые клетки. Но так как мы не проводим дополнительных манипуляций с ними, они наиболее биологически безопасны», — говорит заведующий кафедрой генетики Института фундаментальной медицины и биологии Казанского федерального университета Альберт Ризванов.
Клеточный концентрат жировой ткани буквально ставит на ноги пациентов с такими диагнозами, как ишемия нижних конечностей, повреждение периферических нервов и сосудов. То, на что раньше уходили месяцы, а то и годы лечения, теперь врачуют одной инъекцией.
Александр Евсеев меж клеток не вглядывается, для него важен ощутимый результат – чтобы пальцы, наконец, начали чувствовать прикосновения. Врачи уверены — и здесь клетки сложатся как нужно. К примеру, как у Аделя Мухаметдинова. После страшного ДТП он мог остаться инвалидом — но дело в руки взяли специалисты. Через полгода после операции он смог пожать приятелю руку. Спустя полтора – машет врачам и ученым с благодарностью.
Травма периферической нервной системы
Родовая травма периферической нервной системы — это повреждение нервных корешков, нервных сплетений, периферических и черепных нервов под воздействием внешних механических факторов в процессе родов. Сила такого воздействия намного превосходит предел выносливости указанных анатомических структур, в результате чего возникает сдавление, разрушение или разрыв тканей. Все эти нарушения сопровождаются местным или общим расстройством кровообращения, отеками, переломами костей и т.д. Родовой акт – сам по себе очень сложный процесс, состоящий из компрессии, вращения, сжатия и тракции, и если он усугубляется неправильным предлежанием или большими размерами плода, применением акушерских шипцов, то вероятность возникновения родовой травмы очень высока.
Патоморфологически выделяют несколько вариантов повреждения периферических нервов:
- Полный (преганглионарный) отрыв нервного корешка от спинного мозга
- Постганглионарный разрыв корешка
- Разрыв аксона с сохранением миелиновой оболочки.
- «Ушиб» нерва с сохранением аксона и миелиновой оболочки.
Последний вариант повреждения наименее травматичен и создает все предпосылки для полного восстановления периферического нерва.
Любой вид родовой травмы нервной системы нуждается в обязательном осмотре такого пациента врачом-неврологом. Квалифицированную консультацию опытного врача Вы можете получить в «Медицинском центре Берсенева». Наша клиника уже несколько десятков лет специализируется на лечении заболеваний позвоночника и нервной системы, включая и родовые травмы.
Наиболее часто наблюдается травма плечевого сплетения, лицевого, диафрагмального, лучевого и срединного нервов. Чаще всего это возникает при использовании акушерских шипцов.
Травма плечевого сплетения в большинстве случаев обусловлена акушерскими пособиями, применяемыми при затруднении выведения головки и плечиков плода. В результате выполняемых тракций и ротаций головки или плечиков происходит натяжение корешков верхнегрудных и нижнешейных сегментов спинного мозга над поперечными отростками шейных позвонков.
Плечевая натальная плексоневропатия впервые была описана учеными Дюшеном (1872) и Клюмпке (1885).
Акушерский парез (травма плечевого сплетения) чаще всего отмечается у детей, рожденных в ножном или ягодичном предлежании, с большой массой тела, при узком тазе, при чрезмерных тракциях за головку плода, при сдавлении шеи, при затруднении выведения плечиков, при запрокидывании ручек плода.
В зависимости от уровня повреждения плечевая плексоневропатия подразделяется на три основных разновидности:
- верхний (проксимальный) тип
- нижний (дистальный) тип
- тотальный тип
Паралич Эрба- Дюшена или верхний (проксимальный) тип
Это наиболее частый вид родового паралича, который встречается 1 раз на каждые 1500-2000 рождений и связан с повреждением первичного пучка плечевого сплетения или шейных корешков С5-С6.
Клиническая картина характеризуется тем, что пораженная верхняя конечность вяло свисает вдоль туловища, разогнута во всех суставах и повернута внутрь, кисть в ладонном сгибании повернута назад и снаружи (характерный симптом «кукольной ручки» Новика). Плечо опущено. Наблюдается атрофия мышц плеча и предплечья, невозможность поднять плечо выше горизонтального уровня, согнуть в локтевом суставе, т.е. характерно нарушение функции проксимального отдела руки, движения же в пальцах сохранены.
Головка чаще наклонена и повернута к паретическому плечу, засчет этого шея кажется укороченной и с большим количеством поперечных складок. Отмечается снижение мышечного тонуса в проксимальных отделах конечности, рефлекс с двуглавой и трехглавой мышц не вызывается.
В некоторых случаях в процесс вовлекаются корешки С3-С4, и тогда в клинической картине преобладают симптомы, характерные для повреждения диафрагмального нерва (учащенное дыхание, одышка, икота, цианоз, ассимметрия движений грудной клетки).
Паралич Дежерин-Клюмпке или нижний (дистальный) тип
Данный тип плексопатии возникает при травме спинного мозга на уровне С7-Т1 или среднего и нижнего пучков плечевого сплетения. Он встречается намного реже, чем проксимальный (верхний) типа поражения и характеризуется нарушением функции дистального отдела верхней конечности. Характерно резкое снижение или полное выпадение функции мышц-сгибателей кисти и пальцев. Кисть пассивно свисает, движения отсутствуют, мышечный тонус снижен. В некоторых случаях кисть находится в положении «когтистой лапы», что связано с повреждением волокон лучевого или локтевого нервов.
В связи с вовлечением в процесс срединного нерва, иногда встречается симптом «ишемической перчатки», связанный с трофическими расстройствами (побледнение или покраснение кожи кисти, отек).
Тотальный паралич верхней конечности (паралич Керера)
Тотальный тип повреждения отмечается при повреждении на уровне С5-Т1 сегментов спинного мозга и характеризуется сиптомами, присущими для проксимального и дистального типов акушерского пареза. Это наиболее тяжелый вариант протекания акушерского пареза. Отмечается полная обездвиженность руки, мышечная гипотония, трофические расстройства, выпадение рефлексов, снижение всех видов чувствительности.
Повреждение лицевого нерва
Повреждение лицевого нерва чаще всего встречается при лицевом предлежании, длительном стоянии головки в родовых путях, узком тазе, при сдавлении околоушной железы акушерскими шипцами, при самопроизвольных родах.
Существует неутешительная статистика, доказывающая, что парез лицевого нерва в 80% случаев связан именно с наложением щипцов на область головки.
В клинической картине преобладает односторонний парез мимической мускулатуры: угол рта опущен, что особенно видно во время плача ребенка, слабо выражена носогубная складка на стороне поражения, ребенок не может плотно сомкнуть веки, снижен или отсутствует поисковый рефлекс Куссмауля, затруднено сосание, отсутствуют морщины и складки на лбу.
Паралич поясничного сплетения встречается достаточно редко, при неумелом извлечении плода за ножки.
В «Медицинском центре Берсенева» применяется уникальная методика лечебного воздействия, способствующая восстановлению поврежденных нервных волокон. Разработанный и внедренный в практику доктором Берсеневым метамерный метод лечения уже несколько десятков лет позволяет бороться с заболеваниями центральной и периферической нервной системы. Метамерные технологии способствуют регенерации периферической нервной системы благодаря сочетанию рефлексотерапии и нейротрофического действия вводимых препаратов.
Записавшись на консультацию в наш медицинский центр, Вы получите квалифицированные ответы врачей-неврологов и узнаете, как лечить травмы периферических нервов, корешков и сплетений. В нашей клинике для каждого пациента разрабатывается индивидуальная программа лечения и реабилитации, позволяющая достичь оптимального результата.
Для записи на лечение в «Медицинский центр Берсенева» Вы можете позвонить по телефону контакт-центра (044) 238-22-31 или заполнить форму онлайн-записи у нас на сайте.
Механизмы травмы и восстановления
Введение
Повреждения периферических нервов — распространенные состояния с широким диапазоном групп симптомов в зависимости от тяжести и пораженных нервов. Хотя существует много знаний о механизмах травм и регенерации, надежных методов лечения, обеспечивающих полное функциональное восстановление, мало. Этот обзор призван обобщить различные способы классификации этих повреждений в свете десятилетий исследований повреждения и регенерации периферических нервов.
Анатомия периферического нерва
Периферическая нервная система состоит из трех типов клеток: нейрональных клеток, глиальных клеток и стромальных клеток. Периферические нервы передают сигналы между спинным мозгом и остальным телом. Нервы состоят из различных комбинаций моторных, сенсорных и вегетативных нейронов. Эфферентные нейроны (моторные и вегетативные) получают сигналы через свои дендриты от нейронов центральной нервной системы, в первую очередь используя нейромедиатор ацетилхолин.Афферентные (сенсорные) нейроны получают сигналы через свои дендриты от специализированных типов клеток, таких как тельца Пакциниана для тонких ощущений и другие. Эти сигналы отправляются в ЦНС, чтобы предоставить сенсорную информацию в мозг и, возможно, интернейроны в спинном мозге, когда необходим рефлекторный ответ 1 .
Ключевую роль в поддержании и функционировании периферических нервов играют не нейроны, а другие клетки. Шванновские клетки покрывают нервы слоем миелина и обеспечивают трофическую поддержку за счет высвобождения важных нейротрофов, таких как фактор роста нервов (NGF).Миелин улучшает скорость проводимости за счет ограничения участков переноса ионов вдоль аксона к узлам Ранвье, что приводит к более быстрому, «скачущему» распространению потенциала действия, что называется скачкообразной проводимостью. Наиболее сильно миелинизированные волокна — это крупные двигательные нейроны (тип Aα), за которыми следуют веретена афферентных мышц (тип Aβ). Скорость нервной проводимости в этих нейронах составляет примерно 30-120 м / с. Немиелинизированные нейроны (тип C), такие как сенсорные нейроны, участвующие в передаче боли и температуры, и постганглионарные симпатические нервы, являются самыми медленными, проводя их со скоростью примерно 1-2 м / с () 2,3 .
Таблица 1
Типы и свойства нервных волокон
| Класс волокна | Миелин | Диаметр (мм) | Скорость проводимости (м / с) | Спинной мозг | Местоположение | Функция |
|---|---|---|---|---|---|---|
| Aα | + | 6-22 | 30-120 | Ипсилатеральный спинной столб | Воздействующий на мышцы | Двигатель |
| Aβ | + | 6-22 | 30-120 | Контралатеральный спиноталамический тракт | Афферентный от кожи и суставов | Тактильно, проприоцепция |
| Aγ | + | 3-8 | 15-35 | Ипсилатеральный дорсальный столб | Эфферент на мышечные веретена | Мышечный тонус |
| Aδ | + | 1-4 | 5-30 | Контралатеральный спиноталамический тракт | Афферентные сенсорные нервы | Боль, холод, температура, прикосновение |
| B | + | 1-3 | 3-15 | Преганглионарная | Преганглионарная симпатическая | Различные вегетативные функции |
| СК | — | 0.3-1,3 | 0,7-1,3 | — | Постганглионарная симпатическая | Различные вегетативные функции |
| dC | — | 0,4-1,2 | 0,1-2,0 | Контралатеральный спиноталамический тракт | Афферентные сенсорные нервы | Различные вегетативные функции Боль, тепло, температура, прикосновение |
Ненейронные клетки и соединительные ткани, окружающие нейроны аксоны обеспечивают сложный каркас из соединительной ткани стромы 4 для нерва и важны для понимания и классификации повреждений нервов.Внутри отдельных аксонов находится самый глубокий структурный слой, эндоневрий. Окружая эндоневрий, периневрий связывает аксоны вместе, образуя пучки. Наружный соединительнотканный слой нерва, эпиневрий, состоит из двух частей. Между пучками рассредоточен эпифасцикулярный эпиневрий, в то время как собственно нервный ствол окружает эпиневральный эпинерий. Микрососуды постепенно разветвляются по нерву в соответствии со структурными слоями, обеспечивающими кровью аксоны.Из-за своего более периферического расположения эпиневральные сосуды более восприимчивы к травмам, чем более глубокие сосуды нерва 5 .
Классификация повреждений нервов
Повреждения периферических нервов создают для пациентов различные проблемы, от легкого дискомфорта до пожизненных нарушений. Схема классификации предоставляет врачам и ученым общий язык для эффективного обсуждения патофизиологии нервов (). Седдон первым разделил нервные повреждения на три категории в зависимости от наличия демиелинизации и степени повреждения аксонов и соединительной ткани нерва 6 .Самая легкая форма повреждения называется нейрапраксией и определяется очаговой демиелинизацией без повреждения аксонов или соединительных тканей. Нейрапраксия обычно возникает из-за легкого сжатия или растяжения нерва и приводит к снижению скорости проводимости. В зависимости от тяжести демиелинизации эффекты могут варьироваться от асинхронной проводимости до блокады проводимости, вызывающей мышечную слабость. Следующий уровень называется аксонотмезисом, который включает прямое повреждение аксонов в дополнение к очаговой демиелинизации при сохранении целостности соединительной ткани нерва.Самая тяжелая форма травмы называется нейротмезисом, когда происходит полное рассечение аксонов и слоев соединительной ткани, при котором наблюдается полный разрыв нерва.
Таблица 2
Классификация нервных травм по Седдону и Сандерленду
| Седдон | Сандерленд | Травма |
|---|---|---|
| Нейрапраксия | Степень I | Фокальная сегментарная демиелинизация |
| Аксонотмезис | Степень II | Аксон поврежден интактным эндоневрием |
| Аксонотмезис | Степень III | Аксон и эндоневрий повреждены интактным периневрием |
| Аксонотмезис | Степень IV | Аксон, эндоневрий и периневрий, поврежденные интактным эпиневрием |
| Нейротмезис | Grade V | Полная перерезка нерва. |
| Степень VI (MacKinnon & Dellon) | Смешанные уровни повреждения вдоль нерва |
Позднее Сандерленд расширил эту классификацию, чтобы определить степень повреждения соединительной ткани 7 . В его схеме классификации степень I и степень V соответствовали нейропраксии и невротмезису Седдона соответственно. Однако все степени II-IV — это формы аксонотмезиса с возрастающим повреждением соединительной ткани.При степени II наблюдается повреждение аксона без повреждения соединительной ткани. Степень III включает повреждение эндоневрия, а степень IV включает повреждение периневрия (). Позже Маккеннон и Деллон представили поражение степени VI для обозначения сочетания повреждений степени III-V вдоль поврежденного нерва, хотя его использование не было широко распространено 8 . Попытки упростить эту схему путем классификации нервов как недегенеративных или дегенеративных были предложены Thomas и Holdroff в 1993 году, но клиническое значение этого упрощения все еще остается под вопросом 9 .
Классификация нервных травм
От: Цао Б., Булис Н., Бетукс Ф., Мюррей Б. Травма нервной системы, травма периферического нерва. В: Дарофф: Неврология Брэдли в клинической практике, 6-е изд. 2012 с. 984-1001. (Изображение любезно предоставлено Cleveland Clinic, 2006. Иллюстратор, Дэвид Шумик, BS, CMI.)
Компрессионная травма
Причины
Компрессионные травмы не всегда охватываются обычно используемыми классификационными схемами. Тем не менее, нет никаких сомнений в том, что большинство компрессий периферических нервов подпадают под общий класс нейрапраксии или повреждений нервов I степени и обычно возникают в местах, где нервы проходят через узкие анатомические отверстия.Наиболее частыми участками верхней конечности являются запястный канал и локтевой канал. Как и ожидалось, травма нерва I степени определяется очаговой демиелинизацией в месте сдавления при отсутствии повреждения аксонов и соединительной ткани. Компрессии могут носить острый или хронический характер. Острые компрессии, наблюдаемые при лучевой мононевропатии, обычно возникают после ночного внешнего сжатия, например, когда рука висит на стуле, и обычно сопровождаются преходящей парестезией, онемением и опущением запястья.Полное восстановление сильно сдавленного нерва может занять от нескольких недель до нескольких лет. Напротив, хронические компрессии, наблюдаемые при синдроме запястного канала, представляют собой прогрессирующее ухудшение состояния, которое сохраняется без надлежащего вмешательства. Симптомы могут начинаться с парестезии и онемения дистальных отделов, но, в отличие от острых компрессий, часто со временем прогрессируют до мышечной слабости и мышечного истощения, в зависимости от степени повреждения аксонов на более поздних стадиях 10 .
Патофизиология
Световая и электронная микроскопия показывают, что нормальная морфология нервов и нервно-мышечные соединения все еще присутствуют при хронических компрессионных повреждениях 11,12 .Однако наблюдается деградированная, более тонкая миелиновая оболочка, о чем свидетельствует увеличенное отношение g (отношение диаметра аксона к диаметру аксона плюс диаметр миелиновой оболочки) и уменьшенная длина междоузлий (расстояние между соседними узлами Ранвье). В присутствии этой демиелинизации также наблюдаются пролиферация шванновских клеток, дедиффенция и увеличение резцов Шмидта-Лантермана (SLI). SLI — это цитоплазматические компоненты шванновских клеток, которые, как считается, поддерживают метаболизм миелиновой оболочки, поэтому увеличение SLI предполагает, что шванновские клетки увеличивают свой метаболизм, чтобы подвергнуться повторной миелинизации в присутствии демиелинизации, что обычно наблюдается при хроническом сжатии. травмы 13 900 10.
Существуют различные предложенные механизмы, которые, как считается, приводят к компрессионным травмам 14,103 . С анатомической точки зрения сужение отверстий приводит к повышению давления в этом месте, сдавливанию кровеносных сосудов и приводит к ишемии нервов, как это происходит при васкулите и артеросклоротических заболеваниях. Другой предложенный механизм является результатом более низкого давления, что снижает венозный возврат и может привести к венозному застою. В этом состоянии со временем может образоваться экстраневральный отек с последующей фиброзной и рубцовой тканью вокруг нерва и, в конечном итоге, внутриневральным отеком.Изучение этих двух механизмов порождает вопрос, почему в основном поражаются шванновские клетки, а не сами нейроны.
Хроническая компрессионная травма когда-то считалась более легкой формой валлеровской дегенерации. Это давно опровергается из-за отсутствия повреждения аксонов при этой травме. Роль макрофагов и их митогенных факторов, таких как интерлейкины 1, 6, 10 и 12, в стимулировании активности шванновских клеток была высказана, но маловероятна, поскольку шванновские клетки способны пролиферировать в отсутствие макрофагов 15 .Кроме того, поскольку незаметное постепенное повреждение при хроническом сжатии не приводит к воспалительной реакции, макрофаги прибывают медленно и после того, как большая часть пролиферации шванновских клеток уже произошла, еще больше отдаляя макрофаги от вероятных виновников 16 .
Более поздние исследования показали, что напряжение сдвига само по себе может вызывать демиелинизацию, пролиферацию и ремиелинизацию шванновских клеток 17 . Эти эксперименты in vitro проводились в камерах с жестким контролем, где концентрации газа, давление и растворенные вещества в среде регулировались и контролировались в режиме реального времени.В таких контролируемых условиях приложение сдвиговых напряжений приводит к измененной экспрессии белка шванновских клеток с ключевыми изменениями, приписываемыми измененной экспрессии интегрина B4 17 .
Большая часть работы in vivo по изучению компрессионной невропатии была связана с использованием биологически инертных полимерных силиконовых силиконовых трубок, более широко используемых для хирургических имплантатов и протезов. Эти пробирки использовались для создания надежных моделей компрессионной невропатии на многих моделях животных, таких как крысы 29 , кролики 30 и мыши 32 , а также были использованы для изучения двойного раздавливания in vivo
31 .Модели компрессионной невропатии обычно оцениваются исследованиями нервной проводимости, которые в этих моделях показывают постепенное снижение скорости нервной проводимости при отсутствии нейронального или мышечного повреждения, что было бы очевидно, если бы был обнаружен измененный потенциал действия сложной мышцы (CMAP), но не было 14,31,103 .
Двойное раздавливание
В области компрессии существует еще одна форма травмы, известная как двойное раздавливание 23 . Двойное сдавливание относится к повышенной склонности нерва к развитию компрессионной невропатии, когда обнаруживается проксимальное компрессионное поражение того же нерва.Этот феномен был впервые определен Аптоном и МакКомасом в 1973 году после наблюдения, что 81 из 115 пациентов с запястным или локтевым туннелем также имели поражение нервной системы на шее 24 . Обратная форма двойного сдавливания, удачно названная обратным двойным сдавливанием, была позже описана Далином и Лундборгом в 1990 году после наблюдения за пациентами с защемлением локтевого нерва на запястье, позже у которых развилось аналогичное повреждение проксимального отдела локтевого сустава 25 . Морфология поражения, наблюдаемого при двойном сдавливании, идентична поражениям, наблюдаемым при хроническом сдавливании, с уникальным аддитивным эффектом множественных участков с подпороговыми уровнями давления, способствующими надпороговому воздействию на нерв.Прерывание аксонального движения и потока может служить объяснением того, как две незначительные области сжатия могут сговариваться, чтобы сформировать эффект, который имитировал бы более сильное сжатие инсульта в одном месте. В статье Уилборна 1997 года обсуждаются эти модели, их ограничения и указывается на необходимость дальнейших исследований и клинического злоупотребления термином «двойное поражение» в случаях, которые не соответствуют конкретным параметрам, описанным Аптоном, МакКомасом и Далином и Лундборгом. 26 . Существование двойного сдавливания было подтверждено в различных исследованиях при подостром сжатии 27,28 и сильном сжатии 29 с использованием нервов животных in vitro и in vivo .
Травма с раздавливанием и рассечением
Причины
Травмы с раздавливанием могут вызывать нервные повреждения различной степени, которые могут представлять любой класс схем, описанных Седдоном 6 или Сандерлендом 7 . Более того, большинство этих травм, вероятно, часто представляют собой смешанные травмы, подобные тем, которые предложили Деллон и Маккиннон 21 .
Раздавливание обычно происходит в результате острого травматического сдавливания нерва тупым предметом, например летучей мышью, хирургическим зажимом или другим раздавливающим предметом, которое не приводит к полному перерезанию нерва.Напротив, при перерезании, также известном как нейротмезис или повреждение нерва V степени, нерв полностью прекращается, обычно из-за разрыва ножом, огнестрельного оружия, осколка стекла и т. Д. 34 . Баллистические травмы — это особый случай, который имеет тенденцию сочетать как рассечение, так и раздавливание нерва ударной волной, которая проходит через ткань после прохождения пули. Это оказывает как разрывающее, так и сдавливающее действие на нерв даже без фактического прохождения снаряда через сам нерв.
Механизм восстановления
Когда орган-мишень становится денервированным, реиннервация может происходить двумя способами: через коллатеральное разветвление интактных аксонов или путем регенерации поврежденного аксона 35 . При травмах, когда повреждено 20-30% аксонов, коллатеральное ветвление является основным механизмом восстановления. Это начинается в первые 4 дня после травмы и будет продолжаться примерно 3-6 месяцев, пока не наступит выздоровление. Как и следовало ожидать, наблюдается увеличение размера двигательных единиц и гипертрофия оставшейся иннервируемой мышцы в попытке компенсировать первоначальную денервацию других участков мышцы.Однако со временем мышца в конечном итоге атрофируется, поскольку волокна без иннервации сжимаются и опережают способность оставшихся мышечных волокон к расширению 34 . Прорастающих аксональных ветвей больше, чем фактическое количество нервов, которые в конечном итоге иннервируют орган-мишень 36 . Те ветви, которые не получают нейротрофические факторы от органа-мишени, подвергаются процессу отсечения и обречены на вырождение 37,38 .
При травмах, затрагивающих более 90% популяции аксонов в нерве, регенерация аксонов является основным средством восстановления. 39 .Для достижения полного восстановления нерв должен пройти через три основных процесса: дегенерацию Валлера (процесс очищения дистальной культи), регенерацию аксонов и реиннервацию органа-мишени. Нарушение любого из этих процессов может способствовать плохому функциональному результату, обычно наблюдаемому у пациентов с повреждениями периферических нервов.
Валлеровская дегенерация
Аксоны, которые подвергаются травматическому повреждению, будут подвергаться валлеровской дегенерации, чтобы создать микросреду, способствующую повторному росту и реиннервации аксонов.Этот процесс происходит в течение первой недели после травмы, когда появляются типичные маркеры аксонального повреждения: потеря целостности клеточной мембраны и разрушение аксонального цитоскелета. Первоначально набухание происходит на обоих концах поврежденного нейрона из-за бесполезного продолжения ретроградного и антероградного аксонального транспорта 40 .
Изменения на дистальной культе
Аксональные изменения на дистальном конце разорванного нерва в конечном итоге приводят к разрушению культи нерва, чтобы освободить место для вновь регенерирующего аксона.Отличительной чертой этой фазы является гранулярный распад цитоскелета 41 . Это происходит после того, как внезапный приток внеклеточных ионов, в первую очередь Ca + и Na + , приводит к каскаду событий, напоминающих апоптоз, который служит для рекрутирования макрофагов с использованием сигналов, вырабатываемых шванновскими клетками. Было показано, что дистальная культя нерва способна передавать потенциал действия через несколько часов после перерезки, факт, используемый для изучения синаптической передачи и мышечной функции в изолированных нервно-мышечных препаратах 42-44 .Совсем недавно было обнаружено, что мРНК нейротрофического фактора головного мозга (BDNF) и нейротрофического фактора линии глиальных клеток (GDNF) активируется, в то время как нейротрофин-3 и цилиарный нейротрофический фактор (CNTF) подавляются в дистальных культях. рассеченных большеберцовых нервов. Хронически денервированные культи дистального нерва сохранят эти изменения через 6 месяцев после травмы, если регенерация не произошла 45 .
Изменения проксимальной культи
Изменения проксимальной культи различаются в зависимости от местоположения травмы относительно тела нейрона и тяжести травмы.Разрушение проксимальной культи ограничено и обычно прогрессирует только до первого узла Ранвье. Однако, если место повреждения находится очень близко к телу нейрона, может произойти апоптоз 46 . При тяжелых травмах проксимальная часть нерва подвергается хроматолизу, изменяя генетический мотив клетки, чтобы изменить ее фокус на фенотип регенерации. Во время этого процесса белки, связанные с ростом, такие как GAP-43, тубулин и актин, активируются, а нейрофиламенты (участвующие в поддержании диаметра аксонов) подавляются 47-49 .Эндогенные нейропротекторы, такие как белок теплового шока-27 (HSP27), активируются, возможно, чтобы способствовать выживанию поврежденного нейрона 50,51 . Когда аксон в конечном итоге достигает своего органа-мишени и требуется созревание нейронов, экспрессия гена нейрона возвращается из регенеративного состояния в состояние поддержания.
Шванновские клетки и макрофаги
Шванновские клетки являются первичными медиаторами в запуске многих событий валлеровской дегенерации, и изменения в экспрессии их белков в месте повреждения являются ключом к регенерации аксонов.В отсутствие контакта с аксонами шванновские клетки переходят в немиелинизирующее поведение — подавляя экспрессию нескольких белков, таких как PMP22, Krox-20 52 , P0 и коннексин-32 53 . Синтез приостанавливается, а дифференцировка стимулируется за счет продукции C-jun 54 , и продуцируются нейротрофические факторы, такие как NGF и CNTF 55 . Вместе эти факторы способствуют формированию пула новых шванновских клеток под защитой эндогенных соединений, таких как эритропоэтин 56 .Эти изменения позже прогрессируют митогенами, высвобождаемыми из проксимальных нейронов культи, такими как АТФ и нейрегулин 57,58 , которые вместе с ацетилхолином помогают созреть новые шванновские клетки до миелинизирующего фенотипа 59 .
Шванновские клетки очищают дебрис посредством фагоцитоза и рекрутирования макрофагов 60 . Этот эффект зависит от белка MAC-2, который поддерживает фагоцитоз шванновских клеток 61 . Клетки Шванна также являются источником моноцитарного хемоаттрактантного белка-1 (MCP-1), который задействует макрофаги 62 .
Макрофаги, привлеченные шванновскими клетками, быстро достигают места повреждения нерва. 63 — процесс, зависящий от разрушения гемато-нервного барьера. Макрофаги также спасают ценный холестерин в поврежденных нервах и вырабатывают аполипопротеин E, среди других липопротеинов 64-66 . Наконец, сами макрофаги продуцируют факторы, которые дополнительно способствуют пролиферации шванновских клеток 64,67 .
После очистки от остатков миелина дедифференцированные шванновские клетки пролиферируют на оставшихся эндоневральных трубках внеклеточного матрикса (ЕСМ).В совокупности они известны как полосы Бангнера, и образовавшаяся полая трубка обеспечивает путь для регенерирующего аксона, чтобы вырасти заново 68 . Больший успех реиннервации из регенеративного аксона наблюдается, когда эндоневральная трубка не повреждена и формирование невромы благоприятно без трубки, которая со временем сжимается, если аксон не заселяется в течение четырех месяцев 34,39 . Это накопление каналов Na + в эктопических аксонах, неспособных найти трубки, что может привести к нейропатической боли.
Регенерация нервов
Хотя изменение генетической экспрессии нейрона из состояния покоя в состояние регенерации происходит одновременно с событиями валлеровской дегенерации, сама регенерация аксонов начинается после завершения валлеровской дегенерации 69 . На дистальном конце проксимального зачатка образуется конус роста. Было обнаружено, что кальций играет роль в проксимальной культи, способствуя формированию конуса роста 70 . Через несколько часов после травмы конус роста отправляет филаподии для анализа микросреды.На своем пути к дистальному культю нерва филоподии изначально ориентированы случайным образом, но приобретают направление, как только экспрессия актина и миозина активируется в теле клетки 49,71,72 .
Подвижность конуса роста зависит от наличия определенных рецепторов в его мембране. Его можно привлечь или оттолкнуть с помощью контактных или химических средств, известных как нейротрофизм 73,74 . Примерами направляющих молекул являются семафорины, эфрины, нетрины и щели.Тормозные направляющие молекулы, такие как коллапсин-1, способствуют коллапсу конуса роста. Нейротрофины, такие как BDNF, уменьшают восприимчивость конусов роста, способствуя регенерации нервов 75 . Скорость регенерации может сильно зависеть от расположения вдоль нейрона, в котором проксимальные сегменты могут увеличиваться на 2-3 мм / день, в то время как более дистальные сегменты могут прогрессировать со скоростью 1-2 мм / день.
Путь конуса роста может быть нарушен рубцовой тканью, и конусы роста высвобождают протеазы и активаторы плазминогена, чтобы освободить его путь.Это также необходимо для устранения любых взаимодействий между клетками или клетками с матрицей ненейронных клеток, которые препятствуют его пути 70 .
Шванновские клетки играют существенную роль в стимулировании регенерации аксонов, поскольку они являются основным источником нейротрофических факторов, которые обычно взаимодействуют с рецепторами тирозинкиназы, изменяя профиль экспрессии генов нейрона и способствуя регенерации. Это сообщение передается через ретроградный транспорт 76 . NGF имеет низкий уровень экспрессии в здоровых нервах, но активируется в шванновских клетках во время повреждения.NGF способствует росту и пролиферации шванновских клеток, а также обнаруживается на рецепторах выстилающих полос шванновских клеток Бангнера, обеспечивая трофику прорастающего аксона 39 . Было обнаружено множество нейротрофических факторов, функции которых варьируются от улучшения выживаемости клеток за счет механизмов предотвращения апоптоза до стимулирования регенерирующих факторов в нейронах и в шванновских клетках ( 77 ). Кроме того, шванновские клетки продуцируют факторы, способствующие развитию нейритов, которые включены в ЕСМ, такие как фибронектин и ламинин.Конусы роста используют эти белки для адгезии к базальной пластинке эндоневральных трубок 78 .
Таблица 3
Сводка изменений молекулярной экспрессии в аксотомированных нейронах после повреждения периферического нерва.
| Поражение | Местоположение | Ссылки | ||
|---|---|---|---|---|
| Нейротрофические факторы / рецепторы | ||||
| BDNF | ↑ | Трансекция | DRG | Кашиба и Сенба (1999) |
| ↑ | CCI | DRG | Obata et al.(2003) | |
| ↑ | Раздавливание | DRG | Ernfors et al. (1993) | |
| ↑ | Перевязка | DRG | Fukuoka et al. (2001) | |
| ↑ | Трансекция | MN | Gu et al. (1997) | |
| ↑ | Раздавить | DRG | Tonra et al. (1998) | |
| ↑ | Перевязка позвоночника | DRG | Shen et al.(1999) | |
| Trk | = | Crush | DRG | Ernfors et al. (1993) |
| Ret (рецептор GDNF) | ↑ | Трансекция | MN | Hammarberg et al. (2000) |
| = | Разрез | DRG | Bennett et al. (2000) | |
| GFRalpha2 | ↓ | Трансекция | DRG | Bennett et al.(2000), Hoke et al. (2000) |
| GFRalpha1, GFRalpha3 | ↑ | Трансекция | DRG | Bennett et al. (2000) |
| Ret и GDNFalpha1 | ↑ | Разрушение | DRG / MN | Naveilhan et al. (1997) |
| NT-3 | = | Перевязка позвоночника | DRG | Shen et al. (1999) |
| = | Разрез | SC | Funakoshi et al.(1993) | |
| trkB | ↑ | Трансекция | MN | Hammarberg et al. (2000) |
| ↑ | Раздавливание | SC / DRG | Ernfors et al. (1993) | |
| NGF | = | Пересечение | MN | Gu et al. (1997) |
| ↑ | Перевязка позвоночника | DRG | Shen et al. (1999) | |
| NGF-R | ↑ | Раздавливание | MN | Ernfors et al.(1989) |
| 16 кДа протеин, ассоциированный с панкреатитом (Reg-2) | ↑ | Трансекция | DRG | Averill et al. (2002) |
| Факторы транскрипции | ||||
| c-fos | ↑ | Трансекция | MN | Gu et al. (1997) |
| ↑ | Трансекция | DH | Kajander et al.(1996) | |
| ↑ | CCI | DH | Kajander et al. (1996) и Ro et al. (2004) | |
| с-июн | ↑ | Пересечение | DRG | Jenkins and Hunt (1991), Broude et al. (1997) и Кенни и Кочиш (1997) |
| ↑ | Crush | MN | Jenkins and Hunt (1991) | |
| Сигнал трансд. и актив. транск.3 (STAT3) | ↑ (A) | Разрез | DRG | Qiu et al. (2005) |
| Активирующий фактор транскрипции 3 (ATF-3) | ↑ | Трансекция | MN / DRG / DH | Tsujino et al. (2000) |
| NFκB | ↑ | Трансекция | DH | Pollock et al. (2005) |
| ↑ (A) | PNSL / CCI | DRG | Ma and Bisby (1998) | |
| Isl 1 | ↑ | Раздавливание | DRG | Vogelaar et al.(2004) |
| DRG11, LmX1b, Pax3 | = | Разрушение | DRG | Vogelaar et al. (2004) |
| Прочие | ||||
| TNF-альфа рецептор 1 (p55) | ↑ | Crush | DRG | Ohtori et al. (2004) |
| Нектин-3 | ↑ | Трансекция | MN | Zelano et al. (2006) |
| P38 митоген-активированная протеинкиназа (MAPK) | ↑ (A) | CCI | DRG | Obata et al.(2004a) |
| Моноцитарный хемоаттрактантный белок-1 (MCP-1) | ↑ | CCI | DRG / MN / DH | Zhang and De Koninck (2006) |
| Фактор роста фибробластов 2 (FGF-2) | ↑ | Перевязка | DRG | Madiai et al. (2003) |
| Транспортер глутамата EAAC1 | ↓ | CCI | DH | Wang et al. (2006) |
| cAMP-ответный элемент-связывающий белок (CREB) | ↑ (P) | CCI | DH | Miletic et al.(2004) |
Проникающие макрофаги после фагоцитоза миелина также способствуют регенерации нервов за счет секреции IL-1, который индуцирует экспрессию NGF в шванновских клетках и увеличивает плотность рецепторов NGF на шванновских клетках. Этот механизм прямой связи приводит к секреции митогенов, чтобы еще больше запустить пролиферацию шванновских клеток 79 . Макрофаги, однако, также секретируют антагонист рецептора IL-1, который уменьшает повторный рост миелинизированных и немиелинизированных аксонов при применении через трубку для имплантации у мышей с перерезанными седалищными нервами 80 .
Когда конус роста достигает эндоневральной трубки, у него больше шансов достичь органа-мишени. Созревание должно произойти до того, как функциональная связь будет завершена. Процесс созревания включает ремиелинизацию, увеличение аксонов и, наконец, функциональную реиннервацию. Отросток аксонов продуцирует АТФ и ацетилхолин, которые способствуют изменению фенотипа шванновских клеток с немиелинизирующего на миелинизирующий 59 .
Недостатки для восстановления
Хотя повреждения периферических нервов не опасны для жизни, они могут вызвать значительное снижение качества жизни пациента 81 .Это мотивирует дальнейшие исследования для поиска способов оптимизации восстановления. Реиннервация не является синонимом полного функционального восстановления. Это формирующееся понимание этих факторов наряду с ключевой ролью клинически значимых факторов, которое означает полное выздоровление от исключения к правилу. Ключевые элементы нейрорегенерации — это расстояние между промежутками, валлеровская дегенерация, специфичность управления аксонами и жизнеспособность органов-мишеней.
Наличие интактной эндоневральной трубки чаще приводит к лучшему результату регенерации нерва.Таким образом, поражения степени II, которые вызывают повреждение только аксонов без какого-либо повреждения окружающей соединительной ткани, имеют оптимальные условия для возобновления роста аксонов. Однако поражения III и IV степени не только имеют поврежденную эндоневральную трубку, затрудняющую формирование соответствующих полос Бангнера, но также имеют повышенное образование рубцовой ткани, которое может быть значительным сдерживающим фактором для конуса роста, что приводит к дезорганизованному разрастанию.
Чем более дистальнее повреждение нейрона, тем более вероятно, что оно вылечится с очень проксимальными повреждениями, близко к телам нейрональных клеток, что часто вызывает запрограммированную гибель нейронных клеток.Длина промежутка отрицательно коррелирует с успешной регенерацией, связывая размер повреждения с точностью отростка аксона.
Во время восстановления нерва отросток аксонов ремиелинизируется резидентными шванновскими клетками. Однако эта миелинизация обычно намного тоньше, чем обычно, с предсказуемыми электрическими последствиями 82 . В недавнем исследовании с использованием модели раздавливания седалищного нерва на мышах было обнаружено, что аксональный нейрегулин-1 (NRG1) типа III и I восстанавливает нормальные уровни миелинизации при сверхэкспрессии у трансгенных мышей.Неожиданно денервированные шванновские клетки также экспрессировали NRG-1 типа I как паракринный / аутокринный сигнал, предполагая, что полная ремиелинизация регенерированных аксонов может не происходить из-за просто недостаточной стимуляции факторами роста нейронов из шванновских клеток 83 .
Даже если регенерированный аксон способен достичь цели, созревание возможно только при сохранении органа-мишени. Это включает стабилизацию нервно-мышечного соединения (НМС) в случае мотонейронов 84 .НМС состоит из терминального конца моторного аксона, терминальных шванновских клеток (TSC) и концевой пластинки мышечных волокон, содержащих рецепторы ацетилхолина (AChR) 85 (). Присутствие аксона обеспечивает трофическую поддержку концевой пластинки, а длинный дегенерированный аксон приводит к дисперсии кластеров AChR. Предполагается, что агрин, гликопротеин, высвобождаемый дистальным нервным окончанием, является ключевым регулятором образования кластеров 86 . У мутантных мышей с дефицитом агрина наблюдается плохое формирование кластеров и синаптогенез после повреждения нерва 87 .Взятые вместе, эти данные предполагают, что добавление агрина в NMJ может сохранить ACh-R-ключи и может привести к лучшему сохранению органа-мишени. Помимо дисперсии, оборот AChR укорачивается в десять раз, когда аксон отсутствует на концевой пластине 88 .
Нервно-мышечное соединение
1. Изображение нервно-мышечного соединения и ПЭМ. От: Lee Y, Thompson WJ. Глава 54 — Нервно-мышечное соединение позвоночных. Мускул, 2012; 2: 775-787.
Мышечные волокна атрофируются уже через 3 недели после денервации с образованием отложений коллагена в эндомизии и перимизии.Однако структурная архитектура мышцы и целостность концевой пластинки могут сохраняться до 1 года 69,89 . Через 2 года произошел необратимый мышечный фиброз вместе с мышечной дегенерацией, что привело к необратимой потере функциональной мышечной ткани. Сенсорные конечные органы, такие как тельца Пакциниана, тельца Мейснера и клетки Меркеля, могут длиться до 2-3 лет, поэтому сенсорная функция все еще может быть восстановлена даже после того, как функция мышц окончательно утрачена 33 . Время для поддерживающих рост фенотипов шванновских клеток, связанных с тканями-мишенями, также ограничено 90 .Перенос нервов использовался, чтобы попытаться предотвратить атрофию денервации, но полная сила и функция мышц редко полностью восстанавливаются 91 .
Экспериментальные стратегии
Различные модели повреждения нервов используются для разработки эффективных методов лечения повреждений нервов. Модели выживания нейронов in vitro включают культуры клеток, трехмерные культуры тканевой инженерии, органотипические культуры и культуры глиальных клеток, тогда как модели in vivo чаще всего включают повреждение седалищного нерва различных видов.
Модели in vitro
Нейроны ганглия задних корешков являются наиболее часто используемыми нейронами в экспериментах на культуре нейрональных клеток. Однако животные производят лишь небольшое количество клеток, что может быть недопустимо 92 . Обычной культивируемой клеточной линией является клеточная линия феохромоцитомы крысы (PC12), которая выполняет нейроэндокринную функцию и может быть индуцирована к дифференцировке в симпатические нейроны взрослых с использованием NGF 93 . Клеточные культуры обычно используются для исследований in vitro прямого воздействия лекарств, субстратов и факторов роста на нейроны.Нейроны, культивированные совместно с глиальными клетками и клетками других типов, были полезны для выяснения взаимодействий между этими клетками.
Культивирование нервных клеток также предоставило информацию о зависимости нервных клеток от соответствующей микросреды для оптимального восстановления. Созданные с помощью тканевой инженерии трехмерные культуры используют ECM на основе искусственной ткани для изучения роста нейритов. Сигналы контактного наведения могут быть добавлены для изучения прохождения аксонов и наведения через эндоневральную трубку 94,95 .Органотипические культуры используют естественную среду нейронов таким образом, что цитоархитектура тканей и взаимодействия между типами клеток сохраняются 96,97 .
In vivo — модели повреждения нервов
Животные модели повреждений раздавливания и разрыва могут быть созданы для отслеживания клинически значимых поражений и обеспечения более глубокого изучения молекулярных и клеточных процессов в определенных типах нервов (т.е. смешанных, моторных, сенсорных). Грызуны являются наиболее часто используемыми животными из-за их недорогих жилищных затрат и аналогичного распределения нервных стволов среди людей.Кроме того, имеется большое количество данных о генетической, клеточной и системной физиологии на моделях грызунов, включая трансгенных животных 98 .
Самая распространенная модель травмы — седалищный нерв. Хотя повреждения седалищного нерва у людей возникают редко из-за глубокого анатомического расположения в нижней конечности, модель на животных предоставляет множество информации относительно восстановления определенных типов нервов и их потенциала к восстановлению при попытке достичь эндоневральных канальцев.Повреждения бедренного нерва у крыс имеют отношение к изучению конкретных типов нервов, потому что у них одна исключительно двигательная ветвь, а другая — исключительно сенсорная 99,100 .
Повреждения срединного нерва важны, поскольку они обычны в клинической практике, но на практике анализы функции срединного нерва у грызунов чрезвычайно трудны. Некоторые работы посвящены электрофизиологическим методам наряду с оценкой функционального результата у крыс 101 и, в последнее время, у мышей, использующих швы 12/0 для восстановления после травмы 102 .
Измерение результатов на моделях in vivo
Надлежащие измерения результатов при оценке функционального восстановления нервов имеют решающее значение для получения окончательных результатов. Измерения результатов включают в себя: подсчет количества конкретных структур, таких как аксоны, нейроны, реиннервируемые концевые пластинки и реиннервируемые двигательные единицы, оценку вызванных потенциалов нервов и мышц с помощью электрофизиологических исследований и оценку сократительной силы и веса мышц. Также существуют поведенческие исследования, в частности, для оценки функции седалищного нерва с помощью анализа пешеходных дорожек и тестирования функции сенсорной гиперчувствительности.Число аксонов и сократительные силы мышц могут быть наиболее достоверной оценкой на ранних этапах регенерации аксонов. На этой фазе ростки регенерируют асинхронно и достигают своей конечной цели в разное время, поэтому могут предоставить вводящую в заблуждение информацию с использованием вызванных потенциалов или других форм оценки 103 .
От лабораторных исследований до прикроватного применения
Охснер Дж. 2013 Весна; 13 (1): 100–108.
, MD, PhD, FRCS * и, PhD †
Wale Sulaiman
* Отделение нейрохирургии, Ochsner Clinic Foundation, и Медицинская школа Университета Квинсленда, Клиническая школа Охснера, Новый Орлеан, Луизиана
Тесса Гордон
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
* Отделение нейрохирургии, Фонд клиники Окснера и Школа медицины Университета Квинсленда, Клиническая школа Окснера, Новый Орлеан, Луизиана
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
Адресная корреспонденция Wale Sulaiman, MD, PhD, FRCS, Отделение нейрохирургии, Ochsner Clinic Foundation, 1514 Jefferson Hwy., New Orleans, LA 70121. Тел: (504) 842-2181 Факс: (504) 842-6531 Электронная почта: gro.renshco@namialusw Авторские права © Академическое подразделение Фонда клиники Окснера Эта статья цитируется в других статьях в PMC.
РЕФЕРАТ
Объективы
Мы рассматриваем современную нейробиологию повреждения и регенерации нервов, особенно в том, что касается восстановления полезной функции у пациентов, которые получили травмы крупных нервных стволов, таких как плечевое сплетение.
Методы
В этом обзоре основное внимание уделяется исследованиям, проведенным в нашей лаборатории в Окснеру и в других лабораториях, связанных с нейробиологией повреждения нервов, с акцентом на то, как некоторые из ключевых результатов исследований на животных помогают нам понять патофизиологию плохого функционального восстановления после нервных окончаний. травма, повреждение.
Выводы
Опубликованные исследования нейробиологии повреждения и регенерации нервов убедительно свидетельствуют о том, что хроническая денервация шванновских клеток, хроническая аксотомия нейронов и неправильное направление регенерирующих аксонов в неправильные эндоневральные трубки в первую очередь ответственны за плохое функциональное восстановление. Эффект атрофии мышечной денервации вторичен. Экспериментальные терапевтические стратегии (которые мы в настоящее время исследуем в нашей лаборатории в Охснере) для борьбы с этими 3 нейробиологическими явлениями могут улучшить восстановление функции у пациентов, перенесших нервные повреждения.
Ключевые слова: Аксотомия , цитокины , денервация , факторы роста нервов , регенерация нервов , нейробиология , Повреждения периферических нервов , восстановление функций
ВВЕДЕНИЕ
Повреждения нервов запускают сложные клеточно-молекулярные взаимодействия, которые необходимы для регенерации аксонов и, как следствие, для значимого функционального восстановления пациентов.Эти клеточные и молекулярные ответы ненейрональных клеток периферической нервной системы (ПНС), а именно шванновских клеток (СК), создают благоприятную для роста среду для поврежденных аксонов, в отличие от реакции центральной нервной системы (ЦНС) на повреждения. Однако существует дихотомия между способностью поврежденных нейронов ПНС регенерировать свои аксоны после повреждения и субоптимальным восстановлением функции после повреждения нервов.
Значительные успехи были достигнуты в технике микрохирургии поврежденных нервов, что иногда приводит к улучшению результатов для пациентов. 1-3 К сожалению, функциональное восстановление после травм периферических нервов часто остается слабым, особенно когда повреждены нервные стволы, расположенные близко к спинному мозгу и далеко от органов-мишеней. Эти травмы включают большинство травм отрывного типа, разрывов нервов и ушибов нервов. После этих типов нервных повреждений поврежденные нейроны должны регенерировать свои аксоны на больших расстояниях с очень медленной скоростью 1 мм / день. При такой медленной скорости регенерации восстановление функциональной двигательной единицы или реиннервация органа чувств может занять месяцы или даже годы ( 1-4 ), состояние, называемое хронической аксотомией. 5-7 Аналогичным образом, SC в дистальных обрубках нерва и в органах-мишенях остаются денервированными в течение длительных периодов, состояний, известных как хроническая SC денервация 2,8-11 и хроническая денервация мышц 11 соответственно.
Схематическое изображение повреждений проксимального нерва плечевого сплетения, которые требуют длительного времени регенерации регенерирующих аксонов для реиннервации мышц руки. В это время поврежденные нейроны подвергаются хронической аксотомии, а шванновские клетки дистальной культи поврежденных нервов хронически денервированы.Графики на вставках демонстрируют прогрессирующее снижение способности поврежденных мотонейронов, которые регенерируют аксоны (число MN) и реиннервируют мышцы (число двигательных единиц [MU]), в результате хронической денервации шванновских клеток и хронической аксотомии нейронов.
Нарушение функционального восстановления кисти, например, обычно объясняется необратимой атрофией денервированных мышц и органов чувств и замещением жировой ткани. 1 Однако наши эксперименты за последние ~ 15 лет предоставляют убедительные доказательства того, что прогрессирующая неспособность хронически аксотомированных нейронов регенерировать свои аксоны и прогрессирующая неспособность хронически денервированных SCs поддерживать регенерирующие аксоны играют преобладающую роль в клинических неудачах функциональное восстановление после микрохирургического восстановления. 2,5,7,8,10-18 Плохое функциональное восстановление также усугубляется задержкой микрохирургического восстановления поврежденных нервов, когда пациенты наблюдаются для спонтанного функционального восстановления, обычно в течение 3-6 месяцев после травм тупым нервом. Регенерация аксонов в неправильные эндоневральные трубки и конечные органы — так называемое неправильное направление регенерирующих аксонов — является еще одним фактором, который способствует плохому функциональному восстановлению.
РЕГЕНЕРАЦИОННЫЙ РЕАКЦИЯ ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И ЕГО РОЛЬ В ФУНКЦИОНАЛЬНОМ РЕЗУЛЬТАТЕ
Начальная фаза регенерации аксонов
После повреждения нерва проксимальная и дистальная культя поврежденного нерва претерпевают структурные и молекулярные изменения, готовясь к процессу аксональной регенерации.Проксимальная культя претерпевает обратную дегенерацию до первого узла Ранвье, а затем каждый поврежденный аксон вырабатывает несколько дочерних аксонов. 19,20 Многие дочерние аксоны обрезаются; те, что остались, начинают процесс удлинения через дистальные культи нерва и составляют регенерирующие единицы. 21 Эта начальная стадия регенерации аксонов поддерживается как доступностью локально продуцируемых цитоскелетных материалов, так и продуцируемыми нейронами и транспортируемыми антероградно цитоскелетными белками, такими как актин и тубулин (A). 6,20,22,23 Регенерация аксонов протекает со скоростью 1-3 мм / сут, что соответствует медленной скорости транспорта цитоскелетных материалов. Дальнейшее удлинение и регенерация через дистальную культю нерва зависят от среды, поддерживающей рост, обеспечиваемой SC дистальных культей нерва. Отсутствие эндоневриальных каналов, нагруженных шванновскими клетками (полосы Бюнгнера), приводит к неправильной регенерации и образованию невром. 3,24-26
После повреждения нерва гены, связанные с регенерацией (RAG), временно активируются в нейронах, в то время как гены, связанные с нормальной синаптической передачей, подавляются (A и B).Шванновские клетки денервированных обрубков нерва подвергаются пролиферации во время дегенерации Валлера и экспрессируют множество RAG, в то время как миелин-ассоциированные гены подавляются (A). Профили генов поддерживают рост аксонов, но экспрессия очень недолговечна, так что со временем (в то время как регенерирующие аксоны растут с медленной скоростью 1 мм / сут) экспрессия RAG снижается, и способность поврежденных нейронов регенерировать свои аксонов и шванновских клеток, поддерживающих регенерацию, уменьшается (C).Примеры прогрессивного снижения уровней мРНК нанесены на графики для тубулина в нейронах и GDNF в шванновских клетках.
Роль шванновских клеток в регенерации аксонов
Дистальные культи перерезанных нервов подвергаются дегенеративному процессу, названному в честь Августа Валлера, валлеровской дегенерации. 27 Теперь мы понимаем, что валлеровская дегенерация является важным подготовительным этапом процесса регенерации аксонов, посредством которого удаляются молекулы, которые могут препятствовать регенерации.В недавнем обзоре события разбиты на три морфологически различимые стадии: острая дегенерация аксонов, латентный период и стадия резкой гранулярной дегенерации. 28 В течение часа после травмы отмирание как в проксимальной, так и в дистальной культях опосредуется каналом-опосредованным притоком кальция и активацией кальпаинов, которые расщепляют нейрофиламенты и связанные с микротрубочками компоненты, такие как спектрин и тубулин. В проксимальной культи эта перемычка относится к первому узлу Ранвье. Следующим этапом примерно в 24 часа, в течение которого продолжаются измерения аксонных потенциалов, 29 следует более длительный период разборки белков цитоскелета кальцием-активированными кальпаинами и убиквитин-протеасомной системой. 30 СК играют главную роль в этом процессе посредством фагоцитоза аксонов и миелиновых остатков. Они также секретируют хемоаттрактивные факторы, такие как интерлейкин-1 и хемоаттрактантный белок-1 моноцитов, которые рекрутируют макрофаги в денервированные дистальные культи нервов, которые вносят значительный вклад в фагоцитоз аксонов и миелиновых остатков. 23,31,32 Обломки аксонов высвобождают митогены, которые способствуют митотическому делению SC и инициируют сеть цитокинов и факторов транскрипции, которые стимулируют распад миелина и инвазию макрофагов и нацелены на миелин для фагоцитоза SC и макрофагами. 6,33
Преобразование из режима действия-потенциала-проводимости в режим, поддерживающий регенерацию: роль генов, связанных с регенерацией
Сразу после повреждения нерва потеря аксонального контакта запускает пролиферацию СК и их фенотип от миелинизирующего к немиелинизирующему фенотипу, поддерживающему рост. 6,23,34 Экспрессия матричной РНК (мРНК) миелин-ассоциированных белков, таких как P0 и миелин-ассоциированный гликопротеин, подавляется.Нейротрофины (например, фактор роста нервов, нейротрофический фактор головного мозга и нейротрофический фактор глиального происхождения), их рецепторы (например, p75, GFRA-1, GFRA-2) и молекулы адгезии (например, молекула адгезии нервных клеток) активируются при подготовке к процессу регенерации аксонов (). 6,26,35,36 Эти гены с усиленной регуляцией в совокупности называются генами, связанными с регенерацией (RAG). Изменение экспрессии генов, дегенерация и клиренс миелина и аксонов являются ключевыми особенностями процесса валлеровской дегенерации. 6,23,34
Точно так же нейроны, чьи нервы были повреждены, подавляют мРНК белков, необходимых для нейротрансмиссии, и активируют их для восстановления своих периферических процессов. 6,12,37 Следовательно, актин, тубулин и GAP-43 активируются сразу после повреждения (A и B). 6,37 Однако активация RAG не поддерживается ни в поврежденных нейронах, ни в SC. К 6 месяцам у экспериментальных животных подавляющая регуляция большинства мРНК с активизацией снижается, что приводит к потере среды, поддерживающей рост, для регенерации аксонов (A и B). 6,37 Значение ограниченной по времени активации RAG продемонстрировано в прогрессирующем снижении способности поврежденных нейронов регенерировать свои аксоны и SCs поддерживать регенерирующие аксоны по мере увеличения продолжительности восстановления нерва ().
Регенеративная способность снижается со временем из-за продолжительной аксотомии и денервации шванновских клеток. На крысах мы экспериментально увеличили либо (А) продолжительность времени, в течение которого мотонейроны не могли регенерировать свои аксоны (хроническая аксотомия; путем отсрочки наложения швов от культи проксимального нерва к недавно денервированной культе дистального нерва), либо (В) денервации. клеток Шванна в дистальных обрубках нерва (хроническая денервация; путем отсрочки наложения швов свежеразрезанной проксимальной культи нерва на культю нерва, которая была хронически денервирована до восстановления нерва).Способность нейронов к регенерации после хронической аксотомии мотонейронов или хронической денервации шванновских клеток и мышц-мишеней определялась либо путем (а) расчета количества реиннервируемых мышечных единиц (нервов и мышечных волокон, которые снабжает один мотонейрон) с использованием силы измерения в ответ на стимуляцию одиночных аксонов и мышечного нерва или (б) подсчет мечения мотонейронов, которые успешно регенерировали свои аксоны, путем нанесения ретроградного красителя на регенерирующие аксоны в культе нерва дистальнее места восстановления нерва.Оценки регенеративного успеха, полученные с помощью методов подсчета мотонейронов и подсчета двигательных единиц, хорошо согласуются, демонстрируя прогрессирующее снижение регенеративной способности как функцию (C) хронической аксотомии мотонейронов и (D) хронической денервации шванновских клеток. .
ЭКСПЕРИМЕНТАЛЬНЫЕ ПАРАДИГМЫ И ОЦЕНКА РЕГЕНЕРАЦИИ АКСОНА ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И МИКРОХИРУРГИЧЕСКОГО РЕМОНТА
Экспериментальные исследования, проведенные во время Второй мировой войны Гутманном и Янгом, показали, что замедленное восстановление нерва не влияет на скорость роста и количество аксонов. 38,39 Эти данные, свидетельствующие о крайней атрофии денервированных мышц, привели к ошибочному выводу, что плохое функциональное восстановление после повреждения нерва было связано с необратимой денервационной атрофией мышцы и ее неспособностью воспринимать иннервацию, особенно после длительных периодов воздействия. время. 38,39 Этот вывод стал довольно популярным и, к сожалению, часто повторяется даже в последних публикациях. Недавние данные нашей лаборатории и других источников не подтвердили ошибочный вывод, главным образом потому, что (1) авторы не оценили напрямую количество поврежденных нейронов, которые регенерировали в дистальную культи нерва, и тех, которые реиннервируют денервированные мышцы, и (2) подсчитали нервные волокна в дистальной культе. Культя нерва переоценивает количество регенерированных нервных волокон, потому что многие дочерние аксоны возникают из единичных регенерирующих нервных волокон в проксимальной культе нерва и могут ошибочно считаться репрезентативными для отдельных нервных волокон в проксимальной культи.
В нескольких экспериментах мы изучали процесс регенерации аксонов после немедленного и отсроченного восстановления повреждения нерва. Кроме того, мы использовали количественные методы подсчета мотонейронов, которые регенерировали свои аксоны в дистальные обрубки нервов, и подсчета количества реиннервируемых моторных единиц в целевых мышцах, чтобы оценить способность мотонейронов регенерировать свои аксоны и реиннервировать мышцы. Используя метод перекрестного шва на крысах, который позволяет независимо изучить эффекты отсроченной реиннервации дистальной культи нерва (так называемая хроническая или длительная денервация) и отсроченной регенерации нейронов до их мишеней (так называемая хроническая или длительная аксотомия), мы оценили количество регенерированных мотонейронов с использованием техники обратной мечения, при которой ретроградно транспортируемые флуоресцентные красители наносятся на срезанные концы реиннервируемых дистальных культей нерва, которые маркируют мотонейроны, регенерировавшие свои аксоны.Таким образом, мы смогли подсчитать количество мотонейронов, которые регенерировали свои аксоны в вентральном роге спинного мозга после наших окончательных экспериментов. Мы оценили количество двигательных единиц, применяя сверхмаксимальную стимуляцию к отдельным вентральным корешкам и измеряя общую мышечную силу. Затем мы выделили волокна вентрального корня и стимулировали их индивидуально, чтобы определить силу двигательных единиц. Затем количество реиннервируемых двигательных единиц рассчитывали путем деления мышечной силы на силу двигательных единиц.иллюстрирует нашу хирургическую парадигму и методы оценки регенерации аксонов и реиннервации мышц после повреждения и восстановления нерва.
Хроническая денервация шванновских клеток
Предоставление SCs среды, поддерживающей рост, тесно связано с потерей и своевременным восстановлением контакта аксонов с клетками. 10 SC, реиннервация которых задерживается регенерирующими аксонами, считаются хронически денервированными, и пагубное влияние хронической денервации на SC ухудшается по мере увеличения продолжительности хронической денервации. 2,10,18 В наших экспериментах, чтобы проверить эффект хронической денервации, общая малоберцовая (CP) ветвь седалищного нерва перерезается, и ее реиннервация откладывается на период от 1 до 6 месяцев до наложения поперечного шва. проксимальная культя свежеперерезанного большеберцового (TIB) нерва в хронически денервированный дистальный CP-нерв. Таким образом, можно непосредственно оценить влияние хронической денервации ЦП на регенерацию свежеповрежденных мотонейронов TIB. Используя эту парадигму, хроническая денервация уменьшила количество мотонейронов, которые были обратно помечены фторорубовым красителем, который был нанесен на дистальную культи нерва в 10 мм от места перекрестного шва, до менее чем 10% от количества, которое регенерировалось после немедленного ушивания культи нервов. ().Было превосходное соответствие между этой долей мотонейронов, которые регенерировали свои аксоны в хронически денервированную культю нерва, и долей недавно аксотомированных мотонейронов, которые регенерировали и реиннервировали денервированную мышцу через 4-6 месяцев (). 10 Уменьшение количества мотонейронов, которые регенерируют свои аксоны через хронически денервированные SC, не только реиннервирует денервированные мышечные волокна, но и реиннервирует в 3 раза больше мышечных волокон, чем обычно.Поскольку это увеличение реиннервируемых двигательных единиц составляет максимальную способность к отрастанию мотонейронов, 2,10 , отсюда следует, что неспособность хронически денервированных мышечных волокон принять реиннервацию ограничивает функциональное восстановление после хронических нервных повреждений. 2,10,23 Устойчивая способность денервированной мышцы принимать реиннервацию также была продемонстрирована параллельным восстановлением как мышечной массы, так и изометрической силы. Тем не менее, изоляция эффектов денервации SC от денервации мышц показала, что хронически денервированные мышечные волокна не полностью восстановили свой прежний размер, утверждая, что ограниченное количество сателлитных клеток включается в каждое атрофическое мышечное волокно для восстановления площади поперечного сечения мышечных волокон. 11 Следовательно, прогрессирующее ухудшение способности SCs поддерживать рост в дистальных культях нерва играет главную роль в плохом функциональном восстановлении после повреждения нерва, а роль мышечной атрофии вторична. Поразительно, что долгоживущие хронически денервированные SC сохраняли свою способность ремиелинизировать меньшее количество регенерированных аксонов, 2 , особенно с учетом прогрессирующей регрессии способности денервированных SC поддерживать свой разрешающий рост фенотип и прогрессирующее снижение. в количестве поддерживающих рост SC в хронически денервированных культях дистального нерва. 36,40
Клиническим коррелятом нашей экспериментальной парадигмы перекрестного шва являются повреждения больших культи нерва, при которых СК дистальных культей нерва остаются хронически денервированными из-за медленной скорости регенерации аксонов. Чем больше времени требуется регенерирующим аксонам для реиннервации СК в дистальных культях нерва, тем дольше длится период хронической денервации, в течение которого СК не контактируют с аксонами, и тем выше вероятность того, что их способность поддерживать регенерирующие аксоны нарушена. 2,6,10 Мы считаем, что отчасти поэтому реиннервация и последующее функциональное восстановление в мышечных мишенях, расположенных рядом с регенерирующими мотонейронами, лучше, чем в более дистально расположенных мышечных мишенях.
Хроническая аксотомия нейронов
Поврежденные нейроны, аксоны которых могут или не могут регенерироваться, но не установили целевые соединения, подвергаются хронической аксотомии. 5,12,14 Например, после повреждения плечевого сплетения поврежденные нейроны должны регенерироваться на большом расстоянии, прежде чем они смогут реиннервировать некоторые денервированные мышцы и, таким образом, подвергнуться хронической аксотомии.Чтобы проверить эффект хронической аксотомии, мы также использовали технику перекрестного шва с использованием 2 ветвей седалищного нерва крысы. Ветвь большеберцовой кости седалищного нерва перерезается, и ее регенерация задерживается на периоды от 1 до 6 месяцев до наложения поперечного шва проксимальной культи хронически аксотомированной проксимальной культи большеберцового нерва в только что разрезанный дистальный центральный нерв. Таким образом, можно напрямую оценить влияние хронической аксотомии TIB на реиннервацию только что поврежденного дистального CP-нерва. Используя эту парадигму и показатели результатов, описанные выше, мы обнаружили, что количество мотонейронов, регенерировавших свои аксоны, постепенно снижалось в зависимости от продолжительного времени хронической аксотомии до ~ 37% от тех, которые регенерировали без эффекта хронической аксотомии. 5,7 Это снижение способности к регенерации после хронической аксотомии является значительным, особенно в сочетании с пагубным эффектом хронической денервации, которые возникают одновременно, особенно после травм крупных нервных стволов, таких как плечевое сплетение. Прогрессирующее снижение регенеративной способности хронически аксотомированных мотонейронов сопровождалось снижением активированных RAG. 41 Драматический негативный эффект снижения регенерационной способности хронически аксотомированных мотонейронов не повлиял на способность нейронов реиннервировать в 3 раза больше мышечных волокон, чем обычно, меньшее количество мотонейронов увеличивало свои двигательные единицы и количество мышечных волокон. снабжается каждым реиннервирующим мотонейроном. 5 Таким образом, хроническая аксотомия нарушает регенеративную способность мотонейронов, но не снижает способность меньшего числа регенерированных моторных аксонов устанавливать функциональные связи с денервированными мышечными волокнами.
Шахматная регенерация аксонов и неправильное направление регенерирующих аксонов
Одним из ключевых условий успешного функционального восстановления является то, что регенерирующие аксоны регенерируют в правильные эндоневральные трубки, которые направляют их обратно к исходным органам-мишеням.Однако мы обнаружили, что регенерирующие аксоны сталкиваются со значительной задержкой в месте повреждения и проходят через место повреждения в дистальные культи нерва в шахматном порядке (шахматная регенерация аксонов). Действительно, пересечение регенерирующих аксонов происходит медленно до входа регенерирующих аксонов в дистальные обрубки нерва. Только после того, как аксоны попадают в дистальную культю, они регенерируют с медленной скоростью транспорта 1-3 мм / сут.
Функциональные результаты после травм аксонов являются наилучшими после травм с раздавливанием нерва, при которых эндоневрий остается анатомически неповрежденным на всем пути к цели, а отростки аксонов регенерирующего блока содержатся в исходном эндоневрии, поэтому регенерирующие аксоны возвращаются в исходное положение. цели. 3 Со временем реиннервируемые мышечные волокна демонстрируют нормальное мозаичное распределение типов мышечных волокон, двигательные единицы и изометрические силы мышц полностью восстанавливаются, первоначальное количество функциональных двигательных единиц восстанавливается, а количество и диаметры миелинизированных нервных волокон возвращаются к прежним значениям. обычный. 42
Повреждения нервов, приводящие к повреждению эндоневральных трубок (например, аксонотметические или нейротметические повреждения), в результате которых регенерирующие аксоны должны найти свои исходные эндоневральные трубки, представляют собой потенциальную возможность ошибок в процессе регенерации.Отростки аксонов, исходящие из культи проксимального нерва, могут попадать в несколько различных эндоневральных трубок с целевыми точками, которые они раньше не обеспечивали, как в случае моторных аксонов, которые регенерируют в путях, ведущих к коже, а не к мышцам. 43 Эксперименты, в которых ретроградные красители применялись к регенерированным аксонам перерезанных седалищных и лицевых нервов у крыс, напрямую продемонстрировали неправильное направление регенерирующих моторных аксонов от нескольких различных пулов мотонейронов к мышцам, которые мотонейроны ранее не снабжали. 44 В случаях, когда нервы снабжали мышцы антагонистическими действиями, неправильное направление аксонов было связано с несоответствующими движениями, что вполне согласуется с данными о том, что мотонейроны сгибателей и разгибателей поддерживают свой нормальный, но несоответствующий паттерн возбуждения, когда они направлены на ненадлежащую реиннервацию мышц-разгибателей и сгибателей. , соответственно. 45 Мы считаем, что это неправильное направление регенерирующих поврежденных аксонов играет важную роль в снижении функционального восстановления после травм нервов.
Есть некоторые исключения из неправильного направления регенерирующих аксонов даже после травмы транзакции. К ним относятся несколько периферических нервных стволов, ветви которых иннервируют скелетные мышцы и органы чувств в коже и суставах. Примеры включают бедренный нерв с двумя ветвями, мышечную ветвь, содержащую двигательные и сенсорные нервы к четырехглавой мышце, и другую чисто сенсорную ветвь подкожного нерва к коже. 43 Хотя реиннервация моторных и сенсорных нервных ветвей изначально случайна, мотонейроны, которые постепенно регенерируют свои аксоны через место шва, посылают свои аксоны в соответствующую моторную ветвь. 43,46 Эта преимущественная реиннервация моторики возникла параллельно с прогрессирующей или ступенчатой регенерацией моторных аксонов в дистальных обрубках нерва. 46 Необычный кислый гликан, связанный с миелиновыми профилями моторных, но не сенсорных аксонов мыши, который распознается моноклональным антителом L2 / HNK-1, может быть одним из механизмов предпочтительной реиннервации соответствующих путей. 47,48 Дифференциальный профиль нейротрофических факторов в SCs, происходящих из сенсорных и моторных аксонов с профилем BDNF / NT4 / 5, также, вероятно, играет важную роль в предпочтительной реиннервации моторных путей. 49
ВЫВОДЫ
Огромный прогресс был достигнут в нашем понимании микроанатомии, патофизиологии и микрохирургического лечения поврежденных нервов. Эти достижения улучшили качество ухода за пациентами с повреждениями нервов и часто приводят к лучшему функциональному восстановлению. Однако пациенты, у которых не удается восстановить хорошую функцию, несмотря на превосходный микрохирургический уход, представляют собой проблему для нервного хирурга. Повышенная регуляция RAG недолговечна, и, по-видимому, существует временное окно возможностей, в течение которого SCs обеспечивают среду, поддерживающую рост, и поврежденные нейроны могут регенерировать свои аксоны.Ограниченная по времени повышающая регуляция RAG и медленная скорость регенерации аксонов приводят к прогрессирующей потере нейротрофической поддержки поврежденных нейронов и их регенерирующих аксонов, отсюда хроническая денервация SC и хроническая аксотомия нейронов. 5,6,23 Поэтому в нашей лаборатории в Ochsner мы фокусируемся на 2 возможных подходах к борьбе с разрывом между временем активации RAG и медленной скоростью регенерации, включая экспериментальные стратегии, которые ускоряют скорость регенерации аксонов и поддерживают нейротрофическая среда для регенерации аксонов в течение более длительных периодов времени.Мы продемонстрировали положительный эффект кратковременной (1 ч) низкочастотной электростимуляции на ускорение регенерации аксонов за счет модуляции экспрессии RAG. 46,50-52 Мы также показали положительный эффект трансформации фактора роста бета в реактивации и поддержании экспрессии RAGS хронически денервированными SC. В настоящее время мы изучаем роль трансплантации стволовых клеток в дистальные обрубки нерва и использование усиленных нервных направляющих каналов или проводников, которые не только устранят необходимость в аутотрансплантатах, но также позволят позитивно модулировать среду, способствующую росту аксонов. прохождение направляющих каналов в дистальные культи нерва. 53,54
Мы считаем, что окончательное решение для улучшения функционального восстановления в подгруппе пациентов, которые в настоящее время не справляются с микрохирургическим вмешательством, представляет собой комбинацию нескольких нейробиологических подходов для преодоления проблемы ограниченного временного окна для оптимального восстановления нерва. регенерация.
Сноски
Авторы не имеют финансовой или имущественной заинтересованности в предмете данной статьи .
Эта статья соответствует требованиям Совета по аккредитации для последипломного медицинского образования и Американского совета по медицинским специальностям, поддерживающим сертификационные компетенции по уходу за пациентами и медицинским знаниям.
СПИСОК ЛИТЕРАТУРЫ
1. Сулейман О.А.Р., Мидха Р., Гордон Т. Патофизиология хирургических нервных расстройств. В: Winn HR, редактор. Юманс Неврологическая хирургия . 6-е изд. Филадельфия, Пенсильвания: Сондерс ;; 2011. С. 2368–2379. В. изд. [Google Scholar] 2. Сулейман О.А., Гордон Т. Влияние краткосрочной и долгосрочной денервации шванновских клеток на регенерацию периферических нервов, миелинизацию и размер. Глия . 2000 декабрь; 32 (3): 234–246. [PubMed] [Google Scholar] 3. Сандерленд С. Нервы и травмы нервов .Эдинбург, Шотландия: Черчилль Ливингстон ;; 1978. [Google Scholar] 4. Сулейман В.А., Клайн Д.Г. Хирургия нервов: обзор и взгляд на будущее. Clin Neurosurg . 2006; 53: 38–47. [PubMed] [Google Scholar] 5. Fu SY, Gordon T. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная аксотомия. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3876–3885. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Фу С.Ю., Гордон Т. Клеточные и молекулярные основы регенерации периферических нервов. Мол Нейробиол . 1997, февраль-апрель; 14 (1-2): 67–116. [PubMed] [Google Scholar] 7. Бойд Дж. Г., Гордон Т. Дозозависимое облегчение и ингибирование регенерации периферических нервов нейротрофическим фактором мозга. евро J Neurosci . 2002 Февраль; 15 (4): 613–626. [PubMed] [Google Scholar] 8. Сулейман О.А., Мидха Р., Манро К.А. и др. Хроническая денервация шванновских клеток и наличие сенсорного нерва снижают регенерацию моторных аксонов. Опыт Нейрол . 2002 август; 176 (2): 342–354. [PubMed] [Google Scholar] 9.Sulaiman OA, Voda J, Gold BG, Gordon T. FK506 увеличивает регенерацию периферических нервов после хронической аксотомии, но не после хронической денервации шванновских клеток. Опыт Нейрол . 2002 Май; 175 (1): 127–137. [PubMed] [Google Scholar] 10. Фу С.Ю., Гордон Т. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная денервация. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3886–3895. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Гордон Т., Тайреман Н., Раджи М.А. Основа для снижения функционального восстановления после отсроченного восстановления периферических нервов. Дж. Neurosci . 2011 6 апреля; 31 (14): 5325–5334. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Бойд Дж. Г., Гордон Т. Нейротрофические факторы и их рецепторы в регенерации аксонов и функциональном восстановлении после повреждения периферических нервов. Мол Нейробиол . 2003 июн; 27 (3): 277–324. [PubMed] [Google Scholar] 13. Сулейман О.А., Гордон Т. Трансформирующий фактор роста-бета и форсколин ослабляют неблагоприятные эффекты долгосрочной денервации шванновских клеток на регенерацию периферических нервов in vivo. Глия . 2002, 1 марта; 37 (3): 206–218. [PubMed] [Google Scholar] 14. Гордон Т., Фу С.Ю. Долгосрочная реакция на повреждение нерва. Adv Neurol . 1997. 72: 185–199. [PubMed] [Google Scholar] 15. Boyd JG, Gordon T. Нейротрофический фактор, полученный из линии глиальных клеток, и нейротрофический фактор, полученный из мозга, поддерживают регенерацию аксонов хронически аксотомированных мотонейронов in vivo. Опыт Нейрол . Октябрь 2003 г., 183 (2): 610–619. [PubMed] [Google Scholar] 16. Sulaiman OAR, Gordon T. TGF-бета обращает пагубный эффект длительной денервации шванновских клеток на регенерацию нервов, индуцируя экспрессию рецептора erbB3. Глия . 2003 сентябрь; 43 (Дополнение 2): 24. [Google Scholar] 17. Midha R, Munro CA, Chan S, Nitising A, Xu QG, Gordon T. Регенерация в защищенные и хронически денервированные культи периферических нервов. Нейрохирургия . 2005 декабрь; 57 (6): 1289–1299. [PubMed] [Google Scholar] 18. Сулейман О.А., Гордон Т. Роль хронической денервации шванновских клеток в плохом функциональном восстановлении после нервных повреждений и экспериментальные стратегии борьбы с ней. Нейрохирургия . Октябрь 2009 г., 65 (4 доп.): A105 – A114. [PubMed] [Google Scholar] 19.Cajal SR. Дегенерация и регенерация нервной системы . Оксфорд, Великобритания: Издательство Оксфордского университета; 1928. Перевод Р. М. Мэй. [Google Scholar] 20. Эртюрк А., Хеллал Ф., Энес Дж., Брадке Ф. Дезорганизованные микротрубочки лежат в основе образования ретракционных луковиц и нарушения регенерации аксонов. Дж. Neurosci . 22 августа 2007 г.; 27 (34): 9169–9180. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Моррис Дж. Х., Хадсон А. Р., Уэдделл Г. Исследование дегенерации и регенерации разделенного седалищного нерва крысы на основе электронной микроскопии.II. Развитие «регенерационного агрегата. Z Zellforsch Mikrosk Anat . 1972: 124 (1): 103–130. [PubMed] [Google Scholar] 22. McQuarrie IG. Влияние кондиционирующего поражения на формирование аксонального отростка в узлах Ранвье. Дж. Комп. Neurol . 1985, 8 января; 231 (2): 239–249. [PubMed] [Google Scholar] 23. Сулейман О.А., Бойд Дж. Г., Гордон Т. Регенерация аксонов в периферической системе млекопитающих. В: Kettenmann H, Ransom BR, editors. Нейроглия . 2-е изд. Оксфорд, Великобритания: Издательство Оксфордского университета; 2005 г.С. 454–466. В. ред. [Google Scholar] 24. Эйткен Дж. Т., Шарман М., Янг Дж. З. Созревание периферических нервных волокон с различными периферическими связями. Дж Анат . 1947; 81: 1–22. [PubMed] [Google Scholar] 25. Toft PB, Fugleholm K, Schmalbruch H. Разветвление аксонов после раздавливания периферических нервов крысы. Мышечный нерв . 1988 августа; 11 (8): 880–889. [PubMed] [Google Scholar] 26. Наварро X, Виво М., Валеро-Кабре А. Нейропластичность после повреждения и регенерации периферических нервов. Прог Нейробиол . Июль 2007 г .; 82 (4): 163–201. Epub 2007 22 июня. [PubMed] [Google Scholar] 27. Уоллер А. Эксперименты на разрезе языкоглоточного и подъязычного нервов лягушки и наблюдения за изменениями, вызванными этим в структуре их примитивных волокон. Philos Trans R Soc London . 1850; 140: 423–429. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Ван Дж.Т., Медресс З.А., Баррес Б.А. Дегенерация аксонов: молекулярные механизмы пути самоуничтожения. Дж. Ячейка Биол . 2012 9 января; 196 (1): 7–18. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Чжай К., Ван Дж., Ким А. и др. Участие убиквитин-протеасомной системы на ранних стадиях валлеровской дегенерации. Нейрон . 17 июля 2003 г.; 39 (2): 217–225. [PubMed] [Google Scholar] 31. Сулейман О.А., Клеточный Гордон Т. и молекулярные взаимодействия после повреждения периферических и центральных нервов. Биомед Ред. . 2003. 14: 51–62. [Google Scholar] 32. Хёке А., Ми Р. В поисках новых методов лечения периферических невропатий и регенерации нервов. Дисков Мед . 2007 август; 7 (39): 109–112. [PubMed] [Google Scholar] 33. Brushart TM. Ремонт нервов . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета ;; 2011. [Google Scholar] 34. Scherer SS, Salzer JL. Аксон-шванновские клеточные взаимодействия во время дегенерации и регенерации периферических нервов. В: Jessen KR, Richardson WD, редакторы. Развитие глиальных клеток: основные принципы и клиническая значимость . Оксфорд, Великобритания: Bios Scientific ;; 1996. С. 169–196. В. ред. [Google Scholar] 35. Хёке А., Гордон Т., Зочодне Д.В., Сулейман О.А.Снижение экспрессии нейротрофического фактора, происходящего из глиальных клеток, связано с нарушением регенерации после длительной денервации шванновских клеток. Опыт Нейрол . 2002 Янв; 173 (1): 77–85. [PubMed] [Google Scholar] 36. You S, Petrov T, Chung PH, Gordon T. Экспрессия рецептора фактора роста нервов с низким сродством в долгосрочно денервированных шванновских клетках. Глия . 1997 июн; 20 (2): 87–100. [PubMed] [Google Scholar] 37. Бисби М.А., Тецлафф В. Изменения в синтезе цитоскелетного белка после повреждения аксона и во время регенерации аксона. Мол Нейробиол . 1992; 6 (2-3): 107–123. Лето осень. [PubMed] [Google Scholar] 38. Гутманн Э. Влияние задержки иннервации на восстановление мышц после нервных поражений. Дж. Нейрофизиол . 1948 июл; 11 (4): 279–294. [PubMed] [Google Scholar] 40. Ли Х, Теренги Г, зал СМ. Влияние отсроченной реиннервации на экспрессию рецепторов c-erbB хронически денервированными шванновскими клетками крысы in vivo. Глия . 1997 августа; 20 (4): 333–347. [PubMed] [Google Scholar] 41. Тецлафф В., Леонард С., Крекоски К.А., Пархад И.М., Бисби Массачусетс.Снижение синтеза мотонейрональных нейрофиламентов путем последовательных аксотомий: возможное объяснение эффекта кондиционирующего поражения на регенерацию аксонов. Опыт Нейрол . 1996 Май; 139 (1): 95–106. [PubMed] [Google Scholar] 42. Гордон Т., Паттулло М.С. Пластичность мышечных волокон и типов двигательных единиц. Exerc Sport Sci Ред. . 1993; 21: 331–362. [PubMed] [Google Scholar] 44. Brushart TM, Mesulam MM. Трансганглионарная демонстрация центральных сенсорных проекций кожи и мышц с конъюгатами HRP-лектин. Neurosci Lett . 1980 апр; 17 (1-2): 1–6. [PubMed] [Google Scholar] 45. Гордон Т, Стейн РБ, Томас СК. Организация двигательных единиц после перекрестной реиннервации мышц-антагонистов в задней конечности кошки. Дж. Физиология . 1986 Май; 374: 443–456. [Бесплатная статья PMC] [PubMed] [Google Scholar] 46. Аль-Маджед А.А., Нойманн С.М., Брушарт TM, Гордон Т. Кратковременная электрическая стимуляция повышает скорость и точность регенерации моторных аксонов. Дж. Neurosci . 2000 г., 1 апреля; 20 (7): 2602–2608.[Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Мартини Р., Шахнер М., Брушарт TM. Углевод L2 / HNK-1 предпочтительно экспрессируется шванновскими клетками, ранее ассоциированными с моторными аксонами, в реиннервируемых периферических нервах. Дж. Neurosci . 1994, ноябрь; 14 (11, часть 2): 7180–7191. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Эберхардт К.А., Иринчев А.А., Аль-Маджед А.А. и др. Передача сигналов BDNF / TrkB регулирует экспрессию углеводов HNK-1 в регенерирующих двигательных нервах и способствует функциональному восстановлению после восстановления периферических нервов. Опыт Нейрол . 2006 Апрель; 198 (2): 500–510. Epub 2006 7 февраля. [PubMed] [Google Scholar] 49. Хёке А., Редетт Р., Хамид Х и др. Шванновские клетки выражают моторные и сенсорные фенотипы, которые регулируют регенерацию аксонов. Дж. Neurosci . 2006 20 сентября; 26 (38): 9646–9655. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Al-Majed AA, Brushart TM, Gordon T. Электрическая стимуляция ускоряет и увеличивает экспрессию мРНК BDNF и trkB в регенерирующих мотонейронах бедренной кости крысы. евро J Neurosci .2000 декабрь; 12 (12): 4381–4390. [PubMed] [Google Scholar] 51. Brushart TM, Hoffman PN, Royall RM и др. Электрическая стимуляция способствует регенерации мотонейрона без увеличения его скорости и без кондиционирования нейрона. Дж. Neurosci . 2002, 1 августа; 22 (15): 6631–6638. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Гордон Т., Сулейман О.А., Ладак А. Глава 24: Электростимуляция для улучшения регенерации нервов: где мы находимся? Международная версия Neurobiol . 2009. 87: 433–444. [PubMed] [Google Scholar] 53.Мидха Р. Новые методы восстановления нервов: нервные передачи и нервные трубки. Clin Neurosurg . 2006. 53: 185–190. [PubMed] [Google Scholar] 54. Пфистер Л.А., Папалоизос М., Меркл Г.П., Гандер Б. Нервные проводники и доставка факторов роста при восстановлении периферических нервов. J Peripher Nerv Syst . 2007 июн; 12 (2): 65–82. [PubMed] [Google Scholar]
От лабораторных исследований до прикроватных приложений
Охснер Дж. 2013 Весна; 13 (1): 100–108.
, MD, PhD, FRCS * и, PhD †
Wale Sulaiman
* Отделение нейрохирургии, Ochsner Clinic Foundation, и Медицинская школа Университета Квинсленда, Клиническая школа Охснера, Новый Орлеан, Луизиана
Тесса Гордон
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
* Отделение нейрохирургии, Фонд клиники Окснера и Школа медицины Университета Квинсленда, Клиническая школа Окснера, Новый Орлеан, Луизиана
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
Адресная корреспонденция Wale Sulaiman, MD, PhD, FRCS, Отделение нейрохирургии, Ochsner Clinic Foundation, 1514 Jefferson Hwy., New Orleans, LA 70121. Тел: (504) 842-2181 Факс: (504) 842-6531 Электронная почта: gro.renshco@namialusw Авторские права © Академическое подразделение Фонда клиники Окснера Эта статья цитируется в других статьях в PMC.
РЕФЕРАТ
Объективы
Мы рассматриваем современную нейробиологию повреждения и регенерации нервов, особенно в том, что касается восстановления полезной функции у пациентов, которые получили травмы крупных нервных стволов, таких как плечевое сплетение.
Методы
В этом обзоре основное внимание уделяется исследованиям, проведенным в нашей лаборатории в Окснеру и в других лабораториях, связанных с нейробиологией повреждения нервов, с акцентом на то, как некоторые из ключевых результатов исследований на животных помогают нам понять патофизиологию плохого функционального восстановления после нервных окончаний. травма, повреждение.
Выводы
Опубликованные исследования нейробиологии повреждения и регенерации нервов убедительно свидетельствуют о том, что хроническая денервация шванновских клеток, хроническая аксотомия нейронов и неправильное направление регенерирующих аксонов в неправильные эндоневральные трубки в первую очередь ответственны за плохое функциональное восстановление. Эффект атрофии мышечной денервации вторичен. Экспериментальные терапевтические стратегии (которые мы в настоящее время исследуем в нашей лаборатории в Охснере) для борьбы с этими 3 нейробиологическими явлениями могут улучшить восстановление функции у пациентов, перенесших нервные повреждения.
Ключевые слова: Аксотомия , цитокины , денервация , факторы роста нервов , регенерация нервов , нейробиология , Повреждения периферических нервов , восстановление функций
ВВЕДЕНИЕ
Повреждения нервов запускают сложные клеточно-молекулярные взаимодействия, которые необходимы для регенерации аксонов и, как следствие, для значимого функционального восстановления пациентов.Эти клеточные и молекулярные ответы ненейрональных клеток периферической нервной системы (ПНС), а именно шванновских клеток (СК), создают благоприятную для роста среду для поврежденных аксонов, в отличие от реакции центральной нервной системы (ЦНС) на повреждения. Однако существует дихотомия между способностью поврежденных нейронов ПНС регенерировать свои аксоны после повреждения и субоптимальным восстановлением функции после повреждения нервов.
Значительные успехи были достигнуты в технике микрохирургии поврежденных нервов, что иногда приводит к улучшению результатов для пациентов. 1-3 К сожалению, функциональное восстановление после травм периферических нервов часто остается слабым, особенно когда повреждены нервные стволы, расположенные близко к спинному мозгу и далеко от органов-мишеней. Эти травмы включают большинство травм отрывного типа, разрывов нервов и ушибов нервов. После этих типов нервных повреждений поврежденные нейроны должны регенерировать свои аксоны на больших расстояниях с очень медленной скоростью 1 мм / день. При такой медленной скорости регенерации восстановление функциональной двигательной единицы или реиннервация органа чувств может занять месяцы или даже годы ( 1-4 ), состояние, называемое хронической аксотомией. 5-7 Аналогичным образом, SC в дистальных обрубках нерва и в органах-мишенях остаются денервированными в течение длительных периодов, состояний, известных как хроническая SC денервация 2,8-11 и хроническая денервация мышц 11 соответственно.
Схематическое изображение повреждений проксимального нерва плечевого сплетения, которые требуют длительного времени регенерации регенерирующих аксонов для реиннервации мышц руки. В это время поврежденные нейроны подвергаются хронической аксотомии, а шванновские клетки дистальной культи поврежденных нервов хронически денервированы.Графики на вставках демонстрируют прогрессирующее снижение способности поврежденных мотонейронов, которые регенерируют аксоны (число MN) и реиннервируют мышцы (число двигательных единиц [MU]), в результате хронической денервации шванновских клеток и хронической аксотомии нейронов.
Нарушение функционального восстановления кисти, например, обычно объясняется необратимой атрофией денервированных мышц и органов чувств и замещением жировой ткани. 1 Однако наши эксперименты за последние ~ 15 лет предоставляют убедительные доказательства того, что прогрессирующая неспособность хронически аксотомированных нейронов регенерировать свои аксоны и прогрессирующая неспособность хронически денервированных SCs поддерживать регенерирующие аксоны играют преобладающую роль в клинических неудачах функциональное восстановление после микрохирургического восстановления. 2,5,7,8,10-18 Плохое функциональное восстановление также усугубляется задержкой микрохирургического восстановления поврежденных нервов, когда пациенты наблюдаются для спонтанного функционального восстановления, обычно в течение 3-6 месяцев после травм тупым нервом. Регенерация аксонов в неправильные эндоневральные трубки и конечные органы — так называемое неправильное направление регенерирующих аксонов — является еще одним фактором, который способствует плохому функциональному восстановлению.
РЕГЕНЕРАЦИОННЫЙ РЕАКЦИЯ ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И ЕГО РОЛЬ В ФУНКЦИОНАЛЬНОМ РЕЗУЛЬТАТЕ
Начальная фаза регенерации аксонов
После повреждения нерва проксимальная и дистальная культя поврежденного нерва претерпевают структурные и молекулярные изменения, готовясь к процессу аксональной регенерации.Проксимальная культя претерпевает обратную дегенерацию до первого узла Ранвье, а затем каждый поврежденный аксон вырабатывает несколько дочерних аксонов. 19,20 Многие дочерние аксоны обрезаются; те, что остались, начинают процесс удлинения через дистальные культи нерва и составляют регенерирующие единицы. 21 Эта начальная стадия регенерации аксонов поддерживается как доступностью локально продуцируемых цитоскелетных материалов, так и продуцируемыми нейронами и транспортируемыми антероградно цитоскелетными белками, такими как актин и тубулин (A). 6,20,22,23 Регенерация аксонов протекает со скоростью 1-3 мм / сут, что соответствует медленной скорости транспорта цитоскелетных материалов. Дальнейшее удлинение и регенерация через дистальную культю нерва зависят от среды, поддерживающей рост, обеспечиваемой SC дистальных культей нерва. Отсутствие эндоневриальных каналов, нагруженных шванновскими клетками (полосы Бюнгнера), приводит к неправильной регенерации и образованию невром. 3,24-26
После повреждения нерва гены, связанные с регенерацией (RAG), временно активируются в нейронах, в то время как гены, связанные с нормальной синаптической передачей, подавляются (A и B).Шванновские клетки денервированных обрубков нерва подвергаются пролиферации во время дегенерации Валлера и экспрессируют множество RAG, в то время как миелин-ассоциированные гены подавляются (A). Профили генов поддерживают рост аксонов, но экспрессия очень недолговечна, так что со временем (в то время как регенерирующие аксоны растут с медленной скоростью 1 мм / сут) экспрессия RAG снижается, и способность поврежденных нейронов регенерировать свои аксонов и шванновских клеток, поддерживающих регенерацию, уменьшается (C).Примеры прогрессивного снижения уровней мРНК нанесены на графики для тубулина в нейронах и GDNF в шванновских клетках.
Роль шванновских клеток в регенерации аксонов
Дистальные культи перерезанных нервов подвергаются дегенеративному процессу, названному в честь Августа Валлера, валлеровской дегенерации. 27 Теперь мы понимаем, что валлеровская дегенерация является важным подготовительным этапом процесса регенерации аксонов, посредством которого удаляются молекулы, которые могут препятствовать регенерации.В недавнем обзоре события разбиты на три морфологически различимые стадии: острая дегенерация аксонов, латентный период и стадия резкой гранулярной дегенерации. 28 В течение часа после травмы отмирание как в проксимальной, так и в дистальной культях опосредуется каналом-опосредованным притоком кальция и активацией кальпаинов, которые расщепляют нейрофиламенты и связанные с микротрубочками компоненты, такие как спектрин и тубулин. В проксимальной культи эта перемычка относится к первому узлу Ранвье. Следующим этапом примерно в 24 часа, в течение которого продолжаются измерения аксонных потенциалов, 29 следует более длительный период разборки белков цитоскелета кальцием-активированными кальпаинами и убиквитин-протеасомной системой. 30 СК играют главную роль в этом процессе посредством фагоцитоза аксонов и миелиновых остатков. Они также секретируют хемоаттрактивные факторы, такие как интерлейкин-1 и хемоаттрактантный белок-1 моноцитов, которые рекрутируют макрофаги в денервированные дистальные культи нервов, которые вносят значительный вклад в фагоцитоз аксонов и миелиновых остатков. 23,31,32 Обломки аксонов высвобождают митогены, которые способствуют митотическому делению SC и инициируют сеть цитокинов и факторов транскрипции, которые стимулируют распад миелина и инвазию макрофагов и нацелены на миелин для фагоцитоза SC и макрофагами. 6,33
Преобразование из режима действия-потенциала-проводимости в режим, поддерживающий регенерацию: роль генов, связанных с регенерацией
Сразу после повреждения нерва потеря аксонального контакта запускает пролиферацию СК и их фенотип от миелинизирующего к немиелинизирующему фенотипу, поддерживающему рост. 6,23,34 Экспрессия матричной РНК (мРНК) миелин-ассоциированных белков, таких как P0 и миелин-ассоциированный гликопротеин, подавляется.Нейротрофины (например, фактор роста нервов, нейротрофический фактор головного мозга и нейротрофический фактор глиального происхождения), их рецепторы (например, p75, GFRA-1, GFRA-2) и молекулы адгезии (например, молекула адгезии нервных клеток) активируются при подготовке к процессу регенерации аксонов (). 6,26,35,36 Эти гены с усиленной регуляцией в совокупности называются генами, связанными с регенерацией (RAG). Изменение экспрессии генов, дегенерация и клиренс миелина и аксонов являются ключевыми особенностями процесса валлеровской дегенерации. 6,23,34
Точно так же нейроны, чьи нервы были повреждены, подавляют мРНК белков, необходимых для нейротрансмиссии, и активируют их для восстановления своих периферических процессов. 6,12,37 Следовательно, актин, тубулин и GAP-43 активируются сразу после повреждения (A и B). 6,37 Однако активация RAG не поддерживается ни в поврежденных нейронах, ни в SC. К 6 месяцам у экспериментальных животных подавляющая регуляция большинства мРНК с активизацией снижается, что приводит к потере среды, поддерживающей рост, для регенерации аксонов (A и B). 6,37 Значение ограниченной по времени активации RAG продемонстрировано в прогрессирующем снижении способности поврежденных нейронов регенерировать свои аксоны и SCs поддерживать регенерирующие аксоны по мере увеличения продолжительности восстановления нерва ().
Регенеративная способность снижается со временем из-за продолжительной аксотомии и денервации шванновских клеток. На крысах мы экспериментально увеличили либо (А) продолжительность времени, в течение которого мотонейроны не могли регенерировать свои аксоны (хроническая аксотомия; путем отсрочки наложения швов от культи проксимального нерва к недавно денервированной культе дистального нерва), либо (В) денервации. клеток Шванна в дистальных обрубках нерва (хроническая денервация; путем отсрочки наложения швов свежеразрезанной проксимальной культи нерва на культю нерва, которая была хронически денервирована до восстановления нерва).Способность нейронов к регенерации после хронической аксотомии мотонейронов или хронической денервации шванновских клеток и мышц-мишеней определялась либо путем (а) расчета количества реиннервируемых мышечных единиц (нервов и мышечных волокон, которые снабжает один мотонейрон) с использованием силы измерения в ответ на стимуляцию одиночных аксонов и мышечного нерва или (б) подсчет мечения мотонейронов, которые успешно регенерировали свои аксоны, путем нанесения ретроградного красителя на регенерирующие аксоны в культе нерва дистальнее места восстановления нерва.Оценки регенеративного успеха, полученные с помощью методов подсчета мотонейронов и подсчета двигательных единиц, хорошо согласуются, демонстрируя прогрессирующее снижение регенеративной способности как функцию (C) хронической аксотомии мотонейронов и (D) хронической денервации шванновских клеток. .
ЭКСПЕРИМЕНТАЛЬНЫЕ ПАРАДИГМЫ И ОЦЕНКА РЕГЕНЕРАЦИИ АКСОНА ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И МИКРОХИРУРГИЧЕСКОГО РЕМОНТА
Экспериментальные исследования, проведенные во время Второй мировой войны Гутманном и Янгом, показали, что замедленное восстановление нерва не влияет на скорость роста и количество аксонов. 38,39 Эти данные, свидетельствующие о крайней атрофии денервированных мышц, привели к ошибочному выводу, что плохое функциональное восстановление после повреждения нерва было связано с необратимой денервационной атрофией мышцы и ее неспособностью воспринимать иннервацию, особенно после длительных периодов воздействия. время. 38,39 Этот вывод стал довольно популярным и, к сожалению, часто повторяется даже в последних публикациях. Недавние данные нашей лаборатории и других источников не подтвердили ошибочный вывод, главным образом потому, что (1) авторы не оценили напрямую количество поврежденных нейронов, которые регенерировали в дистальную культи нерва, и тех, которые реиннервируют денервированные мышцы, и (2) подсчитали нервные волокна в дистальной культе. Культя нерва переоценивает количество регенерированных нервных волокон, потому что многие дочерние аксоны возникают из единичных регенерирующих нервных волокон в проксимальной культе нерва и могут ошибочно считаться репрезентативными для отдельных нервных волокон в проксимальной культи.
В нескольких экспериментах мы изучали процесс регенерации аксонов после немедленного и отсроченного восстановления повреждения нерва. Кроме того, мы использовали количественные методы подсчета мотонейронов, которые регенерировали свои аксоны в дистальные обрубки нервов, и подсчета количества реиннервируемых моторных единиц в целевых мышцах, чтобы оценить способность мотонейронов регенерировать свои аксоны и реиннервировать мышцы. Используя метод перекрестного шва на крысах, который позволяет независимо изучить эффекты отсроченной реиннервации дистальной культи нерва (так называемая хроническая или длительная денервация) и отсроченной регенерации нейронов до их мишеней (так называемая хроническая или длительная аксотомия), мы оценили количество регенерированных мотонейронов с использованием техники обратной мечения, при которой ретроградно транспортируемые флуоресцентные красители наносятся на срезанные концы реиннервируемых дистальных культей нерва, которые маркируют мотонейроны, регенерировавшие свои аксоны.Таким образом, мы смогли подсчитать количество мотонейронов, которые регенерировали свои аксоны в вентральном роге спинного мозга после наших окончательных экспериментов. Мы оценили количество двигательных единиц, применяя сверхмаксимальную стимуляцию к отдельным вентральным корешкам и измеряя общую мышечную силу. Затем мы выделили волокна вентрального корня и стимулировали их индивидуально, чтобы определить силу двигательных единиц. Затем количество реиннервируемых двигательных единиц рассчитывали путем деления мышечной силы на силу двигательных единиц.иллюстрирует нашу хирургическую парадигму и методы оценки регенерации аксонов и реиннервации мышц после повреждения и восстановления нерва.
Хроническая денервация шванновских клеток
Предоставление SCs среды, поддерживающей рост, тесно связано с потерей и своевременным восстановлением контакта аксонов с клетками. 10 SC, реиннервация которых задерживается регенерирующими аксонами, считаются хронически денервированными, и пагубное влияние хронической денервации на SC ухудшается по мере увеличения продолжительности хронической денервации. 2,10,18 В наших экспериментах, чтобы проверить эффект хронической денервации, общая малоберцовая (CP) ветвь седалищного нерва перерезается, и ее реиннервация откладывается на период от 1 до 6 месяцев до наложения поперечного шва. проксимальная культя свежеперерезанного большеберцового (TIB) нерва в хронически денервированный дистальный CP-нерв. Таким образом, можно непосредственно оценить влияние хронической денервации ЦП на регенерацию свежеповрежденных мотонейронов TIB. Используя эту парадигму, хроническая денервация уменьшила количество мотонейронов, которые были обратно помечены фторорубовым красителем, который был нанесен на дистальную культи нерва в 10 мм от места перекрестного шва, до менее чем 10% от количества, которое регенерировалось после немедленного ушивания культи нервов. ().Было превосходное соответствие между этой долей мотонейронов, которые регенерировали свои аксоны в хронически денервированную культю нерва, и долей недавно аксотомированных мотонейронов, которые регенерировали и реиннервировали денервированную мышцу через 4-6 месяцев (). 10 Уменьшение количества мотонейронов, которые регенерируют свои аксоны через хронически денервированные SC, не только реиннервирует денервированные мышечные волокна, но и реиннервирует в 3 раза больше мышечных волокон, чем обычно.Поскольку это увеличение реиннервируемых двигательных единиц составляет максимальную способность к отрастанию мотонейронов, 2,10 , отсюда следует, что неспособность хронически денервированных мышечных волокон принять реиннервацию ограничивает функциональное восстановление после хронических нервных повреждений. 2,10,23 Устойчивая способность денервированной мышцы принимать реиннервацию также была продемонстрирована параллельным восстановлением как мышечной массы, так и изометрической силы. Тем не менее, изоляция эффектов денервации SC от денервации мышц показала, что хронически денервированные мышечные волокна не полностью восстановили свой прежний размер, утверждая, что ограниченное количество сателлитных клеток включается в каждое атрофическое мышечное волокно для восстановления площади поперечного сечения мышечных волокон. 11 Следовательно, прогрессирующее ухудшение способности SCs поддерживать рост в дистальных культях нерва играет главную роль в плохом функциональном восстановлении после повреждения нерва, а роль мышечной атрофии вторична. Поразительно, что долгоживущие хронически денервированные SC сохраняли свою способность ремиелинизировать меньшее количество регенерированных аксонов, 2 , особенно с учетом прогрессирующей регрессии способности денервированных SC поддерживать свой разрешающий рост фенотип и прогрессирующее снижение. в количестве поддерживающих рост SC в хронически денервированных культях дистального нерва. 36,40
Клиническим коррелятом нашей экспериментальной парадигмы перекрестного шва являются повреждения больших культи нерва, при которых СК дистальных культей нерва остаются хронически денервированными из-за медленной скорости регенерации аксонов. Чем больше времени требуется регенерирующим аксонам для реиннервации СК в дистальных культях нерва, тем дольше длится период хронической денервации, в течение которого СК не контактируют с аксонами, и тем выше вероятность того, что их способность поддерживать регенерирующие аксоны нарушена. 2,6,10 Мы считаем, что отчасти поэтому реиннервация и последующее функциональное восстановление в мышечных мишенях, расположенных рядом с регенерирующими мотонейронами, лучше, чем в более дистально расположенных мышечных мишенях.
Хроническая аксотомия нейронов
Поврежденные нейроны, аксоны которых могут или не могут регенерироваться, но не установили целевые соединения, подвергаются хронической аксотомии. 5,12,14 Например, после повреждения плечевого сплетения поврежденные нейроны должны регенерироваться на большом расстоянии, прежде чем они смогут реиннервировать некоторые денервированные мышцы и, таким образом, подвергнуться хронической аксотомии.Чтобы проверить эффект хронической аксотомии, мы также использовали технику перекрестного шва с использованием 2 ветвей седалищного нерва крысы. Ветвь большеберцовой кости седалищного нерва перерезается, и ее регенерация задерживается на периоды от 1 до 6 месяцев до наложения поперечного шва проксимальной культи хронически аксотомированной проксимальной культи большеберцового нерва в только что разрезанный дистальный центральный нерв. Таким образом, можно напрямую оценить влияние хронической аксотомии TIB на реиннервацию только что поврежденного дистального CP-нерва. Используя эту парадигму и показатели результатов, описанные выше, мы обнаружили, что количество мотонейронов, регенерировавших свои аксоны, постепенно снижалось в зависимости от продолжительного времени хронической аксотомии до ~ 37% от тех, которые регенерировали без эффекта хронической аксотомии. 5,7 Это снижение способности к регенерации после хронической аксотомии является значительным, особенно в сочетании с пагубным эффектом хронической денервации, которые возникают одновременно, особенно после травм крупных нервных стволов, таких как плечевое сплетение. Прогрессирующее снижение регенеративной способности хронически аксотомированных мотонейронов сопровождалось снижением активированных RAG. 41 Драматический негативный эффект снижения регенерационной способности хронически аксотомированных мотонейронов не повлиял на способность нейронов реиннервировать в 3 раза больше мышечных волокон, чем обычно, меньшее количество мотонейронов увеличивало свои двигательные единицы и количество мышечных волокон. снабжается каждым реиннервирующим мотонейроном. 5 Таким образом, хроническая аксотомия нарушает регенеративную способность мотонейронов, но не снижает способность меньшего числа регенерированных моторных аксонов устанавливать функциональные связи с денервированными мышечными волокнами.
Шахматная регенерация аксонов и неправильное направление регенерирующих аксонов
Одним из ключевых условий успешного функционального восстановления является то, что регенерирующие аксоны регенерируют в правильные эндоневральные трубки, которые направляют их обратно к исходным органам-мишеням.Однако мы обнаружили, что регенерирующие аксоны сталкиваются со значительной задержкой в месте повреждения и проходят через место повреждения в дистальные культи нерва в шахматном порядке (шахматная регенерация аксонов). Действительно, пересечение регенерирующих аксонов происходит медленно до входа регенерирующих аксонов в дистальные обрубки нерва. Только после того, как аксоны попадают в дистальную культю, они регенерируют с медленной скоростью транспорта 1-3 мм / сут.
Функциональные результаты после травм аксонов являются наилучшими после травм с раздавливанием нерва, при которых эндоневрий остается анатомически неповрежденным на всем пути к цели, а отростки аксонов регенерирующего блока содержатся в исходном эндоневрии, поэтому регенерирующие аксоны возвращаются в исходное положение. цели. 3 Со временем реиннервируемые мышечные волокна демонстрируют нормальное мозаичное распределение типов мышечных волокон, двигательные единицы и изометрические силы мышц полностью восстанавливаются, первоначальное количество функциональных двигательных единиц восстанавливается, а количество и диаметры миелинизированных нервных волокон возвращаются к прежним значениям. обычный. 42
Повреждения нервов, приводящие к повреждению эндоневральных трубок (например, аксонотметические или нейротметические повреждения), в результате которых регенерирующие аксоны должны найти свои исходные эндоневральные трубки, представляют собой потенциальную возможность ошибок в процессе регенерации.Отростки аксонов, исходящие из культи проксимального нерва, могут попадать в несколько различных эндоневральных трубок с целевыми точками, которые они раньше не обеспечивали, как в случае моторных аксонов, которые регенерируют в путях, ведущих к коже, а не к мышцам. 43 Эксперименты, в которых ретроградные красители применялись к регенерированным аксонам перерезанных седалищных и лицевых нервов у крыс, напрямую продемонстрировали неправильное направление регенерирующих моторных аксонов от нескольких различных пулов мотонейронов к мышцам, которые мотонейроны ранее не снабжали. 44 В случаях, когда нервы снабжали мышцы антагонистическими действиями, неправильное направление аксонов было связано с несоответствующими движениями, что вполне согласуется с данными о том, что мотонейроны сгибателей и разгибателей поддерживают свой нормальный, но несоответствующий паттерн возбуждения, когда они направлены на ненадлежащую реиннервацию мышц-разгибателей и сгибателей. , соответственно. 45 Мы считаем, что это неправильное направление регенерирующих поврежденных аксонов играет важную роль в снижении функционального восстановления после травм нервов.
Есть некоторые исключения из неправильного направления регенерирующих аксонов даже после травмы транзакции. К ним относятся несколько периферических нервных стволов, ветви которых иннервируют скелетные мышцы и органы чувств в коже и суставах. Примеры включают бедренный нерв с двумя ветвями, мышечную ветвь, содержащую двигательные и сенсорные нервы к четырехглавой мышце, и другую чисто сенсорную ветвь подкожного нерва к коже. 43 Хотя реиннервация моторных и сенсорных нервных ветвей изначально случайна, мотонейроны, которые постепенно регенерируют свои аксоны через место шва, посылают свои аксоны в соответствующую моторную ветвь. 43,46 Эта преимущественная реиннервация моторики возникла параллельно с прогрессирующей или ступенчатой регенерацией моторных аксонов в дистальных обрубках нерва. 46 Необычный кислый гликан, связанный с миелиновыми профилями моторных, но не сенсорных аксонов мыши, который распознается моноклональным антителом L2 / HNK-1, может быть одним из механизмов предпочтительной реиннервации соответствующих путей. 47,48 Дифференциальный профиль нейротрофических факторов в SCs, происходящих из сенсорных и моторных аксонов с профилем BDNF / NT4 / 5, также, вероятно, играет важную роль в предпочтительной реиннервации моторных путей. 49
ВЫВОДЫ
Огромный прогресс был достигнут в нашем понимании микроанатомии, патофизиологии и микрохирургического лечения поврежденных нервов. Эти достижения улучшили качество ухода за пациентами с повреждениями нервов и часто приводят к лучшему функциональному восстановлению. Однако пациенты, у которых не удается восстановить хорошую функцию, несмотря на превосходный микрохирургический уход, представляют собой проблему для нервного хирурга. Повышенная регуляция RAG недолговечна, и, по-видимому, существует временное окно возможностей, в течение которого SCs обеспечивают среду, поддерживающую рост, и поврежденные нейроны могут регенерировать свои аксоны.Ограниченная по времени повышающая регуляция RAG и медленная скорость регенерации аксонов приводят к прогрессирующей потере нейротрофической поддержки поврежденных нейронов и их регенерирующих аксонов, отсюда хроническая денервация SC и хроническая аксотомия нейронов. 5,6,23 Поэтому в нашей лаборатории в Ochsner мы фокусируемся на 2 возможных подходах к борьбе с разрывом между временем активации RAG и медленной скоростью регенерации, включая экспериментальные стратегии, которые ускоряют скорость регенерации аксонов и поддерживают нейротрофическая среда для регенерации аксонов в течение более длительных периодов времени.Мы продемонстрировали положительный эффект кратковременной (1 ч) низкочастотной электростимуляции на ускорение регенерации аксонов за счет модуляции экспрессии RAG. 46,50-52 Мы также показали положительный эффект трансформации фактора роста бета в реактивации и поддержании экспрессии RAGS хронически денервированными SC. В настоящее время мы изучаем роль трансплантации стволовых клеток в дистальные обрубки нерва и использование усиленных нервных направляющих каналов или проводников, которые не только устранят необходимость в аутотрансплантатах, но также позволят позитивно модулировать среду, способствующую росту аксонов. прохождение направляющих каналов в дистальные культи нерва. 53,54
Мы считаем, что окончательное решение для улучшения функционального восстановления в подгруппе пациентов, которые в настоящее время не справляются с микрохирургическим вмешательством, представляет собой комбинацию нескольких нейробиологических подходов для преодоления проблемы ограниченного временного окна для оптимального восстановления нерва. регенерация.
Сноски
Авторы не имеют финансовой или имущественной заинтересованности в предмете данной статьи .
Эта статья соответствует требованиям Совета по аккредитации для последипломного медицинского образования и Американского совета по медицинским специальностям, поддерживающим сертификационные компетенции по уходу за пациентами и медицинским знаниям.
СПИСОК ЛИТЕРАТУРЫ
1. Сулейман О.А.Р., Мидха Р., Гордон Т. Патофизиология хирургических нервных расстройств. В: Winn HR, редактор. Юманс Неврологическая хирургия . 6-е изд. Филадельфия, Пенсильвания: Сондерс ;; 2011. С. 2368–2379. В. изд. [Google Scholar] 2. Сулейман О.А., Гордон Т. Влияние краткосрочной и долгосрочной денервации шванновских клеток на регенерацию периферических нервов, миелинизацию и размер. Глия . 2000 декабрь; 32 (3): 234–246. [PubMed] [Google Scholar] 3. Сандерленд С. Нервы и травмы нервов .Эдинбург, Шотландия: Черчилль Ливингстон ;; 1978. [Google Scholar] 4. Сулейман В.А., Клайн Д.Г. Хирургия нервов: обзор и взгляд на будущее. Clin Neurosurg . 2006; 53: 38–47. [PubMed] [Google Scholar] 5. Fu SY, Gordon T. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная аксотомия. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3876–3885. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Фу С.Ю., Гордон Т. Клеточные и молекулярные основы регенерации периферических нервов. Мол Нейробиол . 1997, февраль-апрель; 14 (1-2): 67–116. [PubMed] [Google Scholar] 7. Бойд Дж. Г., Гордон Т. Дозозависимое облегчение и ингибирование регенерации периферических нервов нейротрофическим фактором мозга. евро J Neurosci . 2002 Февраль; 15 (4): 613–626. [PubMed] [Google Scholar] 8. Сулейман О.А., Мидха Р., Манро К.А. и др. Хроническая денервация шванновских клеток и наличие сенсорного нерва снижают регенерацию моторных аксонов. Опыт Нейрол . 2002 август; 176 (2): 342–354. [PubMed] [Google Scholar] 9.Sulaiman OA, Voda J, Gold BG, Gordon T. FK506 увеличивает регенерацию периферических нервов после хронической аксотомии, но не после хронической денервации шванновских клеток. Опыт Нейрол . 2002 Май; 175 (1): 127–137. [PubMed] [Google Scholar] 10. Фу С.Ю., Гордон Т. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная денервация. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3886–3895. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Гордон Т., Тайреман Н., Раджи М.А. Основа для снижения функционального восстановления после отсроченного восстановления периферических нервов. Дж. Neurosci . 2011 6 апреля; 31 (14): 5325–5334. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Бойд Дж. Г., Гордон Т. Нейротрофические факторы и их рецепторы в регенерации аксонов и функциональном восстановлении после повреждения периферических нервов. Мол Нейробиол . 2003 июн; 27 (3): 277–324. [PubMed] [Google Scholar] 13. Сулейман О.А., Гордон Т. Трансформирующий фактор роста-бета и форсколин ослабляют неблагоприятные эффекты долгосрочной денервации шванновских клеток на регенерацию периферических нервов in vivo. Глия . 2002, 1 марта; 37 (3): 206–218. [PubMed] [Google Scholar] 14. Гордон Т., Фу С.Ю. Долгосрочная реакция на повреждение нерва. Adv Neurol . 1997. 72: 185–199. [PubMed] [Google Scholar] 15. Boyd JG, Gordon T. Нейротрофический фактор, полученный из линии глиальных клеток, и нейротрофический фактор, полученный из мозга, поддерживают регенерацию аксонов хронически аксотомированных мотонейронов in vivo. Опыт Нейрол . Октябрь 2003 г., 183 (2): 610–619. [PubMed] [Google Scholar] 16. Sulaiman OAR, Gordon T. TGF-бета обращает пагубный эффект длительной денервации шванновских клеток на регенерацию нервов, индуцируя экспрессию рецептора erbB3. Глия . 2003 сентябрь; 43 (Дополнение 2): 24. [Google Scholar] 17. Midha R, Munro CA, Chan S, Nitising A, Xu QG, Gordon T. Регенерация в защищенные и хронически денервированные культи периферических нервов. Нейрохирургия . 2005 декабрь; 57 (6): 1289–1299. [PubMed] [Google Scholar] 18. Сулейман О.А., Гордон Т. Роль хронической денервации шванновских клеток в плохом функциональном восстановлении после нервных повреждений и экспериментальные стратегии борьбы с ней. Нейрохирургия . Октябрь 2009 г., 65 (4 доп.): A105 – A114. [PubMed] [Google Scholar] 19.Cajal SR. Дегенерация и регенерация нервной системы . Оксфорд, Великобритания: Издательство Оксфордского университета; 1928. Перевод Р. М. Мэй. [Google Scholar] 20. Эртюрк А., Хеллал Ф., Энес Дж., Брадке Ф. Дезорганизованные микротрубочки лежат в основе образования ретракционных луковиц и нарушения регенерации аксонов. Дж. Neurosci . 22 августа 2007 г.; 27 (34): 9169–9180. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Моррис Дж. Х., Хадсон А. Р., Уэдделл Г. Исследование дегенерации и регенерации разделенного седалищного нерва крысы на основе электронной микроскопии.II. Развитие «регенерационного агрегата. Z Zellforsch Mikrosk Anat . 1972: 124 (1): 103–130. [PubMed] [Google Scholar] 22. McQuarrie IG. Влияние кондиционирующего поражения на формирование аксонального отростка в узлах Ранвье. Дж. Комп. Neurol . 1985, 8 января; 231 (2): 239–249. [PubMed] [Google Scholar] 23. Сулейман О.А., Бойд Дж. Г., Гордон Т. Регенерация аксонов в периферической системе млекопитающих. В: Kettenmann H, Ransom BR, editors. Нейроглия . 2-е изд. Оксфорд, Великобритания: Издательство Оксфордского университета; 2005 г.С. 454–466. В. ред. [Google Scholar] 24. Эйткен Дж. Т., Шарман М., Янг Дж. З. Созревание периферических нервных волокон с различными периферическими связями. Дж Анат . 1947; 81: 1–22. [PubMed] [Google Scholar] 25. Toft PB, Fugleholm K, Schmalbruch H. Разветвление аксонов после раздавливания периферических нервов крысы. Мышечный нерв . 1988 августа; 11 (8): 880–889. [PubMed] [Google Scholar] 26. Наварро X, Виво М., Валеро-Кабре А. Нейропластичность после повреждения и регенерации периферических нервов. Прог Нейробиол . Июль 2007 г .; 82 (4): 163–201. Epub 2007 22 июня. [PubMed] [Google Scholar] 27. Уоллер А. Эксперименты на разрезе языкоглоточного и подъязычного нервов лягушки и наблюдения за изменениями, вызванными этим в структуре их примитивных волокон. Philos Trans R Soc London . 1850; 140: 423–429. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Ван Дж.Т., Медресс З.А., Баррес Б.А. Дегенерация аксонов: молекулярные механизмы пути самоуничтожения. Дж. Ячейка Биол . 2012 9 января; 196 (1): 7–18. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Чжай К., Ван Дж., Ким А. и др. Участие убиквитин-протеасомной системы на ранних стадиях валлеровской дегенерации. Нейрон . 17 июля 2003 г.; 39 (2): 217–225. [PubMed] [Google Scholar] 31. Сулейман О.А., Клеточный Гордон Т. и молекулярные взаимодействия после повреждения периферических и центральных нервов. Биомед Ред. . 2003. 14: 51–62. [Google Scholar] 32. Хёке А., Ми Р. В поисках новых методов лечения периферических невропатий и регенерации нервов. Дисков Мед . 2007 август; 7 (39): 109–112. [PubMed] [Google Scholar] 33. Brushart TM. Ремонт нервов . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета ;; 2011. [Google Scholar] 34. Scherer SS, Salzer JL. Аксон-шванновские клеточные взаимодействия во время дегенерации и регенерации периферических нервов. В: Jessen KR, Richardson WD, редакторы. Развитие глиальных клеток: основные принципы и клиническая значимость . Оксфорд, Великобритания: Bios Scientific ;; 1996. С. 169–196. В. ред. [Google Scholar] 35. Хёке А., Гордон Т., Зочодне Д.В., Сулейман О.А.Снижение экспрессии нейротрофического фактора, происходящего из глиальных клеток, связано с нарушением регенерации после длительной денервации шванновских клеток. Опыт Нейрол . 2002 Янв; 173 (1): 77–85. [PubMed] [Google Scholar] 36. You S, Petrov T, Chung PH, Gordon T. Экспрессия рецептора фактора роста нервов с низким сродством в долгосрочно денервированных шванновских клетках. Глия . 1997 июн; 20 (2): 87–100. [PubMed] [Google Scholar] 37. Бисби М.А., Тецлафф В. Изменения в синтезе цитоскелетного белка после повреждения аксона и во время регенерации аксона. Мол Нейробиол . 1992; 6 (2-3): 107–123. Лето осень. [PubMed] [Google Scholar] 38. Гутманн Э. Влияние задержки иннервации на восстановление мышц после нервных поражений. Дж. Нейрофизиол . 1948 июл; 11 (4): 279–294. [PubMed] [Google Scholar] 40. Ли Х, Теренги Г, зал СМ. Влияние отсроченной реиннервации на экспрессию рецепторов c-erbB хронически денервированными шванновскими клетками крысы in vivo. Глия . 1997 августа; 20 (4): 333–347. [PubMed] [Google Scholar] 41. Тецлафф В., Леонард С., Крекоски К.А., Пархад И.М., Бисби Массачусетс.Снижение синтеза мотонейрональных нейрофиламентов путем последовательных аксотомий: возможное объяснение эффекта кондиционирующего поражения на регенерацию аксонов. Опыт Нейрол . 1996 Май; 139 (1): 95–106. [PubMed] [Google Scholar] 42. Гордон Т., Паттулло М.С. Пластичность мышечных волокон и типов двигательных единиц. Exerc Sport Sci Ред. . 1993; 21: 331–362. [PubMed] [Google Scholar] 44. Brushart TM, Mesulam MM. Трансганглионарная демонстрация центральных сенсорных проекций кожи и мышц с конъюгатами HRP-лектин. Neurosci Lett . 1980 апр; 17 (1-2): 1–6. [PubMed] [Google Scholar] 45. Гордон Т, Стейн РБ, Томас СК. Организация двигательных единиц после перекрестной реиннервации мышц-антагонистов в задней конечности кошки. Дж. Физиология . 1986 Май; 374: 443–456. [Бесплатная статья PMC] [PubMed] [Google Scholar] 46. Аль-Маджед А.А., Нойманн С.М., Брушарт TM, Гордон Т. Кратковременная электрическая стимуляция повышает скорость и точность регенерации моторных аксонов. Дж. Neurosci . 2000 г., 1 апреля; 20 (7): 2602–2608.[Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Мартини Р., Шахнер М., Брушарт TM. Углевод L2 / HNK-1 предпочтительно экспрессируется шванновскими клетками, ранее ассоциированными с моторными аксонами, в реиннервируемых периферических нервах. Дж. Neurosci . 1994, ноябрь; 14 (11, часть 2): 7180–7191. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Эберхардт К.А., Иринчев А.А., Аль-Маджед А.А. и др. Передача сигналов BDNF / TrkB регулирует экспрессию углеводов HNK-1 в регенерирующих двигательных нервах и способствует функциональному восстановлению после восстановления периферических нервов. Опыт Нейрол . 2006 Апрель; 198 (2): 500–510. Epub 2006 7 февраля. [PubMed] [Google Scholar] 49. Хёке А., Редетт Р., Хамид Х и др. Шванновские клетки выражают моторные и сенсорные фенотипы, которые регулируют регенерацию аксонов. Дж. Neurosci . 2006 20 сентября; 26 (38): 9646–9655. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Al-Majed AA, Brushart TM, Gordon T. Электрическая стимуляция ускоряет и увеличивает экспрессию мРНК BDNF и trkB в регенерирующих мотонейронах бедренной кости крысы. евро J Neurosci .2000 декабрь; 12 (12): 4381–4390. [PubMed] [Google Scholar] 51. Brushart TM, Hoffman PN, Royall RM и др. Электрическая стимуляция способствует регенерации мотонейрона без увеличения его скорости и без кондиционирования нейрона. Дж. Neurosci . 2002, 1 августа; 22 (15): 6631–6638. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Гордон Т., Сулейман О.А., Ладак А. Глава 24: Электростимуляция для улучшения регенерации нервов: где мы находимся? Международная версия Neurobiol . 2009. 87: 433–444. [PubMed] [Google Scholar] 53.Мидха Р. Новые методы восстановления нервов: нервные передачи и нервные трубки. Clin Neurosurg . 2006. 53: 185–190. [PubMed] [Google Scholar] 54. Пфистер Л.А., Папалоизос М., Меркл Г.П., Гандер Б. Нервные проводники и доставка факторов роста при восстановлении периферических нервов. J Peripher Nerv Syst . 2007 июн; 12 (2): 65–82. [PubMed] [Google Scholar]
От лабораторных исследований до прикроватных приложений
Охснер Дж. 2013 Весна; 13 (1): 100–108.
, MD, PhD, FRCS * и, PhD †
Wale Sulaiman
* Отделение нейрохирургии, Ochsner Clinic Foundation, и Медицинская школа Университета Квинсленда, Клиническая школа Охснера, Новый Орлеан, Луизиана
Тесса Гордон
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
* Отделение нейрохирургии, Фонд клиники Окснера и Школа медицины Университета Квинсленда, Клиническая школа Окснера, Новый Орлеан, Луизиана
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
Адресная корреспонденция Wale Sulaiman, MD, PhD, FRCS, Отделение нейрохирургии, Ochsner Clinic Foundation, 1514 Jefferson Hwy., New Orleans, LA 70121. Тел: (504) 842-2181 Факс: (504) 842-6531 Электронная почта: gro.renshco@namialusw Авторские права © Академическое подразделение Фонда клиники Окснера Эта статья цитируется в других статьях в PMC.
РЕФЕРАТ
Объективы
Мы рассматриваем современную нейробиологию повреждения и регенерации нервов, особенно в том, что касается восстановления полезной функции у пациентов, которые получили травмы крупных нервных стволов, таких как плечевое сплетение.
Методы
В этом обзоре основное внимание уделяется исследованиям, проведенным в нашей лаборатории в Окснеру и в других лабораториях, связанных с нейробиологией повреждения нервов, с акцентом на то, как некоторые из ключевых результатов исследований на животных помогают нам понять патофизиологию плохого функционального восстановления после нервных окончаний. травма, повреждение.
Выводы
Опубликованные исследования нейробиологии повреждения и регенерации нервов убедительно свидетельствуют о том, что хроническая денервация шванновских клеток, хроническая аксотомия нейронов и неправильное направление регенерирующих аксонов в неправильные эндоневральные трубки в первую очередь ответственны за плохое функциональное восстановление. Эффект атрофии мышечной денервации вторичен. Экспериментальные терапевтические стратегии (которые мы в настоящее время исследуем в нашей лаборатории в Охснере) для борьбы с этими 3 нейробиологическими явлениями могут улучшить восстановление функции у пациентов, перенесших нервные повреждения.
Ключевые слова: Аксотомия , цитокины , денервация , факторы роста нервов , регенерация нервов , нейробиология , Повреждения периферических нервов , восстановление функций
ВВЕДЕНИЕ
Повреждения нервов запускают сложные клеточно-молекулярные взаимодействия, которые необходимы для регенерации аксонов и, как следствие, для значимого функционального восстановления пациентов.Эти клеточные и молекулярные ответы ненейрональных клеток периферической нервной системы (ПНС), а именно шванновских клеток (СК), создают благоприятную для роста среду для поврежденных аксонов, в отличие от реакции центральной нервной системы (ЦНС) на повреждения. Однако существует дихотомия между способностью поврежденных нейронов ПНС регенерировать свои аксоны после повреждения и субоптимальным восстановлением функции после повреждения нервов.
Значительные успехи были достигнуты в технике микрохирургии поврежденных нервов, что иногда приводит к улучшению результатов для пациентов. 1-3 К сожалению, функциональное восстановление после травм периферических нервов часто остается слабым, особенно когда повреждены нервные стволы, расположенные близко к спинному мозгу и далеко от органов-мишеней. Эти травмы включают большинство травм отрывного типа, разрывов нервов и ушибов нервов. После этих типов нервных повреждений поврежденные нейроны должны регенерировать свои аксоны на больших расстояниях с очень медленной скоростью 1 мм / день. При такой медленной скорости регенерации восстановление функциональной двигательной единицы или реиннервация органа чувств может занять месяцы или даже годы ( 1-4 ), состояние, называемое хронической аксотомией. 5-7 Аналогичным образом, SC в дистальных обрубках нерва и в органах-мишенях остаются денервированными в течение длительных периодов, состояний, известных как хроническая SC денервация 2,8-11 и хроническая денервация мышц 11 соответственно.
Схематическое изображение повреждений проксимального нерва плечевого сплетения, которые требуют длительного времени регенерации регенерирующих аксонов для реиннервации мышц руки. В это время поврежденные нейроны подвергаются хронической аксотомии, а шванновские клетки дистальной культи поврежденных нервов хронически денервированы.Графики на вставках демонстрируют прогрессирующее снижение способности поврежденных мотонейронов, которые регенерируют аксоны (число MN) и реиннервируют мышцы (число двигательных единиц [MU]), в результате хронической денервации шванновских клеток и хронической аксотомии нейронов.
Нарушение функционального восстановления кисти, например, обычно объясняется необратимой атрофией денервированных мышц и органов чувств и замещением жировой ткани. 1 Однако наши эксперименты за последние ~ 15 лет предоставляют убедительные доказательства того, что прогрессирующая неспособность хронически аксотомированных нейронов регенерировать свои аксоны и прогрессирующая неспособность хронически денервированных SCs поддерживать регенерирующие аксоны играют преобладающую роль в клинических неудачах функциональное восстановление после микрохирургического восстановления. 2,5,7,8,10-18 Плохое функциональное восстановление также усугубляется задержкой микрохирургического восстановления поврежденных нервов, когда пациенты наблюдаются для спонтанного функционального восстановления, обычно в течение 3-6 месяцев после травм тупым нервом. Регенерация аксонов в неправильные эндоневральные трубки и конечные органы — так называемое неправильное направление регенерирующих аксонов — является еще одним фактором, который способствует плохому функциональному восстановлению.
РЕГЕНЕРАЦИОННЫЙ РЕАКЦИЯ ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И ЕГО РОЛЬ В ФУНКЦИОНАЛЬНОМ РЕЗУЛЬТАТЕ
Начальная фаза регенерации аксонов
После повреждения нерва проксимальная и дистальная культя поврежденного нерва претерпевают структурные и молекулярные изменения, готовясь к процессу аксональной регенерации.Проксимальная культя претерпевает обратную дегенерацию до первого узла Ранвье, а затем каждый поврежденный аксон вырабатывает несколько дочерних аксонов. 19,20 Многие дочерние аксоны обрезаются; те, что остались, начинают процесс удлинения через дистальные культи нерва и составляют регенерирующие единицы. 21 Эта начальная стадия регенерации аксонов поддерживается как доступностью локально продуцируемых цитоскелетных материалов, так и продуцируемыми нейронами и транспортируемыми антероградно цитоскелетными белками, такими как актин и тубулин (A). 6,20,22,23 Регенерация аксонов протекает со скоростью 1-3 мм / сут, что соответствует медленной скорости транспорта цитоскелетных материалов. Дальнейшее удлинение и регенерация через дистальную культю нерва зависят от среды, поддерживающей рост, обеспечиваемой SC дистальных культей нерва. Отсутствие эндоневриальных каналов, нагруженных шванновскими клетками (полосы Бюнгнера), приводит к неправильной регенерации и образованию невром. 3,24-26
После повреждения нерва гены, связанные с регенерацией (RAG), временно активируются в нейронах, в то время как гены, связанные с нормальной синаптической передачей, подавляются (A и B).Шванновские клетки денервированных обрубков нерва подвергаются пролиферации во время дегенерации Валлера и экспрессируют множество RAG, в то время как миелин-ассоциированные гены подавляются (A). Профили генов поддерживают рост аксонов, но экспрессия очень недолговечна, так что со временем (в то время как регенерирующие аксоны растут с медленной скоростью 1 мм / сут) экспрессия RAG снижается, и способность поврежденных нейронов регенерировать свои аксонов и шванновских клеток, поддерживающих регенерацию, уменьшается (C).Примеры прогрессивного снижения уровней мРНК нанесены на графики для тубулина в нейронах и GDNF в шванновских клетках.
Роль шванновских клеток в регенерации аксонов
Дистальные культи перерезанных нервов подвергаются дегенеративному процессу, названному в честь Августа Валлера, валлеровской дегенерации. 27 Теперь мы понимаем, что валлеровская дегенерация является важным подготовительным этапом процесса регенерации аксонов, посредством которого удаляются молекулы, которые могут препятствовать регенерации.В недавнем обзоре события разбиты на три морфологически различимые стадии: острая дегенерация аксонов, латентный период и стадия резкой гранулярной дегенерации. 28 В течение часа после травмы отмирание как в проксимальной, так и в дистальной культях опосредуется каналом-опосредованным притоком кальция и активацией кальпаинов, которые расщепляют нейрофиламенты и связанные с микротрубочками компоненты, такие как спектрин и тубулин. В проксимальной культи эта перемычка относится к первому узлу Ранвье. Следующим этапом примерно в 24 часа, в течение которого продолжаются измерения аксонных потенциалов, 29 следует более длительный период разборки белков цитоскелета кальцием-активированными кальпаинами и убиквитин-протеасомной системой. 30 СК играют главную роль в этом процессе посредством фагоцитоза аксонов и миелиновых остатков. Они также секретируют хемоаттрактивные факторы, такие как интерлейкин-1 и хемоаттрактантный белок-1 моноцитов, которые рекрутируют макрофаги в денервированные дистальные культи нервов, которые вносят значительный вклад в фагоцитоз аксонов и миелиновых остатков. 23,31,32 Обломки аксонов высвобождают митогены, которые способствуют митотическому делению SC и инициируют сеть цитокинов и факторов транскрипции, которые стимулируют распад миелина и инвазию макрофагов и нацелены на миелин для фагоцитоза SC и макрофагами. 6,33
Преобразование из режима действия-потенциала-проводимости в режим, поддерживающий регенерацию: роль генов, связанных с регенерацией
Сразу после повреждения нерва потеря аксонального контакта запускает пролиферацию СК и их фенотип от миелинизирующего к немиелинизирующему фенотипу, поддерживающему рост. 6,23,34 Экспрессия матричной РНК (мРНК) миелин-ассоциированных белков, таких как P0 и миелин-ассоциированный гликопротеин, подавляется.Нейротрофины (например, фактор роста нервов, нейротрофический фактор головного мозга и нейротрофический фактор глиального происхождения), их рецепторы (например, p75, GFRA-1, GFRA-2) и молекулы адгезии (например, молекула адгезии нервных клеток) активируются при подготовке к процессу регенерации аксонов (). 6,26,35,36 Эти гены с усиленной регуляцией в совокупности называются генами, связанными с регенерацией (RAG). Изменение экспрессии генов, дегенерация и клиренс миелина и аксонов являются ключевыми особенностями процесса валлеровской дегенерации. 6,23,34
Точно так же нейроны, чьи нервы были повреждены, подавляют мРНК белков, необходимых для нейротрансмиссии, и активируют их для восстановления своих периферических процессов. 6,12,37 Следовательно, актин, тубулин и GAP-43 активируются сразу после повреждения (A и B). 6,37 Однако активация RAG не поддерживается ни в поврежденных нейронах, ни в SC. К 6 месяцам у экспериментальных животных подавляющая регуляция большинства мРНК с активизацией снижается, что приводит к потере среды, поддерживающей рост, для регенерации аксонов (A и B). 6,37 Значение ограниченной по времени активации RAG продемонстрировано в прогрессирующем снижении способности поврежденных нейронов регенерировать свои аксоны и SCs поддерживать регенерирующие аксоны по мере увеличения продолжительности восстановления нерва ().
Регенеративная способность снижается со временем из-за продолжительной аксотомии и денервации шванновских клеток. На крысах мы экспериментально увеличили либо (А) продолжительность времени, в течение которого мотонейроны не могли регенерировать свои аксоны (хроническая аксотомия; путем отсрочки наложения швов от культи проксимального нерва к недавно денервированной культе дистального нерва), либо (В) денервации. клеток Шванна в дистальных обрубках нерва (хроническая денервация; путем отсрочки наложения швов свежеразрезанной проксимальной культи нерва на культю нерва, которая была хронически денервирована до восстановления нерва).Способность нейронов к регенерации после хронической аксотомии мотонейронов или хронической денервации шванновских клеток и мышц-мишеней определялась либо путем (а) расчета количества реиннервируемых мышечных единиц (нервов и мышечных волокон, которые снабжает один мотонейрон) с использованием силы измерения в ответ на стимуляцию одиночных аксонов и мышечного нерва или (б) подсчет мечения мотонейронов, которые успешно регенерировали свои аксоны, путем нанесения ретроградного красителя на регенерирующие аксоны в культе нерва дистальнее места восстановления нерва.Оценки регенеративного успеха, полученные с помощью методов подсчета мотонейронов и подсчета двигательных единиц, хорошо согласуются, демонстрируя прогрессирующее снижение регенеративной способности как функцию (C) хронической аксотомии мотонейронов и (D) хронической денервации шванновских клеток. .
ЭКСПЕРИМЕНТАЛЬНЫЕ ПАРАДИГМЫ И ОЦЕНКА РЕГЕНЕРАЦИИ АКСОНА ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И МИКРОХИРУРГИЧЕСКОГО РЕМОНТА
Экспериментальные исследования, проведенные во время Второй мировой войны Гутманном и Янгом, показали, что замедленное восстановление нерва не влияет на скорость роста и количество аксонов. 38,39 Эти данные, свидетельствующие о крайней атрофии денервированных мышц, привели к ошибочному выводу, что плохое функциональное восстановление после повреждения нерва было связано с необратимой денервационной атрофией мышцы и ее неспособностью воспринимать иннервацию, особенно после длительных периодов воздействия. время. 38,39 Этот вывод стал довольно популярным и, к сожалению, часто повторяется даже в последних публикациях. Недавние данные нашей лаборатории и других источников не подтвердили ошибочный вывод, главным образом потому, что (1) авторы не оценили напрямую количество поврежденных нейронов, которые регенерировали в дистальную культи нерва, и тех, которые реиннервируют денервированные мышцы, и (2) подсчитали нервные волокна в дистальной культе. Культя нерва переоценивает количество регенерированных нервных волокон, потому что многие дочерние аксоны возникают из единичных регенерирующих нервных волокон в проксимальной культе нерва и могут ошибочно считаться репрезентативными для отдельных нервных волокон в проксимальной культи.
В нескольких экспериментах мы изучали процесс регенерации аксонов после немедленного и отсроченного восстановления повреждения нерва. Кроме того, мы использовали количественные методы подсчета мотонейронов, которые регенерировали свои аксоны в дистальные обрубки нервов, и подсчета количества реиннервируемых моторных единиц в целевых мышцах, чтобы оценить способность мотонейронов регенерировать свои аксоны и реиннервировать мышцы. Используя метод перекрестного шва на крысах, который позволяет независимо изучить эффекты отсроченной реиннервации дистальной культи нерва (так называемая хроническая или длительная денервация) и отсроченной регенерации нейронов до их мишеней (так называемая хроническая или длительная аксотомия), мы оценили количество регенерированных мотонейронов с использованием техники обратной мечения, при которой ретроградно транспортируемые флуоресцентные красители наносятся на срезанные концы реиннервируемых дистальных культей нерва, которые маркируют мотонейроны, регенерировавшие свои аксоны.Таким образом, мы смогли подсчитать количество мотонейронов, которые регенерировали свои аксоны в вентральном роге спинного мозга после наших окончательных экспериментов. Мы оценили количество двигательных единиц, применяя сверхмаксимальную стимуляцию к отдельным вентральным корешкам и измеряя общую мышечную силу. Затем мы выделили волокна вентрального корня и стимулировали их индивидуально, чтобы определить силу двигательных единиц. Затем количество реиннервируемых двигательных единиц рассчитывали путем деления мышечной силы на силу двигательных единиц.иллюстрирует нашу хирургическую парадигму и методы оценки регенерации аксонов и реиннервации мышц после повреждения и восстановления нерва.
Хроническая денервация шванновских клеток
Предоставление SCs среды, поддерживающей рост, тесно связано с потерей и своевременным восстановлением контакта аксонов с клетками. 10 SC, реиннервация которых задерживается регенерирующими аксонами, считаются хронически денервированными, и пагубное влияние хронической денервации на SC ухудшается по мере увеличения продолжительности хронической денервации. 2,10,18 В наших экспериментах, чтобы проверить эффект хронической денервации, общая малоберцовая (CP) ветвь седалищного нерва перерезается, и ее реиннервация откладывается на период от 1 до 6 месяцев до наложения поперечного шва. проксимальная культя свежеперерезанного большеберцового (TIB) нерва в хронически денервированный дистальный CP-нерв. Таким образом, можно непосредственно оценить влияние хронической денервации ЦП на регенерацию свежеповрежденных мотонейронов TIB. Используя эту парадигму, хроническая денервация уменьшила количество мотонейронов, которые были обратно помечены фторорубовым красителем, который был нанесен на дистальную культи нерва в 10 мм от места перекрестного шва, до менее чем 10% от количества, которое регенерировалось после немедленного ушивания культи нервов. ().Было превосходное соответствие между этой долей мотонейронов, которые регенерировали свои аксоны в хронически денервированную культю нерва, и долей недавно аксотомированных мотонейронов, которые регенерировали и реиннервировали денервированную мышцу через 4-6 месяцев (). 10 Уменьшение количества мотонейронов, которые регенерируют свои аксоны через хронически денервированные SC, не только реиннервирует денервированные мышечные волокна, но и реиннервирует в 3 раза больше мышечных волокон, чем обычно.Поскольку это увеличение реиннервируемых двигательных единиц составляет максимальную способность к отрастанию мотонейронов, 2,10 , отсюда следует, что неспособность хронически денервированных мышечных волокон принять реиннервацию ограничивает функциональное восстановление после хронических нервных повреждений. 2,10,23 Устойчивая способность денервированной мышцы принимать реиннервацию также была продемонстрирована параллельным восстановлением как мышечной массы, так и изометрической силы. Тем не менее, изоляция эффектов денервации SC от денервации мышц показала, что хронически денервированные мышечные волокна не полностью восстановили свой прежний размер, утверждая, что ограниченное количество сателлитных клеток включается в каждое атрофическое мышечное волокно для восстановления площади поперечного сечения мышечных волокон. 11 Следовательно, прогрессирующее ухудшение способности SCs поддерживать рост в дистальных культях нерва играет главную роль в плохом функциональном восстановлении после повреждения нерва, а роль мышечной атрофии вторична. Поразительно, что долгоживущие хронически денервированные SC сохраняли свою способность ремиелинизировать меньшее количество регенерированных аксонов, 2 , особенно с учетом прогрессирующей регрессии способности денервированных SC поддерживать свой разрешающий рост фенотип и прогрессирующее снижение. в количестве поддерживающих рост SC в хронически денервированных культях дистального нерва. 36,40
Клиническим коррелятом нашей экспериментальной парадигмы перекрестного шва являются повреждения больших культи нерва, при которых СК дистальных культей нерва остаются хронически денервированными из-за медленной скорости регенерации аксонов. Чем больше времени требуется регенерирующим аксонам для реиннервации СК в дистальных культях нерва, тем дольше длится период хронической денервации, в течение которого СК не контактируют с аксонами, и тем выше вероятность того, что их способность поддерживать регенерирующие аксоны нарушена. 2,6,10 Мы считаем, что отчасти поэтому реиннервация и последующее функциональное восстановление в мышечных мишенях, расположенных рядом с регенерирующими мотонейронами, лучше, чем в более дистально расположенных мышечных мишенях.
Хроническая аксотомия нейронов
Поврежденные нейроны, аксоны которых могут или не могут регенерироваться, но не установили целевые соединения, подвергаются хронической аксотомии. 5,12,14 Например, после повреждения плечевого сплетения поврежденные нейроны должны регенерироваться на большом расстоянии, прежде чем они смогут реиннервировать некоторые денервированные мышцы и, таким образом, подвергнуться хронической аксотомии.Чтобы проверить эффект хронической аксотомии, мы также использовали технику перекрестного шва с использованием 2 ветвей седалищного нерва крысы. Ветвь большеберцовой кости седалищного нерва перерезается, и ее регенерация задерживается на периоды от 1 до 6 месяцев до наложения поперечного шва проксимальной культи хронически аксотомированной проксимальной культи большеберцового нерва в только что разрезанный дистальный центральный нерв. Таким образом, можно напрямую оценить влияние хронической аксотомии TIB на реиннервацию только что поврежденного дистального CP-нерва. Используя эту парадигму и показатели результатов, описанные выше, мы обнаружили, что количество мотонейронов, регенерировавших свои аксоны, постепенно снижалось в зависимости от продолжительного времени хронической аксотомии до ~ 37% от тех, которые регенерировали без эффекта хронической аксотомии. 5,7 Это снижение способности к регенерации после хронической аксотомии является значительным, особенно в сочетании с пагубным эффектом хронической денервации, которые возникают одновременно, особенно после травм крупных нервных стволов, таких как плечевое сплетение. Прогрессирующее снижение регенеративной способности хронически аксотомированных мотонейронов сопровождалось снижением активированных RAG. 41 Драматический негативный эффект снижения регенерационной способности хронически аксотомированных мотонейронов не повлиял на способность нейронов реиннервировать в 3 раза больше мышечных волокон, чем обычно, меньшее количество мотонейронов увеличивало свои двигательные единицы и количество мышечных волокон. снабжается каждым реиннервирующим мотонейроном. 5 Таким образом, хроническая аксотомия нарушает регенеративную способность мотонейронов, но не снижает способность меньшего числа регенерированных моторных аксонов устанавливать функциональные связи с денервированными мышечными волокнами.
Шахматная регенерация аксонов и неправильное направление регенерирующих аксонов
Одним из ключевых условий успешного функционального восстановления является то, что регенерирующие аксоны регенерируют в правильные эндоневральные трубки, которые направляют их обратно к исходным органам-мишеням.Однако мы обнаружили, что регенерирующие аксоны сталкиваются со значительной задержкой в месте повреждения и проходят через место повреждения в дистальные культи нерва в шахматном порядке (шахматная регенерация аксонов). Действительно, пересечение регенерирующих аксонов происходит медленно до входа регенерирующих аксонов в дистальные обрубки нерва. Только после того, как аксоны попадают в дистальную культю, они регенерируют с медленной скоростью транспорта 1-3 мм / сут.
Функциональные результаты после травм аксонов являются наилучшими после травм с раздавливанием нерва, при которых эндоневрий остается анатомически неповрежденным на всем пути к цели, а отростки аксонов регенерирующего блока содержатся в исходном эндоневрии, поэтому регенерирующие аксоны возвращаются в исходное положение. цели. 3 Со временем реиннервируемые мышечные волокна демонстрируют нормальное мозаичное распределение типов мышечных волокон, двигательные единицы и изометрические силы мышц полностью восстанавливаются, первоначальное количество функциональных двигательных единиц восстанавливается, а количество и диаметры миелинизированных нервных волокон возвращаются к прежним значениям. обычный. 42
Повреждения нервов, приводящие к повреждению эндоневральных трубок (например, аксонотметические или нейротметические повреждения), в результате которых регенерирующие аксоны должны найти свои исходные эндоневральные трубки, представляют собой потенциальную возможность ошибок в процессе регенерации.Отростки аксонов, исходящие из культи проксимального нерва, могут попадать в несколько различных эндоневральных трубок с целевыми точками, которые они раньше не обеспечивали, как в случае моторных аксонов, которые регенерируют в путях, ведущих к коже, а не к мышцам. 43 Эксперименты, в которых ретроградные красители применялись к регенерированным аксонам перерезанных седалищных и лицевых нервов у крыс, напрямую продемонстрировали неправильное направление регенерирующих моторных аксонов от нескольких различных пулов мотонейронов к мышцам, которые мотонейроны ранее не снабжали. 44 В случаях, когда нервы снабжали мышцы антагонистическими действиями, неправильное направление аксонов было связано с несоответствующими движениями, что вполне согласуется с данными о том, что мотонейроны сгибателей и разгибателей поддерживают свой нормальный, но несоответствующий паттерн возбуждения, когда они направлены на ненадлежащую реиннервацию мышц-разгибателей и сгибателей. , соответственно. 45 Мы считаем, что это неправильное направление регенерирующих поврежденных аксонов играет важную роль в снижении функционального восстановления после травм нервов.
Есть некоторые исключения из неправильного направления регенерирующих аксонов даже после травмы транзакции. К ним относятся несколько периферических нервных стволов, ветви которых иннервируют скелетные мышцы и органы чувств в коже и суставах. Примеры включают бедренный нерв с двумя ветвями, мышечную ветвь, содержащую двигательные и сенсорные нервы к четырехглавой мышце, и другую чисто сенсорную ветвь подкожного нерва к коже. 43 Хотя реиннервация моторных и сенсорных нервных ветвей изначально случайна, мотонейроны, которые постепенно регенерируют свои аксоны через место шва, посылают свои аксоны в соответствующую моторную ветвь. 43,46 Эта преимущественная реиннервация моторики возникла параллельно с прогрессирующей или ступенчатой регенерацией моторных аксонов в дистальных обрубках нерва. 46 Необычный кислый гликан, связанный с миелиновыми профилями моторных, но не сенсорных аксонов мыши, который распознается моноклональным антителом L2 / HNK-1, может быть одним из механизмов предпочтительной реиннервации соответствующих путей. 47,48 Дифференциальный профиль нейротрофических факторов в SCs, происходящих из сенсорных и моторных аксонов с профилем BDNF / NT4 / 5, также, вероятно, играет важную роль в предпочтительной реиннервации моторных путей. 49
ВЫВОДЫ
Огромный прогресс был достигнут в нашем понимании микроанатомии, патофизиологии и микрохирургического лечения поврежденных нервов. Эти достижения улучшили качество ухода за пациентами с повреждениями нервов и часто приводят к лучшему функциональному восстановлению. Однако пациенты, у которых не удается восстановить хорошую функцию, несмотря на превосходный микрохирургический уход, представляют собой проблему для нервного хирурга. Повышенная регуляция RAG недолговечна, и, по-видимому, существует временное окно возможностей, в течение которого SCs обеспечивают среду, поддерживающую рост, и поврежденные нейроны могут регенерировать свои аксоны.Ограниченная по времени повышающая регуляция RAG и медленная скорость регенерации аксонов приводят к прогрессирующей потере нейротрофической поддержки поврежденных нейронов и их регенерирующих аксонов, отсюда хроническая денервация SC и хроническая аксотомия нейронов. 5,6,23 Поэтому в нашей лаборатории в Ochsner мы фокусируемся на 2 возможных подходах к борьбе с разрывом между временем активации RAG и медленной скоростью регенерации, включая экспериментальные стратегии, которые ускоряют скорость регенерации аксонов и поддерживают нейротрофическая среда для регенерации аксонов в течение более длительных периодов времени.Мы продемонстрировали положительный эффект кратковременной (1 ч) низкочастотной электростимуляции на ускорение регенерации аксонов за счет модуляции экспрессии RAG. 46,50-52 Мы также показали положительный эффект трансформации фактора роста бета в реактивации и поддержании экспрессии RAGS хронически денервированными SC. В настоящее время мы изучаем роль трансплантации стволовых клеток в дистальные обрубки нерва и использование усиленных нервных направляющих каналов или проводников, которые не только устранят необходимость в аутотрансплантатах, но также позволят позитивно модулировать среду, способствующую росту аксонов. прохождение направляющих каналов в дистальные культи нерва. 53,54
Мы считаем, что окончательное решение для улучшения функционального восстановления в подгруппе пациентов, которые в настоящее время не справляются с микрохирургическим вмешательством, представляет собой комбинацию нескольких нейробиологических подходов для преодоления проблемы ограниченного временного окна для оптимального восстановления нерва. регенерация.
Сноски
Авторы не имеют финансовой или имущественной заинтересованности в предмете данной статьи .
Эта статья соответствует требованиям Совета по аккредитации для последипломного медицинского образования и Американского совета по медицинским специальностям, поддерживающим сертификационные компетенции по уходу за пациентами и медицинским знаниям.
СПИСОК ЛИТЕРАТУРЫ
1. Сулейман О.А.Р., Мидха Р., Гордон Т. Патофизиология хирургических нервных расстройств. В: Winn HR, редактор. Юманс Неврологическая хирургия . 6-е изд. Филадельфия, Пенсильвания: Сондерс ;; 2011. С. 2368–2379. В. изд. [Google Scholar] 2. Сулейман О.А., Гордон Т. Влияние краткосрочной и долгосрочной денервации шванновских клеток на регенерацию периферических нервов, миелинизацию и размер. Глия . 2000 декабрь; 32 (3): 234–246. [PubMed] [Google Scholar] 3. Сандерленд С. Нервы и травмы нервов .Эдинбург, Шотландия: Черчилль Ливингстон ;; 1978. [Google Scholar] 4. Сулейман В.А., Клайн Д.Г. Хирургия нервов: обзор и взгляд на будущее. Clin Neurosurg . 2006; 53: 38–47. [PubMed] [Google Scholar] 5. Fu SY, Gordon T. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная аксотомия. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3876–3885. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Фу С.Ю., Гордон Т. Клеточные и молекулярные основы регенерации периферических нервов. Мол Нейробиол . 1997, февраль-апрель; 14 (1-2): 67–116. [PubMed] [Google Scholar] 7. Бойд Дж. Г., Гордон Т. Дозозависимое облегчение и ингибирование регенерации периферических нервов нейротрофическим фактором мозга. евро J Neurosci . 2002 Февраль; 15 (4): 613–626. [PubMed] [Google Scholar] 8. Сулейман О.А., Мидха Р., Манро К.А. и др. Хроническая денервация шванновских клеток и наличие сенсорного нерва снижают регенерацию моторных аксонов. Опыт Нейрол . 2002 август; 176 (2): 342–354. [PubMed] [Google Scholar] 9.Sulaiman OA, Voda J, Gold BG, Gordon T. FK506 увеличивает регенерацию периферических нервов после хронической аксотомии, но не после хронической денервации шванновских клеток. Опыт Нейрол . 2002 Май; 175 (1): 127–137. [PubMed] [Google Scholar] 10. Фу С.Ю., Гордон Т. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная денервация. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3886–3895. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Гордон Т., Тайреман Н., Раджи М.А. Основа для снижения функционального восстановления после отсроченного восстановления периферических нервов. Дж. Neurosci . 2011 6 апреля; 31 (14): 5325–5334. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Бойд Дж. Г., Гордон Т. Нейротрофические факторы и их рецепторы в регенерации аксонов и функциональном восстановлении после повреждения периферических нервов. Мол Нейробиол . 2003 июн; 27 (3): 277–324. [PubMed] [Google Scholar] 13. Сулейман О.А., Гордон Т. Трансформирующий фактор роста-бета и форсколин ослабляют неблагоприятные эффекты долгосрочной денервации шванновских клеток на регенерацию периферических нервов in vivo. Глия . 2002, 1 марта; 37 (3): 206–218. [PubMed] [Google Scholar] 14. Гордон Т., Фу С.Ю. Долгосрочная реакция на повреждение нерва. Adv Neurol . 1997. 72: 185–199. [PubMed] [Google Scholar] 15. Boyd JG, Gordon T. Нейротрофический фактор, полученный из линии глиальных клеток, и нейротрофический фактор, полученный из мозга, поддерживают регенерацию аксонов хронически аксотомированных мотонейронов in vivo. Опыт Нейрол . Октябрь 2003 г., 183 (2): 610–619. [PubMed] [Google Scholar] 16. Sulaiman OAR, Gordon T. TGF-бета обращает пагубный эффект длительной денервации шванновских клеток на регенерацию нервов, индуцируя экспрессию рецептора erbB3. Глия . 2003 сентябрь; 43 (Дополнение 2): 24. [Google Scholar] 17. Midha R, Munro CA, Chan S, Nitising A, Xu QG, Gordon T. Регенерация в защищенные и хронически денервированные культи периферических нервов. Нейрохирургия . 2005 декабрь; 57 (6): 1289–1299. [PubMed] [Google Scholar] 18. Сулейман О.А., Гордон Т. Роль хронической денервации шванновских клеток в плохом функциональном восстановлении после нервных повреждений и экспериментальные стратегии борьбы с ней. Нейрохирургия . Октябрь 2009 г., 65 (4 доп.): A105 – A114. [PubMed] [Google Scholar] 19.Cajal SR. Дегенерация и регенерация нервной системы . Оксфорд, Великобритания: Издательство Оксфордского университета; 1928. Перевод Р. М. Мэй. [Google Scholar] 20. Эртюрк А., Хеллал Ф., Энес Дж., Брадке Ф. Дезорганизованные микротрубочки лежат в основе образования ретракционных луковиц и нарушения регенерации аксонов. Дж. Neurosci . 22 августа 2007 г.; 27 (34): 9169–9180. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Моррис Дж. Х., Хадсон А. Р., Уэдделл Г. Исследование дегенерации и регенерации разделенного седалищного нерва крысы на основе электронной микроскопии.II. Развитие «регенерационного агрегата. Z Zellforsch Mikrosk Anat . 1972: 124 (1): 103–130. [PubMed] [Google Scholar] 22. McQuarrie IG. Влияние кондиционирующего поражения на формирование аксонального отростка в узлах Ранвье. Дж. Комп. Neurol . 1985, 8 января; 231 (2): 239–249. [PubMed] [Google Scholar] 23. Сулейман О.А., Бойд Дж. Г., Гордон Т. Регенерация аксонов в периферической системе млекопитающих. В: Kettenmann H, Ransom BR, editors. Нейроглия . 2-е изд. Оксфорд, Великобритания: Издательство Оксфордского университета; 2005 г.С. 454–466. В. ред. [Google Scholar] 24. Эйткен Дж. Т., Шарман М., Янг Дж. З. Созревание периферических нервных волокон с различными периферическими связями. Дж Анат . 1947; 81: 1–22. [PubMed] [Google Scholar] 25. Toft PB, Fugleholm K, Schmalbruch H. Разветвление аксонов после раздавливания периферических нервов крысы. Мышечный нерв . 1988 августа; 11 (8): 880–889. [PubMed] [Google Scholar] 26. Наварро X, Виво М., Валеро-Кабре А. Нейропластичность после повреждения и регенерации периферических нервов. Прог Нейробиол . Июль 2007 г .; 82 (4): 163–201. Epub 2007 22 июня. [PubMed] [Google Scholar] 27. Уоллер А. Эксперименты на разрезе языкоглоточного и подъязычного нервов лягушки и наблюдения за изменениями, вызванными этим в структуре их примитивных волокон. Philos Trans R Soc London . 1850; 140: 423–429. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Ван Дж.Т., Медресс З.А., Баррес Б.А. Дегенерация аксонов: молекулярные механизмы пути самоуничтожения. Дж. Ячейка Биол . 2012 9 января; 196 (1): 7–18. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Чжай К., Ван Дж., Ким А. и др. Участие убиквитин-протеасомной системы на ранних стадиях валлеровской дегенерации. Нейрон . 17 июля 2003 г.; 39 (2): 217–225. [PubMed] [Google Scholar] 31. Сулейман О.А., Клеточный Гордон Т. и молекулярные взаимодействия после повреждения периферических и центральных нервов. Биомед Ред. . 2003. 14: 51–62. [Google Scholar] 32. Хёке А., Ми Р. В поисках новых методов лечения периферических невропатий и регенерации нервов. Дисков Мед . 2007 август; 7 (39): 109–112. [PubMed] [Google Scholar] 33. Brushart TM. Ремонт нервов . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета ;; 2011. [Google Scholar] 34. Scherer SS, Salzer JL. Аксон-шванновские клеточные взаимодействия во время дегенерации и регенерации периферических нервов. В: Jessen KR, Richardson WD, редакторы. Развитие глиальных клеток: основные принципы и клиническая значимость . Оксфорд, Великобритания: Bios Scientific ;; 1996. С. 169–196. В. ред. [Google Scholar] 35. Хёке А., Гордон Т., Зочодне Д.В., Сулейман О.А.Снижение экспрессии нейротрофического фактора, происходящего из глиальных клеток, связано с нарушением регенерации после длительной денервации шванновских клеток. Опыт Нейрол . 2002 Янв; 173 (1): 77–85. [PubMed] [Google Scholar] 36. You S, Petrov T, Chung PH, Gordon T. Экспрессия рецептора фактора роста нервов с низким сродством в долгосрочно денервированных шванновских клетках. Глия . 1997 июн; 20 (2): 87–100. [PubMed] [Google Scholar] 37. Бисби М.А., Тецлафф В. Изменения в синтезе цитоскелетного белка после повреждения аксона и во время регенерации аксона. Мол Нейробиол . 1992; 6 (2-3): 107–123. Лето осень. [PubMed] [Google Scholar] 38. Гутманн Э. Влияние задержки иннервации на восстановление мышц после нервных поражений. Дж. Нейрофизиол . 1948 июл; 11 (4): 279–294. [PubMed] [Google Scholar] 40. Ли Х, Теренги Г, зал СМ. Влияние отсроченной реиннервации на экспрессию рецепторов c-erbB хронически денервированными шванновскими клетками крысы in vivo. Глия . 1997 августа; 20 (4): 333–347. [PubMed] [Google Scholar] 41. Тецлафф В., Леонард С., Крекоски К.А., Пархад И.М., Бисби Массачусетс.Снижение синтеза мотонейрональных нейрофиламентов путем последовательных аксотомий: возможное объяснение эффекта кондиционирующего поражения на регенерацию аксонов. Опыт Нейрол . 1996 Май; 139 (1): 95–106. [PubMed] [Google Scholar] 42. Гордон Т., Паттулло М.С. Пластичность мышечных волокон и типов двигательных единиц. Exerc Sport Sci Ред. . 1993; 21: 331–362. [PubMed] [Google Scholar] 44. Brushart TM, Mesulam MM. Трансганглионарная демонстрация центральных сенсорных проекций кожи и мышц с конъюгатами HRP-лектин. Neurosci Lett . 1980 апр; 17 (1-2): 1–6. [PubMed] [Google Scholar] 45. Гордон Т, Стейн РБ, Томас СК. Организация двигательных единиц после перекрестной реиннервации мышц-антагонистов в задней конечности кошки. Дж. Физиология . 1986 Май; 374: 443–456. [Бесплатная статья PMC] [PubMed] [Google Scholar] 46. Аль-Маджед А.А., Нойманн С.М., Брушарт TM, Гордон Т. Кратковременная электрическая стимуляция повышает скорость и точность регенерации моторных аксонов. Дж. Neurosci . 2000 г., 1 апреля; 20 (7): 2602–2608.[Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Мартини Р., Шахнер М., Брушарт TM. Углевод L2 / HNK-1 предпочтительно экспрессируется шванновскими клетками, ранее ассоциированными с моторными аксонами, в реиннервируемых периферических нервах. Дж. Neurosci . 1994, ноябрь; 14 (11, часть 2): 7180–7191. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Эберхардт К.А., Иринчев А.А., Аль-Маджед А.А. и др. Передача сигналов BDNF / TrkB регулирует экспрессию углеводов HNK-1 в регенерирующих двигательных нервах и способствует функциональному восстановлению после восстановления периферических нервов. Опыт Нейрол . 2006 Апрель; 198 (2): 500–510. Epub 2006 7 февраля. [PubMed] [Google Scholar] 49. Хёке А., Редетт Р., Хамид Х и др. Шванновские клетки выражают моторные и сенсорные фенотипы, которые регулируют регенерацию аксонов. Дж. Neurosci . 2006 20 сентября; 26 (38): 9646–9655. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Al-Majed AA, Brushart TM, Gordon T. Электрическая стимуляция ускоряет и увеличивает экспрессию мРНК BDNF и trkB в регенерирующих мотонейронах бедренной кости крысы. евро J Neurosci .2000 декабрь; 12 (12): 4381–4390. [PubMed] [Google Scholar] 51. Brushart TM, Hoffman PN, Royall RM и др. Электрическая стимуляция способствует регенерации мотонейрона без увеличения его скорости и без кондиционирования нейрона. Дж. Neurosci . 2002, 1 августа; 22 (15): 6631–6638. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Гордон Т., Сулейман О.А., Ладак А. Глава 24: Электростимуляция для улучшения регенерации нервов: где мы находимся? Международная версия Neurobiol . 2009. 87: 433–444. [PubMed] [Google Scholar] 53.Мидха Р. Новые методы восстановления нервов: нервные передачи и нервные трубки. Clin Neurosurg . 2006. 53: 185–190. [PubMed] [Google Scholar] 54. Пфистер Л.А., Папалоизос М., Меркл Г.П., Гандер Б. Нервные проводники и доставка факторов роста при восстановлении периферических нервов. J Peripher Nerv Syst . 2007 июн; 12 (2): 65–82. [PubMed] [Google Scholar]
От лабораторных исследований до прикроватных приложений
Охснер Дж. 2013 Весна; 13 (1): 100–108.
, MD, PhD, FRCS * и, PhD †
Wale Sulaiman
* Отделение нейрохирургии, Ochsner Clinic Foundation, и Медицинская школа Университета Квинсленда, Клиническая школа Охснера, Новый Орлеан, Луизиана
Тесса Гордон
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
* Отделение нейрохирургии, Фонд клиники Окснера и Школа медицины Университета Квинсленда, Клиническая школа Окснера, Новый Орлеан, Луизиана
† Отделение хирургии, Отделение пластической реконструктивной хирургии, Больница для больных детей, Университет Торонто, Торонто, Канада
Адресная корреспонденция Wale Sulaiman, MD, PhD, FRCS, Отделение нейрохирургии, Ochsner Clinic Foundation, 1514 Jefferson Hwy., New Orleans, LA 70121. Тел: (504) 842-2181 Факс: (504) 842-6531 Электронная почта: gro.renshco@namialusw Авторские права © Академическое подразделение Фонда клиники Окснера Эта статья цитируется в других статьях в PMC.
РЕФЕРАТ
Объективы
Мы рассматриваем современную нейробиологию повреждения и регенерации нервов, особенно в том, что касается восстановления полезной функции у пациентов, которые получили травмы крупных нервных стволов, таких как плечевое сплетение.
Методы
В этом обзоре основное внимание уделяется исследованиям, проведенным в нашей лаборатории в Окснеру и в других лабораториях, связанных с нейробиологией повреждения нервов, с акцентом на то, как некоторые из ключевых результатов исследований на животных помогают нам понять патофизиологию плохого функционального восстановления после нервных окончаний. травма, повреждение.
Выводы
Опубликованные исследования нейробиологии повреждения и регенерации нервов убедительно свидетельствуют о том, что хроническая денервация шванновских клеток, хроническая аксотомия нейронов и неправильное направление регенерирующих аксонов в неправильные эндоневральные трубки в первую очередь ответственны за плохое функциональное восстановление. Эффект атрофии мышечной денервации вторичен. Экспериментальные терапевтические стратегии (которые мы в настоящее время исследуем в нашей лаборатории в Охснере) для борьбы с этими 3 нейробиологическими явлениями могут улучшить восстановление функции у пациентов, перенесших нервные повреждения.
Ключевые слова: Аксотомия , цитокины , денервация , факторы роста нервов , регенерация нервов , нейробиология , Повреждения периферических нервов , восстановление функций
ВВЕДЕНИЕ
Повреждения нервов запускают сложные клеточно-молекулярные взаимодействия, которые необходимы для регенерации аксонов и, как следствие, для значимого функционального восстановления пациентов.Эти клеточные и молекулярные ответы ненейрональных клеток периферической нервной системы (ПНС), а именно шванновских клеток (СК), создают благоприятную для роста среду для поврежденных аксонов, в отличие от реакции центральной нервной системы (ЦНС) на повреждения. Однако существует дихотомия между способностью поврежденных нейронов ПНС регенерировать свои аксоны после повреждения и субоптимальным восстановлением функции после повреждения нервов.
Значительные успехи были достигнуты в технике микрохирургии поврежденных нервов, что иногда приводит к улучшению результатов для пациентов. 1-3 К сожалению, функциональное восстановление после травм периферических нервов часто остается слабым, особенно когда повреждены нервные стволы, расположенные близко к спинному мозгу и далеко от органов-мишеней. Эти травмы включают большинство травм отрывного типа, разрывов нервов и ушибов нервов. После этих типов нервных повреждений поврежденные нейроны должны регенерировать свои аксоны на больших расстояниях с очень медленной скоростью 1 мм / день. При такой медленной скорости регенерации восстановление функциональной двигательной единицы или реиннервация органа чувств может занять месяцы или даже годы ( 1-4 ), состояние, называемое хронической аксотомией. 5-7 Аналогичным образом, SC в дистальных обрубках нерва и в органах-мишенях остаются денервированными в течение длительных периодов, состояний, известных как хроническая SC денервация 2,8-11 и хроническая денервация мышц 11 соответственно.
Схематическое изображение повреждений проксимального нерва плечевого сплетения, которые требуют длительного времени регенерации регенерирующих аксонов для реиннервации мышц руки. В это время поврежденные нейроны подвергаются хронической аксотомии, а шванновские клетки дистальной культи поврежденных нервов хронически денервированы.Графики на вставках демонстрируют прогрессирующее снижение способности поврежденных мотонейронов, которые регенерируют аксоны (число MN) и реиннервируют мышцы (число двигательных единиц [MU]), в результате хронической денервации шванновских клеток и хронической аксотомии нейронов.
Нарушение функционального восстановления кисти, например, обычно объясняется необратимой атрофией денервированных мышц и органов чувств и замещением жировой ткани. 1 Однако наши эксперименты за последние ~ 15 лет предоставляют убедительные доказательства того, что прогрессирующая неспособность хронически аксотомированных нейронов регенерировать свои аксоны и прогрессирующая неспособность хронически денервированных SCs поддерживать регенерирующие аксоны играют преобладающую роль в клинических неудачах функциональное восстановление после микрохирургического восстановления. 2,5,7,8,10-18 Плохое функциональное восстановление также усугубляется задержкой микрохирургического восстановления поврежденных нервов, когда пациенты наблюдаются для спонтанного функционального восстановления, обычно в течение 3-6 месяцев после травм тупым нервом. Регенерация аксонов в неправильные эндоневральные трубки и конечные органы — так называемое неправильное направление регенерирующих аксонов — является еще одним фактором, который способствует плохому функциональному восстановлению.
РЕГЕНЕРАЦИОННЫЙ РЕАКЦИЯ ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И ЕГО РОЛЬ В ФУНКЦИОНАЛЬНОМ РЕЗУЛЬТАТЕ
Начальная фаза регенерации аксонов
После повреждения нерва проксимальная и дистальная культя поврежденного нерва претерпевают структурные и молекулярные изменения, готовясь к процессу аксональной регенерации.Проксимальная культя претерпевает обратную дегенерацию до первого узла Ранвье, а затем каждый поврежденный аксон вырабатывает несколько дочерних аксонов. 19,20 Многие дочерние аксоны обрезаются; те, что остались, начинают процесс удлинения через дистальные культи нерва и составляют регенерирующие единицы. 21 Эта начальная стадия регенерации аксонов поддерживается как доступностью локально продуцируемых цитоскелетных материалов, так и продуцируемыми нейронами и транспортируемыми антероградно цитоскелетными белками, такими как актин и тубулин (A). 6,20,22,23 Регенерация аксонов протекает со скоростью 1-3 мм / сут, что соответствует медленной скорости транспорта цитоскелетных материалов. Дальнейшее удлинение и регенерация через дистальную культю нерва зависят от среды, поддерживающей рост, обеспечиваемой SC дистальных культей нерва. Отсутствие эндоневриальных каналов, нагруженных шванновскими клетками (полосы Бюнгнера), приводит к неправильной регенерации и образованию невром. 3,24-26
После повреждения нерва гены, связанные с регенерацией (RAG), временно активируются в нейронах, в то время как гены, связанные с нормальной синаптической передачей, подавляются (A и B).Шванновские клетки денервированных обрубков нерва подвергаются пролиферации во время дегенерации Валлера и экспрессируют множество RAG, в то время как миелин-ассоциированные гены подавляются (A). Профили генов поддерживают рост аксонов, но экспрессия очень недолговечна, так что со временем (в то время как регенерирующие аксоны растут с медленной скоростью 1 мм / сут) экспрессия RAG снижается, и способность поврежденных нейронов регенерировать свои аксонов и шванновских клеток, поддерживающих регенерацию, уменьшается (C).Примеры прогрессивного снижения уровней мРНК нанесены на графики для тубулина в нейронах и GDNF в шванновских клетках.
Роль шванновских клеток в регенерации аксонов
Дистальные культи перерезанных нервов подвергаются дегенеративному процессу, названному в честь Августа Валлера, валлеровской дегенерации. 27 Теперь мы понимаем, что валлеровская дегенерация является важным подготовительным этапом процесса регенерации аксонов, посредством которого удаляются молекулы, которые могут препятствовать регенерации.В недавнем обзоре события разбиты на три морфологически различимые стадии: острая дегенерация аксонов, латентный период и стадия резкой гранулярной дегенерации. 28 В течение часа после травмы отмирание как в проксимальной, так и в дистальной культях опосредуется каналом-опосредованным притоком кальция и активацией кальпаинов, которые расщепляют нейрофиламенты и связанные с микротрубочками компоненты, такие как спектрин и тубулин. В проксимальной культи эта перемычка относится к первому узлу Ранвье. Следующим этапом примерно в 24 часа, в течение которого продолжаются измерения аксонных потенциалов, 29 следует более длительный период разборки белков цитоскелета кальцием-активированными кальпаинами и убиквитин-протеасомной системой. 30 СК играют главную роль в этом процессе посредством фагоцитоза аксонов и миелиновых остатков. Они также секретируют хемоаттрактивные факторы, такие как интерлейкин-1 и хемоаттрактантный белок-1 моноцитов, которые рекрутируют макрофаги в денервированные дистальные культи нервов, которые вносят значительный вклад в фагоцитоз аксонов и миелиновых остатков. 23,31,32 Обломки аксонов высвобождают митогены, которые способствуют митотическому делению SC и инициируют сеть цитокинов и факторов транскрипции, которые стимулируют распад миелина и инвазию макрофагов и нацелены на миелин для фагоцитоза SC и макрофагами. 6,33
Преобразование из режима действия-потенциала-проводимости в режим, поддерживающий регенерацию: роль генов, связанных с регенерацией
Сразу после повреждения нерва потеря аксонального контакта запускает пролиферацию СК и их фенотип от миелинизирующего к немиелинизирующему фенотипу, поддерживающему рост. 6,23,34 Экспрессия матричной РНК (мРНК) миелин-ассоциированных белков, таких как P0 и миелин-ассоциированный гликопротеин, подавляется.Нейротрофины (например, фактор роста нервов, нейротрофический фактор головного мозга и нейротрофический фактор глиального происхождения), их рецепторы (например, p75, GFRA-1, GFRA-2) и молекулы адгезии (например, молекула адгезии нервных клеток) активируются при подготовке к процессу регенерации аксонов (). 6,26,35,36 Эти гены с усиленной регуляцией в совокупности называются генами, связанными с регенерацией (RAG). Изменение экспрессии генов, дегенерация и клиренс миелина и аксонов являются ключевыми особенностями процесса валлеровской дегенерации. 6,23,34
Точно так же нейроны, чьи нервы были повреждены, подавляют мРНК белков, необходимых для нейротрансмиссии, и активируют их для восстановления своих периферических процессов. 6,12,37 Следовательно, актин, тубулин и GAP-43 активируются сразу после повреждения (A и B). 6,37 Однако активация RAG не поддерживается ни в поврежденных нейронах, ни в SC. К 6 месяцам у экспериментальных животных подавляющая регуляция большинства мРНК с активизацией снижается, что приводит к потере среды, поддерживающей рост, для регенерации аксонов (A и B). 6,37 Значение ограниченной по времени активации RAG продемонстрировано в прогрессирующем снижении способности поврежденных нейронов регенерировать свои аксоны и SCs поддерживать регенерирующие аксоны по мере увеличения продолжительности восстановления нерва ().
Регенеративная способность снижается со временем из-за продолжительной аксотомии и денервации шванновских клеток. На крысах мы экспериментально увеличили либо (А) продолжительность времени, в течение которого мотонейроны не могли регенерировать свои аксоны (хроническая аксотомия; путем отсрочки наложения швов от культи проксимального нерва к недавно денервированной культе дистального нерва), либо (В) денервации. клеток Шванна в дистальных обрубках нерва (хроническая денервация; путем отсрочки наложения швов свежеразрезанной проксимальной культи нерва на культю нерва, которая была хронически денервирована до восстановления нерва).Способность нейронов к регенерации после хронической аксотомии мотонейронов или хронической денервации шванновских клеток и мышц-мишеней определялась либо путем (а) расчета количества реиннервируемых мышечных единиц (нервов и мышечных волокон, которые снабжает один мотонейрон) с использованием силы измерения в ответ на стимуляцию одиночных аксонов и мышечного нерва или (б) подсчет мечения мотонейронов, которые успешно регенерировали свои аксоны, путем нанесения ретроградного красителя на регенерирующие аксоны в культе нерва дистальнее места восстановления нерва.Оценки регенеративного успеха, полученные с помощью методов подсчета мотонейронов и подсчета двигательных единиц, хорошо согласуются, демонстрируя прогрессирующее снижение регенеративной способности как функцию (C) хронической аксотомии мотонейронов и (D) хронической денервации шванновских клеток. .
ЭКСПЕРИМЕНТАЛЬНЫЕ ПАРАДИГМЫ И ОЦЕНКА РЕГЕНЕРАЦИИ АКСОНА ПОСЛЕ ПОВРЕЖДЕНИЯ НЕРВА И МИКРОХИРУРГИЧЕСКОГО РЕМОНТА
Экспериментальные исследования, проведенные во время Второй мировой войны Гутманном и Янгом, показали, что замедленное восстановление нерва не влияет на скорость роста и количество аксонов. 38,39 Эти данные, свидетельствующие о крайней атрофии денервированных мышц, привели к ошибочному выводу, что плохое функциональное восстановление после повреждения нерва было связано с необратимой денервационной атрофией мышцы и ее неспособностью воспринимать иннервацию, особенно после длительных периодов воздействия. время. 38,39 Этот вывод стал довольно популярным и, к сожалению, часто повторяется даже в последних публикациях. Недавние данные нашей лаборатории и других источников не подтвердили ошибочный вывод, главным образом потому, что (1) авторы не оценили напрямую количество поврежденных нейронов, которые регенерировали в дистальную культи нерва, и тех, которые реиннервируют денервированные мышцы, и (2) подсчитали нервные волокна в дистальной культе. Культя нерва переоценивает количество регенерированных нервных волокон, потому что многие дочерние аксоны возникают из единичных регенерирующих нервных волокон в проксимальной культе нерва и могут ошибочно считаться репрезентативными для отдельных нервных волокон в проксимальной культи.
В нескольких экспериментах мы изучали процесс регенерации аксонов после немедленного и отсроченного восстановления повреждения нерва. Кроме того, мы использовали количественные методы подсчета мотонейронов, которые регенерировали свои аксоны в дистальные обрубки нервов, и подсчета количества реиннервируемых моторных единиц в целевых мышцах, чтобы оценить способность мотонейронов регенерировать свои аксоны и реиннервировать мышцы. Используя метод перекрестного шва на крысах, который позволяет независимо изучить эффекты отсроченной реиннервации дистальной культи нерва (так называемая хроническая или длительная денервация) и отсроченной регенерации нейронов до их мишеней (так называемая хроническая или длительная аксотомия), мы оценили количество регенерированных мотонейронов с использованием техники обратной мечения, при которой ретроградно транспортируемые флуоресцентные красители наносятся на срезанные концы реиннервируемых дистальных культей нерва, которые маркируют мотонейроны, регенерировавшие свои аксоны.Таким образом, мы смогли подсчитать количество мотонейронов, которые регенерировали свои аксоны в вентральном роге спинного мозга после наших окончательных экспериментов. Мы оценили количество двигательных единиц, применяя сверхмаксимальную стимуляцию к отдельным вентральным корешкам и измеряя общую мышечную силу. Затем мы выделили волокна вентрального корня и стимулировали их индивидуально, чтобы определить силу двигательных единиц. Затем количество реиннервируемых двигательных единиц рассчитывали путем деления мышечной силы на силу двигательных единиц.иллюстрирует нашу хирургическую парадигму и методы оценки регенерации аксонов и реиннервации мышц после повреждения и восстановления нерва.
Хроническая денервация шванновских клеток
Предоставление SCs среды, поддерживающей рост, тесно связано с потерей и своевременным восстановлением контакта аксонов с клетками. 10 SC, реиннервация которых задерживается регенерирующими аксонами, считаются хронически денервированными, и пагубное влияние хронической денервации на SC ухудшается по мере увеличения продолжительности хронической денервации. 2,10,18 В наших экспериментах, чтобы проверить эффект хронической денервации, общая малоберцовая (CP) ветвь седалищного нерва перерезается, и ее реиннервация откладывается на период от 1 до 6 месяцев до наложения поперечного шва. проксимальная культя свежеперерезанного большеберцового (TIB) нерва в хронически денервированный дистальный CP-нерв. Таким образом, можно непосредственно оценить влияние хронической денервации ЦП на регенерацию свежеповрежденных мотонейронов TIB. Используя эту парадигму, хроническая денервация уменьшила количество мотонейронов, которые были обратно помечены фторорубовым красителем, который был нанесен на дистальную культи нерва в 10 мм от места перекрестного шва, до менее чем 10% от количества, которое регенерировалось после немедленного ушивания культи нервов. ().Было превосходное соответствие между этой долей мотонейронов, которые регенерировали свои аксоны в хронически денервированную культю нерва, и долей недавно аксотомированных мотонейронов, которые регенерировали и реиннервировали денервированную мышцу через 4-6 месяцев (). 10 Уменьшение количества мотонейронов, которые регенерируют свои аксоны через хронически денервированные SC, не только реиннервирует денервированные мышечные волокна, но и реиннервирует в 3 раза больше мышечных волокон, чем обычно.Поскольку это увеличение реиннервируемых двигательных единиц составляет максимальную способность к отрастанию мотонейронов, 2,10 , отсюда следует, что неспособность хронически денервированных мышечных волокон принять реиннервацию ограничивает функциональное восстановление после хронических нервных повреждений. 2,10,23 Устойчивая способность денервированной мышцы принимать реиннервацию также была продемонстрирована параллельным восстановлением как мышечной массы, так и изометрической силы. Тем не менее, изоляция эффектов денервации SC от денервации мышц показала, что хронически денервированные мышечные волокна не полностью восстановили свой прежний размер, утверждая, что ограниченное количество сателлитных клеток включается в каждое атрофическое мышечное волокно для восстановления площади поперечного сечения мышечных волокон. 11 Следовательно, прогрессирующее ухудшение способности SCs поддерживать рост в дистальных культях нерва играет главную роль в плохом функциональном восстановлении после повреждения нерва, а роль мышечной атрофии вторична. Поразительно, что долгоживущие хронически денервированные SC сохраняли свою способность ремиелинизировать меньшее количество регенерированных аксонов, 2 , особенно с учетом прогрессирующей регрессии способности денервированных SC поддерживать свой разрешающий рост фенотип и прогрессирующее снижение. в количестве поддерживающих рост SC в хронически денервированных культях дистального нерва. 36,40
Клиническим коррелятом нашей экспериментальной парадигмы перекрестного шва являются повреждения больших культи нерва, при которых СК дистальных культей нерва остаются хронически денервированными из-за медленной скорости регенерации аксонов. Чем больше времени требуется регенерирующим аксонам для реиннервации СК в дистальных культях нерва, тем дольше длится период хронической денервации, в течение которого СК не контактируют с аксонами, и тем выше вероятность того, что их способность поддерживать регенерирующие аксоны нарушена. 2,6,10 Мы считаем, что отчасти поэтому реиннервация и последующее функциональное восстановление в мышечных мишенях, расположенных рядом с регенерирующими мотонейронами, лучше, чем в более дистально расположенных мышечных мишенях.
Хроническая аксотомия нейронов
Поврежденные нейроны, аксоны которых могут или не могут регенерироваться, но не установили целевые соединения, подвергаются хронической аксотомии. 5,12,14 Например, после повреждения плечевого сплетения поврежденные нейроны должны регенерироваться на большом расстоянии, прежде чем они смогут реиннервировать некоторые денервированные мышцы и, таким образом, подвергнуться хронической аксотомии.Чтобы проверить эффект хронической аксотомии, мы также использовали технику перекрестного шва с использованием 2 ветвей седалищного нерва крысы. Ветвь большеберцовой кости седалищного нерва перерезается, и ее регенерация задерживается на периоды от 1 до 6 месяцев до наложения поперечного шва проксимальной культи хронически аксотомированной проксимальной культи большеберцового нерва в только что разрезанный дистальный центральный нерв. Таким образом, можно напрямую оценить влияние хронической аксотомии TIB на реиннервацию только что поврежденного дистального CP-нерва. Используя эту парадигму и показатели результатов, описанные выше, мы обнаружили, что количество мотонейронов, регенерировавших свои аксоны, постепенно снижалось в зависимости от продолжительного времени хронической аксотомии до ~ 37% от тех, которые регенерировали без эффекта хронической аксотомии. 5,7 Это снижение способности к регенерации после хронической аксотомии является значительным, особенно в сочетании с пагубным эффектом хронической денервации, которые возникают одновременно, особенно после травм крупных нервных стволов, таких как плечевое сплетение. Прогрессирующее снижение регенеративной способности хронически аксотомированных мотонейронов сопровождалось снижением активированных RAG. 41 Драматический негативный эффект снижения регенерационной способности хронически аксотомированных мотонейронов не повлиял на способность нейронов реиннервировать в 3 раза больше мышечных волокон, чем обычно, меньшее количество мотонейронов увеличивало свои двигательные единицы и количество мышечных волокон. снабжается каждым реиннервирующим мотонейроном. 5 Таким образом, хроническая аксотомия нарушает регенеративную способность мотонейронов, но не снижает способность меньшего числа регенерированных моторных аксонов устанавливать функциональные связи с денервированными мышечными волокнами.
Шахматная регенерация аксонов и неправильное направление регенерирующих аксонов
Одним из ключевых условий успешного функционального восстановления является то, что регенерирующие аксоны регенерируют в правильные эндоневральные трубки, которые направляют их обратно к исходным органам-мишеням.Однако мы обнаружили, что регенерирующие аксоны сталкиваются со значительной задержкой в месте повреждения и проходят через место повреждения в дистальные культи нерва в шахматном порядке (шахматная регенерация аксонов). Действительно, пересечение регенерирующих аксонов происходит медленно до входа регенерирующих аксонов в дистальные обрубки нерва. Только после того, как аксоны попадают в дистальную культю, они регенерируют с медленной скоростью транспорта 1-3 мм / сут.
Функциональные результаты после травм аксонов являются наилучшими после травм с раздавливанием нерва, при которых эндоневрий остается анатомически неповрежденным на всем пути к цели, а отростки аксонов регенерирующего блока содержатся в исходном эндоневрии, поэтому регенерирующие аксоны возвращаются в исходное положение. цели. 3 Со временем реиннервируемые мышечные волокна демонстрируют нормальное мозаичное распределение типов мышечных волокон, двигательные единицы и изометрические силы мышц полностью восстанавливаются, первоначальное количество функциональных двигательных единиц восстанавливается, а количество и диаметры миелинизированных нервных волокон возвращаются к прежним значениям. обычный. 42
Повреждения нервов, приводящие к повреждению эндоневральных трубок (например, аксонотметические или нейротметические повреждения), в результате которых регенерирующие аксоны должны найти свои исходные эндоневральные трубки, представляют собой потенциальную возможность ошибок в процессе регенерации.Отростки аксонов, исходящие из культи проксимального нерва, могут попадать в несколько различных эндоневральных трубок с целевыми точками, которые они раньше не обеспечивали, как в случае моторных аксонов, которые регенерируют в путях, ведущих к коже, а не к мышцам. 43 Эксперименты, в которых ретроградные красители применялись к регенерированным аксонам перерезанных седалищных и лицевых нервов у крыс, напрямую продемонстрировали неправильное направление регенерирующих моторных аксонов от нескольких различных пулов мотонейронов к мышцам, которые мотонейроны ранее не снабжали. 44 В случаях, когда нервы снабжали мышцы антагонистическими действиями, неправильное направление аксонов было связано с несоответствующими движениями, что вполне согласуется с данными о том, что мотонейроны сгибателей и разгибателей поддерживают свой нормальный, но несоответствующий паттерн возбуждения, когда они направлены на ненадлежащую реиннервацию мышц-разгибателей и сгибателей. , соответственно. 45 Мы считаем, что это неправильное направление регенерирующих поврежденных аксонов играет важную роль в снижении функционального восстановления после травм нервов.
Есть некоторые исключения из неправильного направления регенерирующих аксонов даже после травмы транзакции. К ним относятся несколько периферических нервных стволов, ветви которых иннервируют скелетные мышцы и органы чувств в коже и суставах. Примеры включают бедренный нерв с двумя ветвями, мышечную ветвь, содержащую двигательные и сенсорные нервы к четырехглавой мышце, и другую чисто сенсорную ветвь подкожного нерва к коже. 43 Хотя реиннервация моторных и сенсорных нервных ветвей изначально случайна, мотонейроны, которые постепенно регенерируют свои аксоны через место шва, посылают свои аксоны в соответствующую моторную ветвь. 43,46 Эта преимущественная реиннервация моторики возникла параллельно с прогрессирующей или ступенчатой регенерацией моторных аксонов в дистальных обрубках нерва. 46 Необычный кислый гликан, связанный с миелиновыми профилями моторных, но не сенсорных аксонов мыши, который распознается моноклональным антителом L2 / HNK-1, может быть одним из механизмов предпочтительной реиннервации соответствующих путей. 47,48 Дифференциальный профиль нейротрофических факторов в SCs, происходящих из сенсорных и моторных аксонов с профилем BDNF / NT4 / 5, также, вероятно, играет важную роль в предпочтительной реиннервации моторных путей. 49
ВЫВОДЫ
Огромный прогресс был достигнут в нашем понимании микроанатомии, патофизиологии и микрохирургического лечения поврежденных нервов. Эти достижения улучшили качество ухода за пациентами с повреждениями нервов и часто приводят к лучшему функциональному восстановлению. Однако пациенты, у которых не удается восстановить хорошую функцию, несмотря на превосходный микрохирургический уход, представляют собой проблему для нервного хирурга. Повышенная регуляция RAG недолговечна, и, по-видимому, существует временное окно возможностей, в течение которого SCs обеспечивают среду, поддерживающую рост, и поврежденные нейроны могут регенерировать свои аксоны.Ограниченная по времени повышающая регуляция RAG и медленная скорость регенерации аксонов приводят к прогрессирующей потере нейротрофической поддержки поврежденных нейронов и их регенерирующих аксонов, отсюда хроническая денервация SC и хроническая аксотомия нейронов. 5,6,23 Поэтому в нашей лаборатории в Ochsner мы фокусируемся на 2 возможных подходах к борьбе с разрывом между временем активации RAG и медленной скоростью регенерации, включая экспериментальные стратегии, которые ускоряют скорость регенерации аксонов и поддерживают нейротрофическая среда для регенерации аксонов в течение более длительных периодов времени.Мы продемонстрировали положительный эффект кратковременной (1 ч) низкочастотной электростимуляции на ускорение регенерации аксонов за счет модуляции экспрессии RAG. 46,50-52 Мы также показали положительный эффект трансформации фактора роста бета в реактивации и поддержании экспрессии RAGS хронически денервированными SC. В настоящее время мы изучаем роль трансплантации стволовых клеток в дистальные обрубки нерва и использование усиленных нервных направляющих каналов или проводников, которые не только устранят необходимость в аутотрансплантатах, но также позволят позитивно модулировать среду, способствующую росту аксонов. прохождение направляющих каналов в дистальные культи нерва. 53,54
Мы считаем, что окончательное решение для улучшения функционального восстановления в подгруппе пациентов, которые в настоящее время не справляются с микрохирургическим вмешательством, представляет собой комбинацию нескольких нейробиологических подходов для преодоления проблемы ограниченного временного окна для оптимального восстановления нерва. регенерация.
Сноски
Авторы не имеют финансовой или имущественной заинтересованности в предмете данной статьи .
Эта статья соответствует требованиям Совета по аккредитации для последипломного медицинского образования и Американского совета по медицинским специальностям, поддерживающим сертификационные компетенции по уходу за пациентами и медицинским знаниям.
СПИСОК ЛИТЕРАТУРЫ
1. Сулейман О.А.Р., Мидха Р., Гордон Т. Патофизиология хирургических нервных расстройств. В: Winn HR, редактор. Юманс Неврологическая хирургия . 6-е изд. Филадельфия, Пенсильвания: Сондерс ;; 2011. С. 2368–2379. В. изд. [Google Scholar] 2. Сулейман О.А., Гордон Т. Влияние краткосрочной и долгосрочной денервации шванновских клеток на регенерацию периферических нервов, миелинизацию и размер. Глия . 2000 декабрь; 32 (3): 234–246. [PubMed] [Google Scholar] 3. Сандерленд С. Нервы и травмы нервов .Эдинбург, Шотландия: Черчилль Ливингстон ;; 1978. [Google Scholar] 4. Сулейман В.А., Клайн Д.Г. Хирургия нервов: обзор и взгляд на будущее. Clin Neurosurg . 2006; 53: 38–47. [PubMed] [Google Scholar] 5. Fu SY, Gordon T. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная аксотомия. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3876–3885. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Фу С.Ю., Гордон Т. Клеточные и молекулярные основы регенерации периферических нервов. Мол Нейробиол . 1997, февраль-апрель; 14 (1-2): 67–116. [PubMed] [Google Scholar] 7. Бойд Дж. Г., Гордон Т. Дозозависимое облегчение и ингибирование регенерации периферических нервов нейротрофическим фактором мозга. евро J Neurosci . 2002 Февраль; 15 (4): 613–626. [PubMed] [Google Scholar] 8. Сулейман О.А., Мидха Р., Манро К.А. и др. Хроническая денервация шванновских клеток и наличие сенсорного нерва снижают регенерацию моторных аксонов. Опыт Нейрол . 2002 август; 176 (2): 342–354. [PubMed] [Google Scholar] 9.Sulaiman OA, Voda J, Gold BG, Gordon T. FK506 увеличивает регенерацию периферических нервов после хронической аксотомии, но не после хронической денервации шванновских клеток. Опыт Нейрол . 2002 Май; 175 (1): 127–137. [PubMed] [Google Scholar] 10. Фу С.Ю., Гордон Т. Факторы, способствующие плохому функциональному восстановлению после отсроченного восстановления нерва: длительная денервация. Дж. Neurosci . 1995 Май; 15 (5, часть 2): 3886–3895. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Гордон Т., Тайреман Н., Раджи М.А. Основа для снижения функционального восстановления после отсроченного восстановления периферических нервов. Дж. Neurosci . 2011 6 апреля; 31 (14): 5325–5334. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Бойд Дж. Г., Гордон Т. Нейротрофические факторы и их рецепторы в регенерации аксонов и функциональном восстановлении после повреждения периферических нервов. Мол Нейробиол . 2003 июн; 27 (3): 277–324. [PubMed] [Google Scholar] 13. Сулейман О.А., Гордон Т. Трансформирующий фактор роста-бета и форсколин ослабляют неблагоприятные эффекты долгосрочной денервации шванновских клеток на регенерацию периферических нервов in vivo. Глия . 2002, 1 марта; 37 (3): 206–218. [PubMed] [Google Scholar] 14. Гордон Т., Фу С.Ю. Долгосрочная реакция на повреждение нерва. Adv Neurol . 1997. 72: 185–199. [PubMed] [Google Scholar] 15. Boyd JG, Gordon T. Нейротрофический фактор, полученный из линии глиальных клеток, и нейротрофический фактор, полученный из мозга, поддерживают регенерацию аксонов хронически аксотомированных мотонейронов in vivo. Опыт Нейрол . Октябрь 2003 г., 183 (2): 610–619. [PubMed] [Google Scholar] 16. Sulaiman OAR, Gordon T. TGF-бета обращает пагубный эффект длительной денервации шванновских клеток на регенерацию нервов, индуцируя экспрессию рецептора erbB3. Глия . 2003 сентябрь; 43 (Дополнение 2): 24. [Google Scholar] 17. Midha R, Munro CA, Chan S, Nitising A, Xu QG, Gordon T. Регенерация в защищенные и хронически денервированные культи периферических нервов. Нейрохирургия . 2005 декабрь; 57 (6): 1289–1299. [PubMed] [Google Scholar] 18. Сулейман О.А., Гордон Т. Роль хронической денервации шванновских клеток в плохом функциональном восстановлении после нервных повреждений и экспериментальные стратегии борьбы с ней. Нейрохирургия . Октябрь 2009 г., 65 (4 доп.): A105 – A114. [PubMed] [Google Scholar] 19.Cajal SR. Дегенерация и регенерация нервной системы . Оксфорд, Великобритания: Издательство Оксфордского университета; 1928. Перевод Р. М. Мэй. [Google Scholar] 20. Эртюрк А., Хеллал Ф., Энес Дж., Брадке Ф. Дезорганизованные микротрубочки лежат в основе образования ретракционных луковиц и нарушения регенерации аксонов. Дж. Neurosci . 22 августа 2007 г.; 27 (34): 9169–9180. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Моррис Дж. Х., Хадсон А. Р., Уэдделл Г. Исследование дегенерации и регенерации разделенного седалищного нерва крысы на основе электронной микроскопии.II. Развитие «регенерационного агрегата. Z Zellforsch Mikrosk Anat . 1972: 124 (1): 103–130. [PubMed] [Google Scholar] 22. McQuarrie IG. Влияние кондиционирующего поражения на формирование аксонального отростка в узлах Ранвье. Дж. Комп. Neurol . 1985, 8 января; 231 (2): 239–249. [PubMed] [Google Scholar] 23. Сулейман О.А., Бойд Дж. Г., Гордон Т. Регенерация аксонов в периферической системе млекопитающих. В: Kettenmann H, Ransom BR, editors. Нейроглия . 2-е изд. Оксфорд, Великобритания: Издательство Оксфордского университета; 2005 г.С. 454–466. В. ред. [Google Scholar] 24. Эйткен Дж. Т., Шарман М., Янг Дж. З. Созревание периферических нервных волокон с различными периферическими связями. Дж Анат . 1947; 81: 1–22. [PubMed] [Google Scholar] 25. Toft PB, Fugleholm K, Schmalbruch H. Разветвление аксонов после раздавливания периферических нервов крысы. Мышечный нерв . 1988 августа; 11 (8): 880–889. [PubMed] [Google Scholar] 26. Наварро X, Виво М., Валеро-Кабре А. Нейропластичность после повреждения и регенерации периферических нервов. Прог Нейробиол . Июль 2007 г .; 82 (4): 163–201. Epub 2007 22 июня. [PubMed] [Google Scholar] 27. Уоллер А. Эксперименты на разрезе языкоглоточного и подъязычного нервов лягушки и наблюдения за изменениями, вызванными этим в структуре их примитивных волокон. Philos Trans R Soc London . 1850; 140: 423–429. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Ван Дж.Т., Медресс З.А., Баррес Б.А. Дегенерация аксонов: молекулярные механизмы пути самоуничтожения. Дж. Ячейка Биол . 2012 9 января; 196 (1): 7–18. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Чжай К., Ван Дж., Ким А. и др. Участие убиквитин-протеасомной системы на ранних стадиях валлеровской дегенерации. Нейрон . 17 июля 2003 г.; 39 (2): 217–225. [PubMed] [Google Scholar] 31. Сулейман О.А., Клеточный Гордон Т. и молекулярные взаимодействия после повреждения периферических и центральных нервов. Биомед Ред. . 2003. 14: 51–62. [Google Scholar] 32. Хёке А., Ми Р. В поисках новых методов лечения периферических невропатий и регенерации нервов. Дисков Мед . 2007 август; 7 (39): 109–112. [PubMed] [Google Scholar] 33. Brushart TM. Ремонт нервов . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета ;; 2011. [Google Scholar] 34. Scherer SS, Salzer JL. Аксон-шванновские клеточные взаимодействия во время дегенерации и регенерации периферических нервов. В: Jessen KR, Richardson WD, редакторы. Развитие глиальных клеток: основные принципы и клиническая значимость . Оксфорд, Великобритания: Bios Scientific ;; 1996. С. 169–196. В. ред. [Google Scholar] 35. Хёке А., Гордон Т., Зочодне Д.В., Сулейман О.А.Снижение экспрессии нейротрофического фактора, происходящего из глиальных клеток, связано с нарушением регенерации после длительной денервации шванновских клеток. Опыт Нейрол . 2002 Янв; 173 (1): 77–85. [PubMed] [Google Scholar] 36. You S, Petrov T, Chung PH, Gordon T. Экспрессия рецептора фактора роста нервов с низким сродством в долгосрочно денервированных шванновских клетках. Глия . 1997 июн; 20 (2): 87–100. [PubMed] [Google Scholar] 37. Бисби М.А., Тецлафф В. Изменения в синтезе цитоскелетного белка после повреждения аксона и во время регенерации аксона. Мол Нейробиол . 1992; 6 (2-3): 107–123. Лето осень. [PubMed] [Google Scholar] 38. Гутманн Э. Влияние задержки иннервации на восстановление мышц после нервных поражений. Дж. Нейрофизиол . 1948 июл; 11 (4): 279–294. [PubMed] [Google Scholar] 40. Ли Х, Теренги Г, зал СМ. Влияние отсроченной реиннервации на экспрессию рецепторов c-erbB хронически денервированными шванновскими клетками крысы in vivo. Глия . 1997 августа; 20 (4): 333–347. [PubMed] [Google Scholar] 41. Тецлафф В., Леонард С., Крекоски К.А., Пархад И.М., Бисби Массачусетс.Снижение синтеза мотонейрональных нейрофиламентов путем последовательных аксотомий: возможное объяснение эффекта кондиционирующего поражения на регенерацию аксонов. Опыт Нейрол . 1996 Май; 139 (1): 95–106. [PubMed] [Google Scholar] 42. Гордон Т., Паттулло М.С. Пластичность мышечных волокон и типов двигательных единиц. Exerc Sport Sci Ред. . 1993; 21: 331–362. [PubMed] [Google Scholar] 44. Brushart TM, Mesulam MM. Трансганглионарная демонстрация центральных сенсорных проекций кожи и мышц с конъюгатами HRP-лектин. Neurosci Lett . 1980 апр; 17 (1-2): 1–6. [PubMed] [Google Scholar] 45. Гордон Т, Стейн РБ, Томас СК. Организация двигательных единиц после перекрестной реиннервации мышц-антагонистов в задней конечности кошки. Дж. Физиология . 1986 Май; 374: 443–456. [Бесплатная статья PMC] [PubMed] [Google Scholar] 46. Аль-Маджед А.А., Нойманн С.М., Брушарт TM, Гордон Т. Кратковременная электрическая стимуляция повышает скорость и точность регенерации моторных аксонов. Дж. Neurosci . 2000 г., 1 апреля; 20 (7): 2602–2608.[Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Мартини Р., Шахнер М., Брушарт TM. Углевод L2 / HNK-1 предпочтительно экспрессируется шванновскими клетками, ранее ассоциированными с моторными аксонами, в реиннервируемых периферических нервах. Дж. Neurosci . 1994, ноябрь; 14 (11, часть 2): 7180–7191. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Эберхардт К.А., Иринчев А.А., Аль-Маджед А.А. и др. Передача сигналов BDNF / TrkB регулирует экспрессию углеводов HNK-1 в регенерирующих двигательных нервах и способствует функциональному восстановлению после восстановления периферических нервов. Опыт Нейрол . 2006 Апрель; 198 (2): 500–510. Epub 2006 7 февраля. [PubMed] [Google Scholar] 49. Хёке А., Редетт Р., Хамид Х и др. Шванновские клетки выражают моторные и сенсорные фенотипы, которые регулируют регенерацию аксонов. Дж. Neurosci . 2006 20 сентября; 26 (38): 9646–9655. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Al-Majed AA, Brushart TM, Gordon T. Электрическая стимуляция ускоряет и увеличивает экспрессию мРНК BDNF и trkB в регенерирующих мотонейронах бедренной кости крысы. евро J Neurosci .2000 декабрь; 12 (12): 4381–4390. [PubMed] [Google Scholar] 51. Brushart TM, Hoffman PN, Royall RM и др. Электрическая стимуляция способствует регенерации мотонейрона без увеличения его скорости и без кондиционирования нейрона. Дж. Neurosci . 2002, 1 августа; 22 (15): 6631–6638. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Гордон Т., Сулейман О.А., Ладак А. Глава 24: Электростимуляция для улучшения регенерации нервов: где мы находимся? Международная версия Neurobiol . 2009. 87: 433–444. [PubMed] [Google Scholar] 53.Мидха Р. Новые методы восстановления нервов: нервные передачи и нервные трубки. Clin Neurosurg . 2006. 53: 185–190. [PubMed] [Google Scholar] 54. Пфистер Л.А., Папалоизос М., Меркл Г.П., Гандер Б. Нервные проводники и доставка факторов роста при восстановлении периферических нервов. J Peripher Nerv Syst . 2007 июн; 12 (2): 65–82. [PubMed] [Google Scholar]
Повреждение периферического нерва — обзор
Визуализирующие исследования
Дополнительные инструменты для неврологической оценки заболеваний периферических нервов включают электродиагностические исследования; ЭМГ и NCS, магнитно-резонансная томография (МРТ), компьютерная томография (КТ), миелография, миелограмма КТ, магнитно-резонансная нейрография (МРН) и простая рентгенография.
Обычный рентгеновский снимок может выявить некоторые опухоли, кисты, повреждения диафрагмы (диафрагмального нерва) и переломы скелета, определенные места которых могут указывать на сопутствующее повреждение нерва. Однако периферические нервы плохо отделяются от окружающей ткани. 35
КТ может идентифицировать опухоли мягких тканей, кисты и переломы скелета или кровоизлияния и может помочь выделить некоторые опухоли периферических нервов из соседних кровеносных сосудов или тканей, хотя в большинстве случаев визуализация обычно плохая. 36
МРТ может определить анатомические детали мягких тканей, периферических нервов и периневральной ткани намного лучше, чем КТ. Он может дать многоплоскостные изображения T1 и T2 (аксиальные, корональные, сагиттальные) больших объемных опухолей. 37 Корональная и сагиттальная проекции показывают полную линейную протяженность опухоли с меньшим количеством разрезов и дают более четкие изображения, чем КТ. CT может потребовать нескольких осевых разрезов. МРТ может использоваться у пациентов с неферромагнитным протезированием. Это также полезно для оценки и диагностики травматических, компрессионных и воспалительных поражений периферических нервов. 35,37 Неадекватные изображения обычно вторичны из-за пространственных ограничений и ограничений разрешения и артефактов движения. 20,38 Травматическое повреждение периферического нерва может привести к потере аксонов с дегенерацией и денервацией мышц с последующей атрофией мышц, если травма носит хронический характер. МРТ мышцы, расположенной дистальнее поражения, может показать повышенные сигналы T2 и короткое восстановление инверсии тау-белка (STIR) в денервированной мышце. Это хорошо коррелирует с клиническим обследованием (слабость 3 степени) и данными ЭМГ о возможностях фибрилляции.Эти сигналы можно увидеть уже через 4 дня после повреждения нерва. 39 МРТ в некоторых случаях позволяет отличить нейропрактику от аксонометрических и нейрометрических повреждений. 39,40 Как STIR, так и T2-взвешенные изображения являются нормальными при нейрапраксических поражениях.
При аксонометрических травмах признаки реиннервации и восстановления отражаются нормализацией сигналов, но результаты МРТ могут отставать. 39 Следовательно, хирургическое обследование с интраоперационными электрическими исследованиями необходимо для надлежащей оценки аксонометрических и нейрометрических повреждений.Это особенно важно, потому что нейрометрические травмы требуют хирургического вмешательства для любого шанса на полезное функциональное восстановление. Повышенные сигналы T2 и STIR также наблюдаются в хронически денервированных мышцах и нервах, например, при ущемлении нейропатий, таких как синдром запястного канала срединного нерва или защемление локтевого нерва в локтевом суставе. При синдроме запястного канала эти данные хорошо коррелируют с ЭМГ, исследованиями нервной проводимости, клиническим обследованием и оперативными данными; однако в локтевом нерве, где локализация симптомов с помощью электродиагностических исследований затруднена, корреляция с визуализацией менее очевидна. 40-43 По неизвестным причинам некоторые аномальные сигналы МРТ нормализуются в течение месяцев даже без признаков выздоровления. 42
В крупном исследовании позвоночника было показано, что МРТ с контрастом (гадолиний) и без него позволяет количественно оценить перидуральный фиброз поясничного отдела через 6 месяцев после дискэктомии по поводу грыжи поясничного диска. 16 Увеличенный рубец хорошо коррелировал с повышенной вероятностью рецидива корешковой боли. Визуализация в этом случае больше смотрела на рубцовую ткань, чем на сам нерв.
Несмотря на все преимущества МРТ и КТ, в некоторых случаях все еще сложно отличить сосудистые структуры и окружающие ткани от периферических нервов. Несмотря на превосходное усиление контраста опухолей с помощью МРТ, его полезность варьируется при заболеваниях периферических нервов. 44 Развитие MRN помогло решить эту проблему благодаря его способности напрямую визуализировать нормальную фасикулярную структуру и рисунок нерва посредством межпучкового и внутрипучкового подавления неневральных структур, таких как жир, мышцы и кровеносные сосуды. 26,35,44 Следовательно, сигнал нервного пучка преобладает. Изображения получены с использованием катушек с фазированной решеткой, последовательностей быстрого спинового эха (FSE) и T2-взвешенных изображений с подавлением жира. Сигналы усиливаются (гиперинтенсивны) при внутриневральном заболевании и коррелируют с клиническим обследованием, интраоперационными электродиагностическими исследованиями и патологией. 38 Импульсы спин-эхо T1 могут показать направление нерва по отношению к соседним структурам. 38,44 Продольные и поперечные фасцикулярные изображения помогают отличить интраневральные образования от периневральных, таких как опухоли или кисты. 44 В одном исследовании предоперационная MRN отличила нефункционирующие и функционирующие пучки от опухоли, что позволило провести полную резекцию у 72,5% пациентов с NF1. 38 После травматического повреждения периферического нерва MRN может оценить целостность нерва и помочь в планировании операции. 44,45 Cudlip et al. 17 использовали модель раздавливания седалищного нерва крысы, чтобы проследить изменения сигнала MRN сразу после травмы и на протяжении дегенеративных и регенеративных процессов.Изменения сигнала коррелировали с функциональным дефицитом. Нормализация сигналов коррелировала с функциональным восстановлением. В начале после тяжелого аксонометрического или нейрометрического повреждения сигналы T2 усиливаются в области поражения и дистальнее. Электрические исследования не показывают нервной проводимости через поражение.
После хирургического восстановления периферического нерва за ходом заживления могут следовать изменения сигнала. 26,44 Через несколько недель или месяцев признаки регенерации могут быть подтверждены клиническим обследованием, уменьшением фибрилляции на ЭМГ и дистальной нормализацией сигналов Т2.Однако по неизвестным причинам эти сигналы могут оставаться высокими в межпозиционных трансплантатах. 26 В целом, эти данные показывают, что MRN хорошо коррелирует с клиническими и интраоперационными данными электродиагностики в процессе выздоровления. 26,38
МРТ, КТ и миелография могут быть очень чувствительными, но не специфичными при обнаружении многоуровневого остеохондроза шейного и поясничного отделов позвоночника. 43 У бессимптомных пациентов бывает много ложноположительных результатов.В этих случаях можно рассмотреть возможность использования MRN. Это может быть полезным дополнением, поскольку уже было показано, что он помогает оценивать и диагностировать травматические и компрессионные поражения периферических нервов, нервных корешков и плечевого сплетения. 35,45 Внешний вид MRN может отличаться в зависимости от симптоматического корня, что позволяет идентифицировать патологический уровень позвоночника.
Оценка острого плечевого паралича, вторичного по отношению к отрыву корня шейки матки, может быть сложной и трудной. Это требует тщательной клинической оценки, визуализации и электродиагностических исследований.Травматические менингоцеле (псевдоменингоцеле) в исследованиях миелограмм считались патогномоничными для интрадурального (интраспинального) отрыва корешков спинного мозга (рис. 195-2). 20,22 Однако хирургическое обследование в некоторых случаях выявляет интрадуральный отрыв корня даже при нормальной предоперационной миелограмме. В настоящее время КТ миелограмма, выполненная на аксиальных срезах размером 1-3 мм, считается точной для оценки отрыва корешков спинного мозга, особенно в плечевом сплетении. 20 Значительное исследование показало, что 85% корешков, оцененных с помощью предоперационной КТ-миелограммы, коррелируют с интраоперационными данными об интрадурально оторванных корешках шейного отдела позвоночника по сравнению с 52%, оцененными с помощью МРТ. 20,42 Вырванные корешки спинного мозга чаще ошибочно диагностируются на С5 или С6 в обоих исследованиях визуализации. МРТ менее надежна при визуализации интрадуральных (интраспинальных) дорсальных и вентральных корешков. 20 Интрадуральный фиброз или травматические менингоцеле (псевдоменингоцеле) могут быть факторами плохой визуализации. При неубедительных или сложных повреждениях плечевого сплетения MRN следует рассматривать в сочетании с традиционными МРТ и электрофизиологическими исследованиями, чтобы помочь локализовать поражения нервных корешков и плечевого сплетения. 45
Травма периферического нерва: текущие отчеты спортивной медицины
Анатомия
Моторный блок: клеточного тела, расположенного в переднем роге спинного мозга, моторном аксоне и мышечном блоке, который он иннервирует.
Сенсорный нейрон: тело клетки, расположенное в ганглии дорсального корешка и сенсорных аксонах, простирающихся на периферию и в центральную нервную систему.
Эндоневрий: соединительной ткани, окружающей отдельный миелинизированный аксон.
Perineurium: соединительная ткань, связывающая множество аксонов в пучки.
Эпиневрий: соединительнотканных пучков, связывающих периферический нерв.
Патофизиология
- Периферические нервы могут быть повреждены прямой травмой, растяжением или тракцией, вибрацией, сжатием, разрывом или ишемией / повреждением микрососудов. Ятрогенное повреждение нерва в результате спортивной хирургии, к счастью, встречается редко ( 2 ).
- Классификация нервных повреждений по классификации Седдона ( 3 ):
- ○ Нейрапраксия: наименее тяжелая из-за повреждения только миелина и сохранения целостности окружающей соединительной ткани и аксонов. Ожидается полное выздоровление.
- ○ Аксонотмезис: повреждение самого аксона, ведущее к валлеровской дегенерации (дистальная часть отделенного аксона дегенерирует вдали от повреждения), но целостность окружающей соединительной ткани сохраняется.Полное выздоровление возможно, но менее вероятно.
- ○ Нейротмезис: наиболее серьезное повреждение, приводящее к нарушению работы аксонов (с дегенерацией Валлера) и окружающих слоев соединительной ткани. Полное выздоровление идет плохо даже при хирургическом вмешательстве.
- Прогноз зависит от тяжести и количества поврежденных аксонов, а также от целостности окружающей соединительной ткани.
Признаки / симптомы
- Повреждение нерва может вызвать боль, слабость или парестезию, как правило, в области пораженного нерва.Повреждение двигательного нерва также может привести к атрофии мышц.
- Повреждение нерва, особенно в результате чрезмерного использования или прогрессирующего сжатия, может имитировать другие распространенные нарушения опорно-двигательного аппарата и должно быть заподозрено, если не обнаружено других повреждений костей, мягких тканей или сосудов.
Диагностические исследования
- Магнитно-резонансная томография: используется для оценки повреждения структуры окружающих мягких тканей или наличия ганглиозных кист, которые могут приводить к компрессии нерва.
- Ультразвук: позволяет оценить отек аксонов, наличие образования невромы или сдавления нерва, а также целостность нерва.
- Электромиография (ЭМГ) и исследования нервной проводимости (NCS) являются золотым стандартом для определения местоположения, тяжести и прогноза повреждения нерва.
- При тестировании ЭМГ оценивается потенциал действия двигательной единицы. Наличие фибрилляции и положительных острых волн указывает на острое неврологическое повреждение или заболевание, и они обычно проявляются через 10–14 дней после острого повреждения.
- NCS измеряет скорость нервной проводимости, чтобы определить дегенерацию аксонов в результате демиелинизирующих заболеваний. Зубцы F присутствуют при периферических невропатиях с более проксимальным поражением, но сами по себе не могут диагностировать радикулопатию. Отсутствие H-рефлекса полезно для диагностики радикулопатии S1 ( 1 ).
- Диагностическая блокада нервов под контролем УЗИ или рентгеноскопии может быть полезна для определения конкретного поражения нерва.
Лечение травм нерва
- Безоперационное лечение повреждений нервов включает защиту от дальнейшего повреждения (травмы или продолжающееся сжатие) и сохранение диапазона движений пораженной конечности / сустава.
- Первоначальный контроль боли обычно достигается с помощью нестероидных противовоспалительных препаратов. С невропатической болью лучше всего справиться с помощью трициклических антидепрессантов, габапентина, прегабалина или селективных ингибиторов обратного захвата серотонина (СИОЗС) / ингибитора обратного захвата серотонина и норэпинефрина (ИОЗСН).
- Лечебная физкультура, предназначенная для укрепления опорных или вспомогательных групп мышц, может помочь уменьшить потерю общей функции из-за более серьезных травм нервов.
- Реиннервация возможна, но медленно, потому что аксоны растут со скоростью около 1 мм · d −1 .
- Острое хирургическое вмешательство рекомендуется при рваных ранах, где возможен анастомоз нервов, или при синдроме острого компартмента, когда вскрытие пораженного компартмента может предотвратить ишемию закрытых нервов.
Список литературы
1. Чо С.К., Ферранте М.А., Левин К.Х. и др. Полезность электродиагностического тестирования при оценке пациентов с пояснично-крестцовой радикулопатией: обзор, основанный на фактах. Мышечный нерв . 2010; 42: 276–82.
2. Маак Т.Г., Осей Д., Делос Д. и др. Повреждения периферических нервов в спортивной хирургии: презентация, оценка и лечение. J. Bone Joint Surg. Являюсь. 2012; 94: e121 (1–10).
3. Нил С., Филдс КБ.Ущемление периферического нерва и повреждение верхней конечности. Am. Fam.